
The Megadiverse Australian Ant Genus Melophorus: Using CO1 Barcoding to Assess Species Richness

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
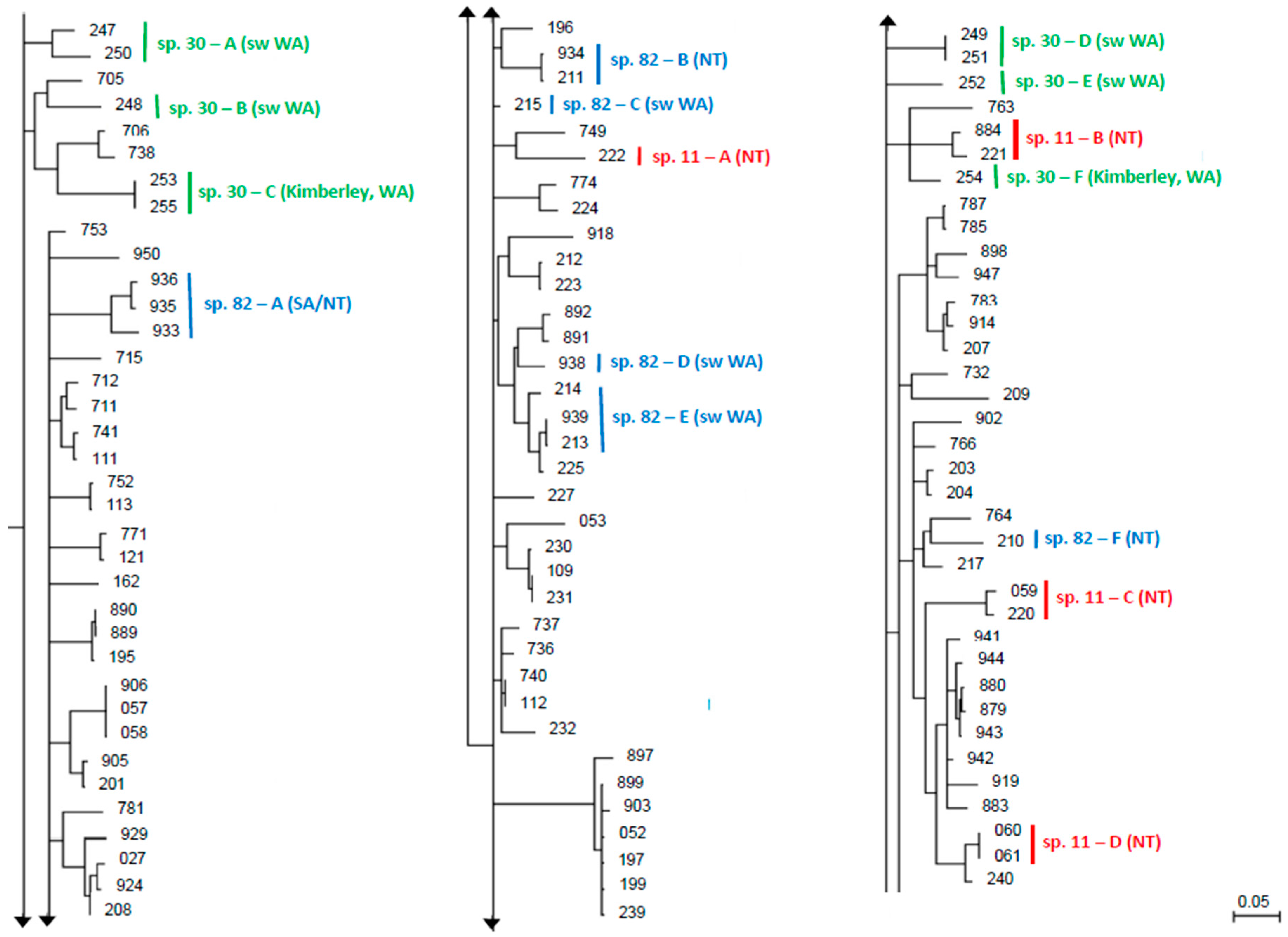
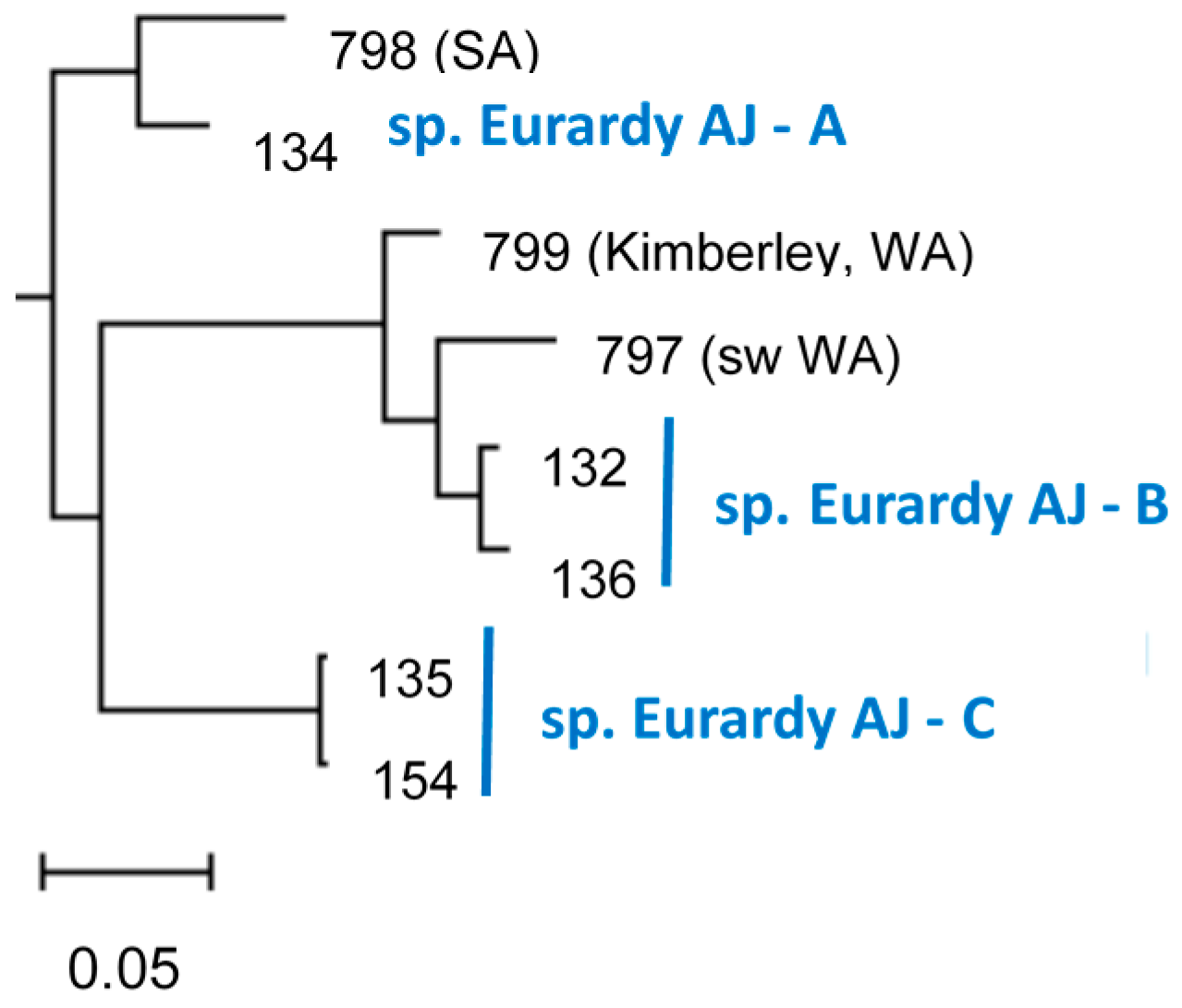
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Greenslade, P.J.M. A Guide to the Ants of South Australia; South Australian Museum: Adelaide, Australia, 1979. [Google Scholar]
- Andersen, A.N. Ant biodiversity in arid Australia: Productivity, species richness and community organization. Rec. S. Aust. Mus. Monogr. Ser. 2003, 7, 79–92. [Google Scholar]
- Conway, J.R. Notes on the excavation of a nest of Melophorus bagoti Lubbock in the Northern Territory, Australia (Hymenoptera, Formicidae). J. Aust. Entomol. Soc. 1992, 31, 247–248. [Google Scholar] [CrossRef]
- Christian, K.A.; Morton, S.R. Extreme thermophilia in a central Australian ant, Melophorus bagoti. Physiol. Zool. 1992, 65, 885–905. [Google Scholar] [CrossRef]
- Schultheiss, P.; Nooten, S.S. Foraging patterns and strategies in an Australian desert ant. Austral Ecol. 2013, 38, 942–951. [Google Scholar] [CrossRef]
- Musa, B.; Sommer, S.; Wolf, H.; Wehner, R. Foraging ecology of the thermophilic Australian desert ant, Melophorus bagoti. Aust. J. Zool. 2005, 53, 301–311. [Google Scholar]
- Narendra, A. Homing strategies of the Australian desert ant Melophorus bagoti. I. Proportional path-integration takes the ant half-way home. J. Exp. Biol. 2007, 210, 1798–1803. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Narendra, A.; Sommer, S.; Wehner, R. Traveling in clutter: Navigation in the central Australian desert ant Melophorus bagoti. Behav. Processes 2009, 80, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Schultheiss, P.; Schwarz, S.; Wystrach, A. Nest Relocation and Colony Founding in the Australian Desert Ant, Melophorus bagoti Lubbock (Hymenoptera: Formicidae). Psyche 2010, 2010. [Google Scholar] [CrossRef]
- Hoffmann, B.D. Thermophilia in a tropical Australian ant of the Melophorus aeneovirens (Lowne) species-group (Hymenoptera: Formicidae). Aust. J. Entomol. 1998, 37, 162–167. [Google Scholar] [CrossRef]
