
Potential Enhanced Survivorship of Crown of Thorns Starfish Larvae due to Near-Annual Nutrient Enrichment during Secondary Outbreaks on the Central Mid-Shelf of the Great Barrier Reef, Australia



Abstract
:1. Introduction
2. Mortality of COTS at Different Life History Stages
3. Conceptual Understanding of the Nutrient Enrichment-CoTS Link
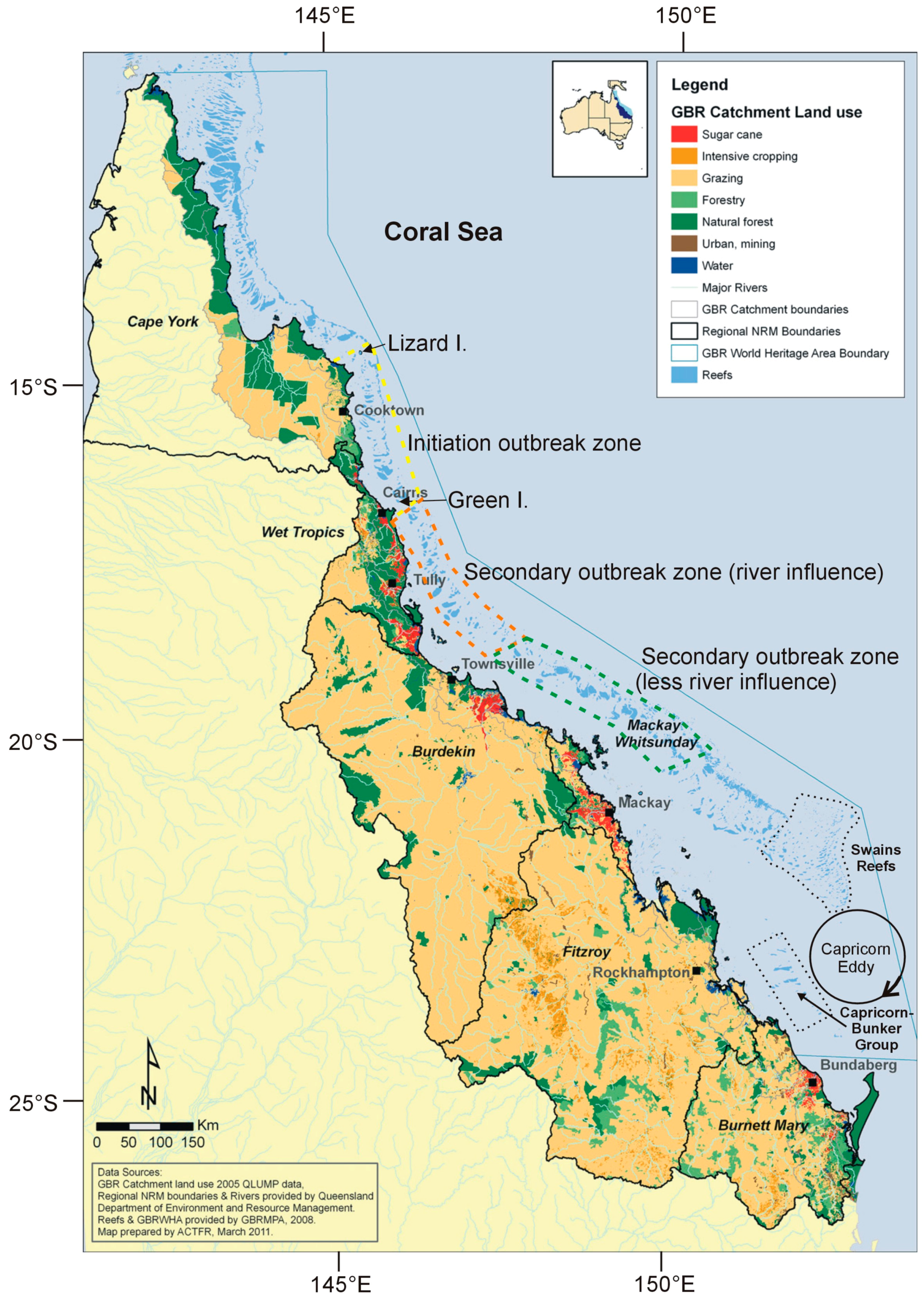
4. Crown of Thorns Starfish and the Great Barrier Reef
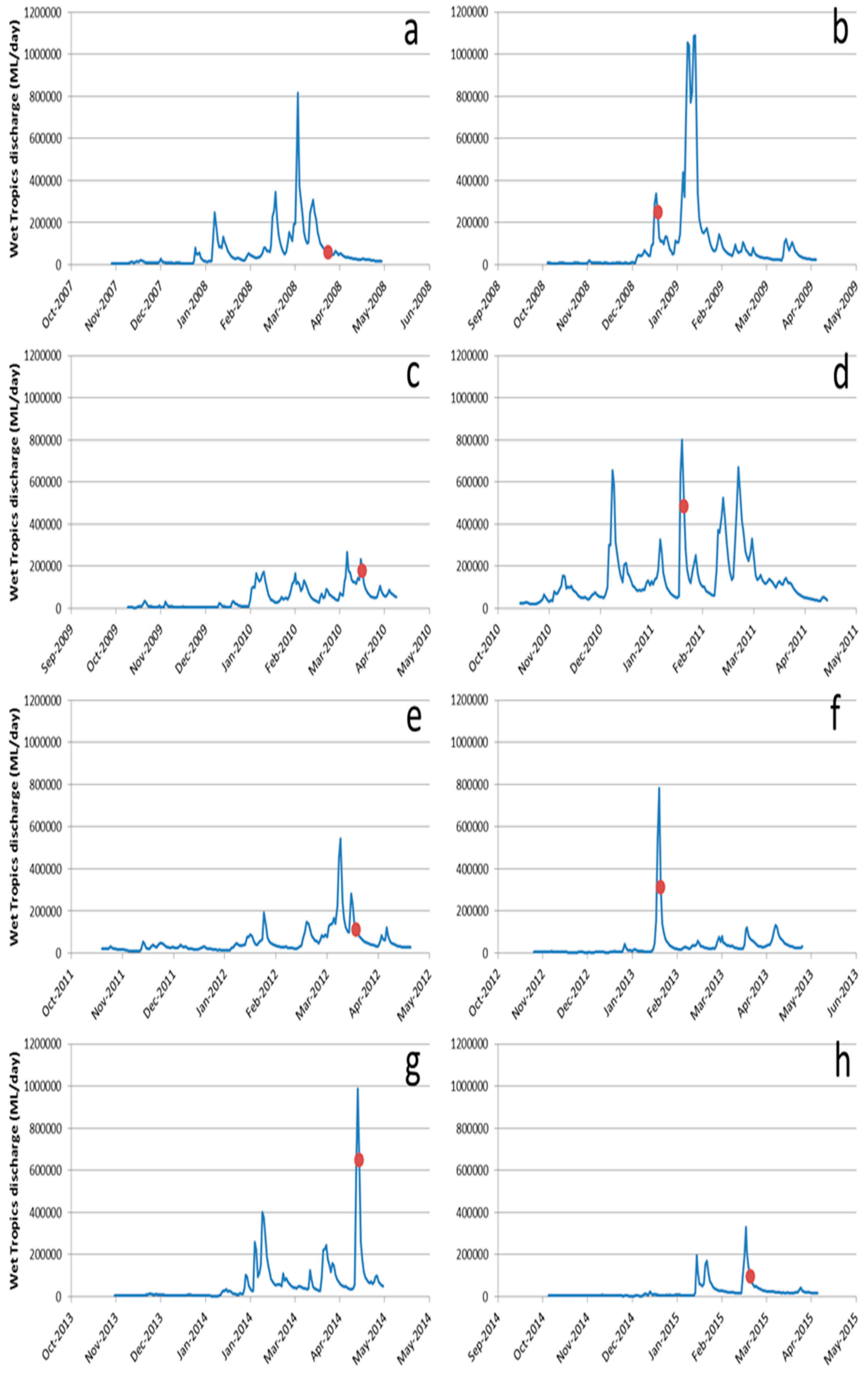
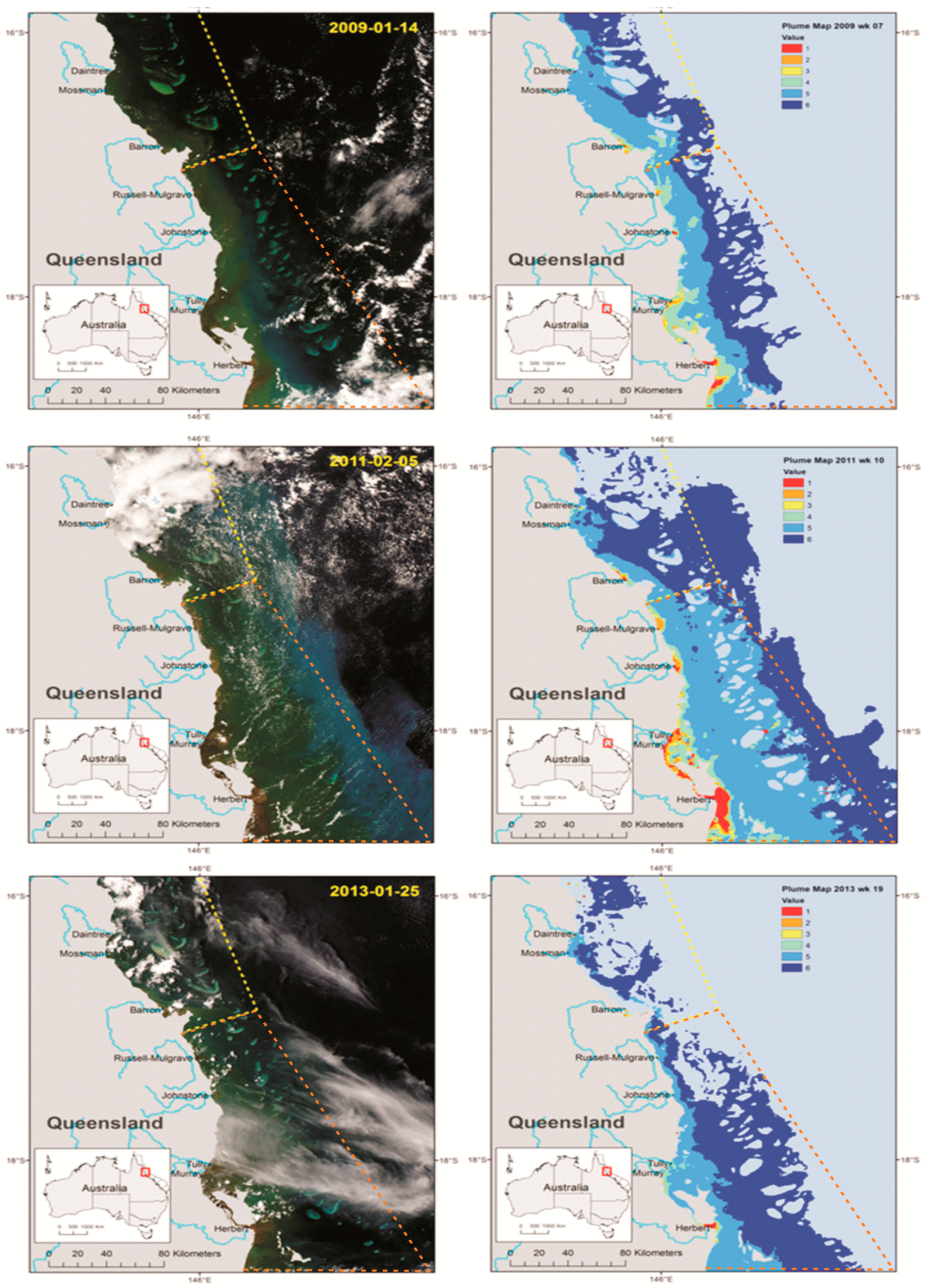
5. Nutrient Enrichment and Secondary Outbreaks
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Birkeland, C.; Lucas, J.S. Acanthaster planci: Major Management Problem of Coral Reefs; CRC Press, Inc.: Boca Raton, FL, USA, 1990. [Google Scholar]
- Moran, P.J. The Acanthaster phenomenon. Oceanogr. Mar. Biol. Ann. Rev. 1986, 24, 379–480. [Google Scholar]
- Zann, L.; Brodie, J.; Berryman, C.; Naqasima, M. Recruitment, ecology, growth and behavior of juvenile Acanthaster planci (L.)(Echinodermata: Asteroidea). Bull. Mar. Sci. 1987, 41, 561–575. [Google Scholar]
- Zann, L.; Brodie, J.; Vuki, V. History and dynamics of the crown-of-thorns starfish Acanthaster planci (L.) in the Suva area, Fiji. Coral Reef. 1990, 9, 135–144. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Caballes, C.F.; Rivera-Posada, J.A.; Sweatman, H.P.A. Limits to understanding and managing outbreaks of crown-of-thorns starfish (Acanthaster spp.). Oceanogr. Mar. Biol. Ann. Rev. 2014, 52, 133–200. [Google Scholar]