- Majer, J.D.; Gove, A.D.; Sochacki, S.; Searle, P.; Portlock, C. A comparison of the autecology of two seed-taking ant genera, Rhytidoponera and Melophorus. Insectes Soc. 2011, 58, 115–125. [Google Scholar] [CrossRef]
- Schultheiss, P.; Schwarz, S.; Cheng, K.; Wehner, R. Foraging ecology of an Australian salt-pan desert ant (genus Melophorus). Aust. J. Zool. 2012, 60, 311–319. [Google Scholar] [CrossRef]
- Snelling, R.R. A revision of the honey ants, genus Myrmecocystus (Hymenoptera: Formicidae). Nat. Hist. Mus. Los Angel. Cty. Sci.Bull. 1976, 24, 1–163. [Google Scholar]
- Agosti, D. Review and reclassification of Cataglyphis (Hymenoptera, Formicidae). J. Nat. Hist. 1990, 24, 1457–1505. [Google Scholar] [CrossRef]
- Clark, J. Australian Formicidae. Notes and new species. Mem. Natl. Mus. Vic. 1941, 12, 71–94. [Google Scholar]
- Agosti, D. Two new engimatic Melophorus species (Hymenoptera: Formicidae) from Australia. J. N. Y. Entomol. Soc. 1997, 105, 161–169. [Google Scholar]
- Andersen, A.N. Seed-harvesting by ants in Australia. In Ant-Plant Interactions; Huxley, C.R., Cutler, D.F., Eds.; Oxford University Press: Oxford, UK, 1991; pp. 493–503. [Google Scholar]
- Andersen, A.N. Ant diversity in arid Australia: A systematic overview. In Advances in Ant (Hymenoptera: Formicidae) Systematics: Homage to E. O. Wilson—50 Years of Contributions; Snellling, R.R., Fisher, B.L., Ward, P.S., Eds.; Memoirs American Entomological Institute Series; American Entomological Institute: Logan, UT, USA, 2007; Volume 80, pp. 19–51. [Google Scholar]
- Hoffmann, B.D.; Griffiths, A.D.; Andersen, A.N. Response of ant communities to dry sulphur deposition from mining emissions in semi-arid northern Australia, with implications for the use of functional groups. Austral Ecol. 2000, 25, 653–663. [Google Scholar] [CrossRef]
- Barrow, L.; Parr, C.L.; Kohen, J.L. Biogeography and diversity of ants in Purnululu (Bungle Bungle) National Park and conservation reserve. Aust. J. Zool. 2006, 54, 123–136. [Google Scholar] [CrossRef]