- Brodie, J.E. Enhancement of larval and juvenile survival and recruitment in Acanthaster planci from the effects of terrestrial runoff: A review. Aust. J. Mar. Freshw. Res. 1992, 43, 539–554. [Google Scholar] [CrossRef]
- Caballes, C.F.; Pratchett, M.S. Reproductive biology and early life history of the crown-of-thorns starfish. In Echinoderms: Ecology, Habitats and Reproductive Biology; Eric, W., Ed.; Nova Science Publishers: New York, NY, USA, 2014; pp. 101–146. [Google Scholar]
- Sweatman, H.P.A. A field study of fish predation on juvenile crown of- thorns starfish. Coral Reefs 1995, 14, 47–53. [Google Scholar] [CrossRef]
- Mendonça, V.M.; Al Jabri, M.M.; Al Ajmi, I.; Al Muharrami, M.; Al Areimi, M.; Al Aghbari, H.A. Starfish Acanthaster planci in the Northwestern Indian Ocean: Are they really a consequence of unsustainable starfish predator removal through overfishing in coral reefs, or a response to a changing environment? Zool. Stud. 2010, 49, 108–123. [Google Scholar]
- Lucas, J.S. Quantitative studies of feeding and nutrition during larval development of the coral reef asteroid Acanthaster planci (L.). J. Exp. Mar. Biol. Ecol. 1982, 65, 173–193. [Google Scholar] [CrossRef]