- Andersen, A.N.; Hayward, J. Ant Survey of Uluru National Park; Report to Parks Australia; CSIRO Darwin: Berrimah, Australia, 2012. [Google Scholar]
- Gosper, C.R.; Pettit, M.J.; Andersen, A.N.; Yates, C.J.; Prober, S.M. Multi-century dynamics of ant communities following fire in Mediterranean-climate woodlands of south-western Australia: Are changes congruent with vegetation succession? For. Ecol. Manag. 2015, 342, 30–38. [Google Scholar] [CrossRef]
- Andersen, A.N. Ant megadiversity and its origins in arid Australia. Austral Entomol. 2016, 55, 132–147. [Google Scholar] [CrossRef]
- Andersen, A.N.; Arnan, X.; Sparks, K. Limited niche differentiation within remarkable co-occurrences of congeneric species: Monomorium ants in the Australian seasonal tropics. Austral Ecol. 2013, 38, 557–567. [Google Scholar] [CrossRef]
- Sparks, K.S.; Andersen, A.N.; Donnellan, S.C.; Austin, A.D. Navigating the mtDNA road map out of the morphological maze: Interpreting morphological variation in the inscrutably diverse Monomorium rothsteini (Forel) (Hymenoptera: Formicidae). Syst. Entomol. 2014, 39, 264–278. [Google Scholar] [CrossRef]
- Paknia, O.; Bergmann, T.; Hadrys, H. Some ‘ant’swers: Application of a layered barcode approach to problems in ant taxonomy. Mol. Ecol. Resour. 2015, 15, 1262–1274. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A.N.; Hoffmann, B.D.; Berman, M. Diversity in the Australian ant genus Iridomyrmex Mayr, 1862 (Hymenoptera: Formicidae): A critique of Heterick & Shattuck (2011), with particular reference to I. coeruleus Heterick & Shattuck, 2011. Myrmecol. News 2013, 18, 103–111. [Google Scholar]
- CCDB: Canadian Center for DNA Barcoding. Available online: http://ccdb.ca/resources (accessed on 7 December 2016).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. 1990, 41, 95–98. [Google Scholar]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. Partitionfinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- CIPRES: Cyperinfrastructure for Phylogenetic Research. Available online: http://www.phylo.org/sub_sections/portal (accessed on 7 December 2016).
- BEAST. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 7 December 2016).
- Smith, M.A.; Fisher, B.L.; Hebert, P.D.N. DNA barcoding for effective biodiversity assessment of a hyperdiverse arthropod group: The ants of Madagascar. Philos. Trans. R. Soc. B 2005, 360, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Wild, A.L. Evolution of the Neotropical ant genus Linepithema. Syst. Entomol. 2009, 34, 49–62. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed]
- Brower, A.V.Z.; DeSalle, R. Practical and theoretical considerations for choice of a DNA sequence region in insect molecular systematics, with a short review of published studies using nuclear gene regions. Ann. Entomol. Soc. Am. 1994, 87, 702–716. [Google Scholar] [CrossRef]
- Lin, C.-P.; Danforth, B.N. How do insect nuclear and mitochondrial gene substitution patterns differ? Insights from Bayesian analyses of combined datasets. Mol. Phylogenet. Evol. 2004, 30, 686–702. [Google Scholar] [CrossRef]
- Schlick-Steiner, B.C.; Steiner, F.M.; Moder, K.; Seifert, B.; Sanetra, M.; Dyreson, E.; Stau, C.; Erhard, C. A multidisciplinary approach reveals cryptic diversity in Western Palearctic Tetramorium ants (Hymenoptera: Formicidae). Mol. Phylogenet. Evol. 2006, 40, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Low, V.L.; Sofian-Azirun, M.; Norma-Rashid, Y. Playing hide-and-seek with the tiny dragonfly: DHA barcoding discriminates multiple lineages of Nannophya pygmaea in Asia. J. Insect Conserv. 2016, 20, 339–343. [Google Scholar] [CrossRef]
- Kodandaramaiah, U.; Simonsen, T.J.; Bromilow, S.; Wahlberg, N.; Sperling, F. Deceptive single-locus taxonomy and phylogeography: Wolbachia-associated divergence in mitochondrial DNA is not reflected in morphology and nuclear markers in a butterfly species. Ecol. Evol. 2013, 16, 5167–5176. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.P.; Harris, J.L.; Roe, K.J. Incongruence between mtDNA and nuclear data in the freshwater mussel genus Cyprogenia (Bivalvia: Unionidae) and its impact on species delineation. Ecol. Evol. 2016, 6, 2439–2452. [Google Scholar] [CrossRef] [PubMed]
- Knadena, M.; Tinautb, A.; Cerda, X.; Wehnera, S.; Wehner, R. Phylogeny of three parapatric species of desert ants, Cataglyphis bicolor, C. viatica, and C. savignyi: A comparison of mitochondrial DNA, nuclear DNA, and morphological data. Zoology 2005, 108, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, E.O. Pheidole in the New World: A Dominant, Hyperdiverse Ant Genus; Harvard University Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Bacci, M., Jr.; Scott, E.; Solomon, S.E.; Mueller, U.G.; Martins, V.G.; Carvalho, A.O.R.; Vieira, L.G.E.; Silva-Pinhati, A.C.O. Phylogeny of leafcutter ants in the genus Atta Fabricius (Formicidae: Attini) based on mitochondrial and nuclear DNA sequences. Mol. Phylogenet. Evol. 2009, 51, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A.N. Ant diversity in arid Australia: A systematic overview. Mem. Am. Entomol. Inst. 2007, 80, 19–51. [Google Scholar]











© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andersen, A.N.; Hoffmann, B.D.; Sparks, K. The Megadiverse Australian Ant Genus Melophorus: Using CO1 Barcoding to Assess Species Richness. Diversity 2016, 8, 30. https://0-doi-org.brum.beds.ac.uk/10.3390/d8040030
Andersen AN, Hoffmann BD, Sparks K. The Megadiverse Australian Ant Genus Melophorus: Using CO1 Barcoding to Assess Species Richness. Diversity. 2016; 8(4):30. https://0-doi-org.brum.beds.ac.uk/10.3390/d8040030
Chicago/Turabian StyleAndersen, Alan N., Benjamin D. Hoffmann, and Kathryn Sparks. 2016. "The Megadiverse Australian Ant Genus Melophorus: Using CO1 Barcoding to Assess Species Richness" Diversity 8, no. 4: 30. https://0-doi-org.brum.beds.ac.uk/10.3390/d8040030