- Brodie, J.; Fabricius, K.; De’ath, G.; Okaji, K. Are increased nutrient inputs responsible for more outbreaks of crown-of-thorns starfish? An appraisal of the evidence. Mar. Pollut. Bull. 2005, 51, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Fabricius, K.E.; Okaji, K.; De’ath, G. Three lines of evidence to link outbreaks of the crown-of-thorns seastar Acanthaster planci to the release of larval food limitation. Coral Reefs 2010, 29, 593–605. [Google Scholar] [CrossRef]
- Potts, D.C. Crown-of-thorns starfish—Man-induced pest or natural phenomenon. In the Ecology of Pests; Commonwealth Scientific and Industrial Research Organization: Melbourne, Australia, 1981; pp. 55–86. [Google Scholar]
- Chesher, R.H. Destruction of the Pacific Corals by the Sea Star Acanthaster planci. Science 1969, 165, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Randall, J.E. Chemical pollution in the sea and the crown-of-thorns starfish (Acanthaster planci). Biotropica 1972, 4, 132–144. [Google Scholar] [CrossRef]
- Wooldridge, S.; Brodie, J. Environmental triggers for primary outbreaks of crown-of-thorns starfish on the Great Barrier Reef, Australia. Mar. Pollut. Bull. 2015, 101, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Hock, K.; Wolff, N.H.; Condie, S.A.; Anthony, K.; Mumby, P.J. Connectivity networks reveal the risks of crown-of-thorns starfish outbreaks on the Great Barrier Reef. J. Appl. Ecol. 2014, 51, 1188–1196. [Google Scholar] [CrossRef]
- Uthicke, S.; Doyle, J.; Duggan, S.; Yasuda, N.; McKinnon, A.D. Outbreak of coral-eating Crown-of-Thorns creates continuous cloud of larvae over 320 km of the Great Barrier Reef. Sci. Rep. 2015, 5, 16885. [Google Scholar] [CrossRef] [PubMed]
- Keesing, J.K.; Halford, A.R. Importance of post settlement processes for the population dynamics of Acanthaster planci (L.). Mar. Freshw. Res. 1992, 43, 635–651. [Google Scholar] [CrossRef]
- Keesing, J.K.; Halford, A.R. Field measurement of survival rates of juvenile Acanthaster planci: Techniques and preliminary results. Mar. Ecol. Prog. Ser. 1992, 85, 107–114. [Google Scholar] [CrossRef]
- Keesing, J.K.; Wiedermeyer, W.L.; Okaji, K.; Halford, A.R.; Hall, K.C.; Cartwright, C.M. Mortality rates of juvenile starfish Acanthaster planci and Nardoa spp. measured on the Great Barrier Reef, Australia and in Okinawa, Japan. Oceanol. Acta 1996, 19, 441–448. [Google Scholar]
- Ayukai, T. Ingestion of ultraplankton by the planktonic larvae of the crown-of-thorns starfish Acanthaster planci. Biol. Bull. 1994, 186, 90–100. [Google Scholar] [CrossRef]
- Okaji, K.; Ayukai, T.; Lucas, J. Selective feeding by larvae of the crown-of-thorns starfish, Acanthaster planci (L.). Coral Reefs 1997, 16, 47–50. [Google Scholar] [CrossRef]
- Okaji, K.; Ayukai, T.; Lucas, J. Are Acanthaster planci larvae food limited in the Great Barrier Reef waters. In Proceedings of the 8th International Coral Reef Symposium, Panama, Panama, 24–29 June 1996.
- Cowan, Z.L.; Dworjanyn, S.A.; Caballes, C.F.; Pratchett, M.S. Predation on crown-of-thorns starfish larvae by damselfishes. Coral Reefs 2016, 35, 1253–1262. [Google Scholar] [CrossRef]
- Birkeland, C. Acanthaster in the cultures of high islands. Atoll Res. Bull. 1981, 255, 55–58. [Google Scholar]
- Birkeland, C. Terrestrial runoff as a cause of outbreaks of Acanthaster planci (Echinodermata: Asteroidea). Mar. Biol. 1982, 69, 175–185. [Google Scholar] [CrossRef]
- Brodie, J.E.; Devlin, M.; Haynes, D.; Waterhouse, J. Assessment of the eutrophication status of the Great Barrier Reef lagoon (Australia). Biogeochemistry 2011, 106, 281–302. [Google Scholar] [CrossRef]
- Mellin, C.; Lugrin, C.; Okaji, K.; Francis, D.S.; Uthicke, S. Selective feeding and microalgal consumption rates by Crown-of-Thorns Seastar (Acanthaster cf. solaris) larvae. Diversity 2017, 9, 8. [Google Scholar] [CrossRef]
- Henderson, J.A.; Lucas, J.S. Larval development and metamorphosis of Acanthaster planci (Asteroidea). Nature 1971, 232, 655–657. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.S. Reproductive and larval biology of Acanthaster planci (L.) in Great Barrier Reef waters. Micronesica 1973, 9, 197–203. [Google Scholar]
- Lucas, J.S. Growth, maturation and effects of diet in Acanthaster planci (L.)(Asteroidea) and hybrids reared in the laboratory. J. Exp. Mar. Biol. Ecol. 1984, 79, 129–147. [Google Scholar] [CrossRef]
- Olson, R.R. In situ culturing as a test of the larval starvation hypothesis for the crown-of-thorns starfish, Acanthaster planci. Limnol. Oceanogr. 1987, 32, 895–904. [Google Scholar] [CrossRef]
- Olson, R.R.; Olson, M.H. Food limitation of planktotrophic marine invertebrate larvae: Does it control recruitment success? Ann. Rev. Ecol. Syst. 1989, 20, 225–247. [Google Scholar] [CrossRef]
- Ayukai, T.; Okaji, K.; Lucas, J.S. Food limitation in the growth and development of crown-of-thorns starfish in the Great Barrier Reef. In Proceedings of the 8th International Coral Reef Symposium, Panama, Panama, 24–29 June 1996; pp. 621–626.
- Fabricius, K.E. Factors determining the resilience of coral reefs to eutrophication: A review and conceptual model. In Coral Reefs: An Ecosystem in Transition; Dubinski, Z., Stambler, N., Eds.; Springer: New York, NY, USA, 2011; pp. 493–509. [Google Scholar]
- Uthicke, S.; Logan, M.; Liddy, M.; Francis, D.; Hardy, N.; Lamare, M. Climate change as an unexpected co-factor promoting coral eating seastar (Acanthaster planci) outbreaks. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.; Graba-Landry, A.; Dworjanyn, S.A.; Byrne, M. Larval starvation to satiation: Influence of nutrient regime on the success of Acanthaster planci. PLoS ONE 2015, 10, e0122010. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.; Graba-Landry, A.; Dworjanyn, S.A.; Byrne, M. Larval phenotypic plasticity in the boom-and-bust crown-of-thorns seastar, Acanthaster planci. Mar. Ecol. Prog. Ser. 2015, 539, 179–189. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Dworjanyn, S.; Mos, B.; Caballes, C.F.; Thompson, C.A.; Blowes, S. Larval Survivorship and Settlement of Crown-of-Thorns Starfish (Acanthaster cf. solaris) at Varying Algal Cell Densities. Diversity 2017, 9, 2. [Google Scholar] [CrossRef]
- Brodie, J.; De’ath, G.; Devlin, M.; Furnas, M.; Wright, M. Spatial and temporal patterns of near-surface chlorophyll a in the Great Barrier Reef lagoon. Mar. Freshw. Res. 2007, 58, 342–353. [Google Scholar] [CrossRef]
- Devlin, M.J.; Brodie, J. Terrestrial discharge into the Great Barrier Reef Lagoon: Nutrient behaviour in coastal waters. Mar. Pollut. Bull. 2005, 51, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.; Schaffelke, B. Spatial extent of riverine flood plumes and exposure of marine ecosystems in the Tully coastal region, Great Barrier Reef. Mar. Freshw. Res. 2009, 60, 1109–1122. [Google Scholar] [CrossRef]
- Devlin, M.J.; Da Silva, E.T.; Petus, C.; Wenger, A.; Zeh, D.; Tracey, D.; Álvarez-Romero, J.G.; Brodie, J. Combining in-situ water quality and remotely sensed data across spatial and temporal scales to measure variability in wet season chlorophyll-a: Great Barrier Reef lagoon (Queensland, Australia). Ecol. Process. 2013, 2, 1–22. [Google Scholar] [CrossRef]
- Schroeder, T.; Devlin, M.J.; Brando, V.E.; Dekker, A.G.; Brodie, J.E.; Clementson, L.A.; McKinna, L. Inter-annual variability of wet season freshwater plume extent into the Great Barrier Reef lagoon based on satellite coastal ocean colour observations. Mar. Pollut. Bull. 2012, 65, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, Z.T.; Wolanski, E.; Álvarez-Romero, J.G.; Lewis, S.E.; Brodie, J.E. Fine sediment and nutrient dynamics related to particle size and floc formation in a Burdekin River flood plume, Australia. Mar. Pollut. Bull. 2012, 65, 236–248. [Google Scholar] [CrossRef] [PubMed]
- De’ath, G.; Fabricius, K.E.; Sweatman, H.; Puotinen, M. The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl. Acad. Sci. USA 2012, 109, 17995–17999. [Google Scholar] [CrossRef] [PubMed]
- Walbran, P.D.; Henderson, R.A.; Faithful, J.W.; Polach, H.A.; Sparks, R.J.; Wallace, G.; Lowe, D.C. Crown of thorns starfish outbreaks on the Great Barrier Reef: A geological perspective based upon the geological record. Coral Reefs 1989, 8, 67–78. [Google Scholar] [CrossRef]
- Hock, K.; Wolff, N.H.; Beeden, R.; Hoey, J.; Condie, S.A.; Anthony, K.; Possingham, H.P.; Mumby, P.I. Controlling range expansion in habitat networks by adaptively targeting source populations. Conserv. Biol. 2016, 30, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Miller, I.; Sweatman, H.; Cheal, A.; Emslie, M.; Johns, K.; Jonker, M.; Osborne, K. Origins and implications of a primary crown-of-thorns starfish outbreak in the southern Great Barrier Reef. J. Mar. Biol. 2015, 2015, 809624. [Google Scholar] [CrossRef]
- Roche, R.C.; Pratchett, M.S.; Carr, P.; Turner, J.R.; Wagner, D.; Head, C.; Sheppard, C.R.C. Localized outbreaks of Acanthaster planci at an isolated and unpopulated reef atoll in the Chagos Archipelago. Mar. Biol. 2015, 162, 1695–1704. [Google Scholar] [CrossRef]
- Houk, P.; Raubani, J. Acanthaster planci outbreaks in Vanuatu coincide with ocean productivity, furthering trends throughout the Pacific Ocean. J. Oceanogr. 2010, 66, 435–438. [Google Scholar] [CrossRef]
- Houk, P.; Bograd, S.; van Woesik, R. The transition zone chlorophyll front can trigger Acanthaster planci outbreaks in the Pacific Ocean: Historical confirmation. J. Oceanogr. 2007, 63, 149–154. [Google Scholar] [CrossRef]
- Weeks, S.; Bakun, A.; Steinberg, C.; Brinkman, R.; Hoegh-Guldberg, O. The Capricorn Eddy: A prominent driver of the ecology and future of the southern Great Barrier Reef. Coral Reefs 2010, 29, 975–985. [Google Scholar] [CrossRef]
- Mao, Y.; Luick, J.L. Circulation in the southern Great Barrier Reef studied through an integration of multiple remote sensing and in situ measurements. J. Geophys. Res. 2014, 119, 1621–1643. [Google Scholar] [CrossRef]
- Condie, S.; Condie, R. Retention of plankton within ocean eddies. Glob. Ecol. Biogeogr. 2016, 25, 1264–1277. [Google Scholar] [CrossRef]
- Weeks, S.J.; Magno-Canto, M.M.; Jaine, F.R.A.; Brodie, J.; Richardson, A.J. Unique sequence of events triggers manta ray feeding frenzy in the southern Great Barrier Reef, Australia. Remote Sens. 2015, 7, 3138–3152. [Google Scholar] [CrossRef]
- Alongi, D.M.; Patten, N.L.; McKinnon, D.; Köstner, N.; Bourne, D.G.; Brinkman, R. Phytoplankton, bacterioplankton and virioplankton structure and function across the southern Great Barrier Reef shelf. J. Mar. Syst. 2015, 142, 25–39. [Google Scholar] [CrossRef]
- Da Silva, E.T.; Devlin, M.; Wenger, A.; Petus, C. Burnett-Mary Wet Season 2012–2013: Water Quality Data Sampling, Analysis and Comparison against Wet Season 2010–2011 Data.; Centre for Tropical Water & Aquatic Ecosystem Research (TropWATER) Publication: James Cook University, Townsville, Australia, 2013; p. 31. [Google Scholar]
- Brodie, J.E.; Mitchell, A.W. Nutrient composition of the January (1991) fitzroy river flood plume. In Workshop on the Impacts of Flooding; GBRMPA Workshop Series No. 17; Byron, G.T., Ed.; Great Barrier Reef Marine Park Authority: Townsville, Australia, 1992; pp. 56–74. [Google Scholar]
- Kroon, F.J.; Kuhnert, P.M.; Henderson, B.L.; Wilkinson, S.N.; Kinsey-Henderson, A.; Abbott, B.; Brodie, J.E.; Turner, R.D.R. River loads of suspended solids, nitrogen, phosphorus and herbicides delivered to the Great Barrier Reef lagoon. Mar. Pollut. Bull. 2012, 65, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, J.; Brodie, J.; Lewis, S.; Mitchell, A. Quantifying the sources of pollutants in the Great Barrier Reef catchments and the relative risk to reef ecosystems. Mar. Pollut. Bull. 2012, 65, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Furnas, M.; Mitchell, A.; Skuza, M.; Brodie, J. In the other 90%: Phytoplankton responses to enhanced nutrient availability in the Great Barrier Reef Lagoon. Mar. Pollut. Bull. 2005, 51, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.J.; McKinna, L.W.; Álvarez-Romero, J.G.; Petus, C.; Abott, B.; Harkness, P.; Brodie, J. Mapping the pollutants in surface riverine flood plume waters in the Great Barrier Reef, Australia. Mar. Pollut. Bull. 2012, 65, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Romero, J.G.; Devlin, M.; Teixeira da Silva, E.; Petus, C.; Ban, N.C.; Pressey, R.L.; Kool, J.; Roberts, J.J.; Cerdeira-Estrada, S.; Wenger, A.S.; et al. A novel approach to model exposure of coastal-marine ecosystems to riverine flood plumes based on remote sensing techniques. J. Environ. Manag. 2013, 119, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Blondeau-Patissier, D.; Schroeder, T.; Brando, V.E.; Maier, S.W.; Dekker, A.G.; Phinn, S. ESA-MERIS 10-year mission reveals contrasting phytoplankton bloom dynamics in two tropical regions of Northern Australia. Remote Sens. 2014, 6, 2963–2988. [Google Scholar] [CrossRef]
- Devlin, M.; Petus, C.; da Silva, E.; Tracey, D.; Wolff, N.; Waterhouse, J.; Brodie, J. Water quality and river plume monitoring in the Great Barrier Reef: An overview of methods based on ocean colour satellite data. Remote Sens. 2015, 7, 12909–12941. [Google Scholar] [CrossRef]
- Devlin, M.; Waterhouse, J.; Taylor, J.; Brodie, J. Flood Plumes in the Great Barrier Reef: Spatial and Temporal Patterns in Composition and Distribution; GBRMPA Research Publication No. 68; Great Barrier Reef Marine Park Authority: Townsville, Australia, 2001. [Google Scholar]
- Petus, C.; Collier, C.; Devlin, M.; Rasheed, M.; McKenna, S. Using MODIS data for understanding changes in seagrass meadow health: A case study in the Great Barrier Reef (Australia). Mar. Environ. Res. 2014, 98, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Petus, C.; Devlin, M.; Thompson, A.; McKenzie, L.; Teixeira da Silva, E.; Collier, C.; Tracey, D.; Martin, K. Estimating the Exposure of Coral Reefs and Seagrass Meadows to Land-Sourced Contaminants in River Flood Plumes of the Great Barrier Reef: Validating a Simple Satellite Risk Framework with Environmental Data. Remote Sens. 2016, 8, 210. [Google Scholar] [CrossRef]
- Waterhouse, J.; Brodie, J.; Petus, C.; Devlin, M.; da Silva, E.; Maynard, J.; Heron, S.; Tracey, D. Recent Findings of an Assessment of Remote Sensing Data for Water Quality Measurement in the Great Barrier Reef: Supporting Information for the GBR Water Quality Improvement Plans; Centre for Tropical Water & Aquatic Ecosystem Research (TropWATER) Publication: James Cook University, Townsville, Australia, 2015; p. 71. [Google Scholar]
- Wolfe, K.; Graba-Landry, A.; Dworjanyn, S.A.; Byrne, M. Superstars: Assessing nutrient thresholds for enhanced larval success of Acanthaster planci, a review of the evidence. Mar. Pollut. Bull. 2017, in press. [Google Scholar] [CrossRef] [PubMed]
- Caballes, C.F.; Pratchett, M.S.; Kerr, A.M.; Rivera-Posada, J.A. The role of maternal nutrition on oocyte size and quality, with respect to early larval development in the coral-eating starfish, Acanthaster planci. PLoS ONE 2016, 11, e0158007. [Google Scholar] [CrossRef] [PubMed]
- Babcock, R.C.; Dambacher, J.M.; Morello, E.B.; Plaganyi, É.E.; Hayes, K.R.; Sweatman, H.P.A.; Pratchett, M.S. Assessing Different Causes of Crown-of-Thorns Starfish Outbreaks and Appropriate Responses for Management on the Great Barrier Reef. PLoS ONE 2016, 11, e0169048. [Google Scholar] [CrossRef] [PubMed]
- Baird, M.E.; Cherukuru, N.; Jones, E.; Margvelashvili, N.; Mongin, M.; Oubelkheir, K.; Ralph, P.J.; Rizwi, F.; Robson, B.J.; Schroeder, T.; et al. Remote-sensing reflectance and true colour produced by a coupled hydrodynamic, optical, sediment, biogeochemical model of the Great Barrier Reef, Australia: Comparison with satellite data. Environ. Model. Softw. 2016, 78, 79–96. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Type and Class | Mean Chlorophyll a (μg·L−1) | Range Chlorophyll a (μg·L−1) |
|---|---|---|
| Primary, Colour Classes one–four | 0.9 | 0.6–3.7 |
| Secondary, Colour Class five | 1.3 | 0.7–2.7 |
| Tertiary, Colour Class six | 0.7 | 0.2–1.6 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brodie, J.; Devlin, M.; Lewis, S. Potential Enhanced Survivorship of Crown of Thorns Starfish Larvae due to Near-Annual Nutrient Enrichment during Secondary Outbreaks on the Central Mid-Shelf of the Great Barrier Reef, Australia. Diversity 2017, 9, 17. https://0-doi-org.brum.beds.ac.uk/10.3390/d9010017
Brodie J, Devlin M, Lewis S. Potential Enhanced Survivorship of Crown of Thorns Starfish Larvae due to Near-Annual Nutrient Enrichment during Secondary Outbreaks on the Central Mid-Shelf of the Great Barrier Reef, Australia. Diversity. 2017; 9(1):17. https://0-doi-org.brum.beds.ac.uk/10.3390/d9010017
Chicago/Turabian StyleBrodie, Jon, Michelle Devlin, and Stephen Lewis. 2017. "Potential Enhanced Survivorship of Crown of Thorns Starfish Larvae due to Near-Annual Nutrient Enrichment during Secondary Outbreaks on the Central Mid-Shelf of the Great Barrier Reef, Australia" Diversity 9, no. 1: 17. https://0-doi-org.brum.beds.ac.uk/10.3390/d9010017