Millennia-Long Co-Existence of Two Major European Whitefish (Coregonus spp.) Lineages in Switzerland Inferred from Ancient Mitochondrial DNA

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
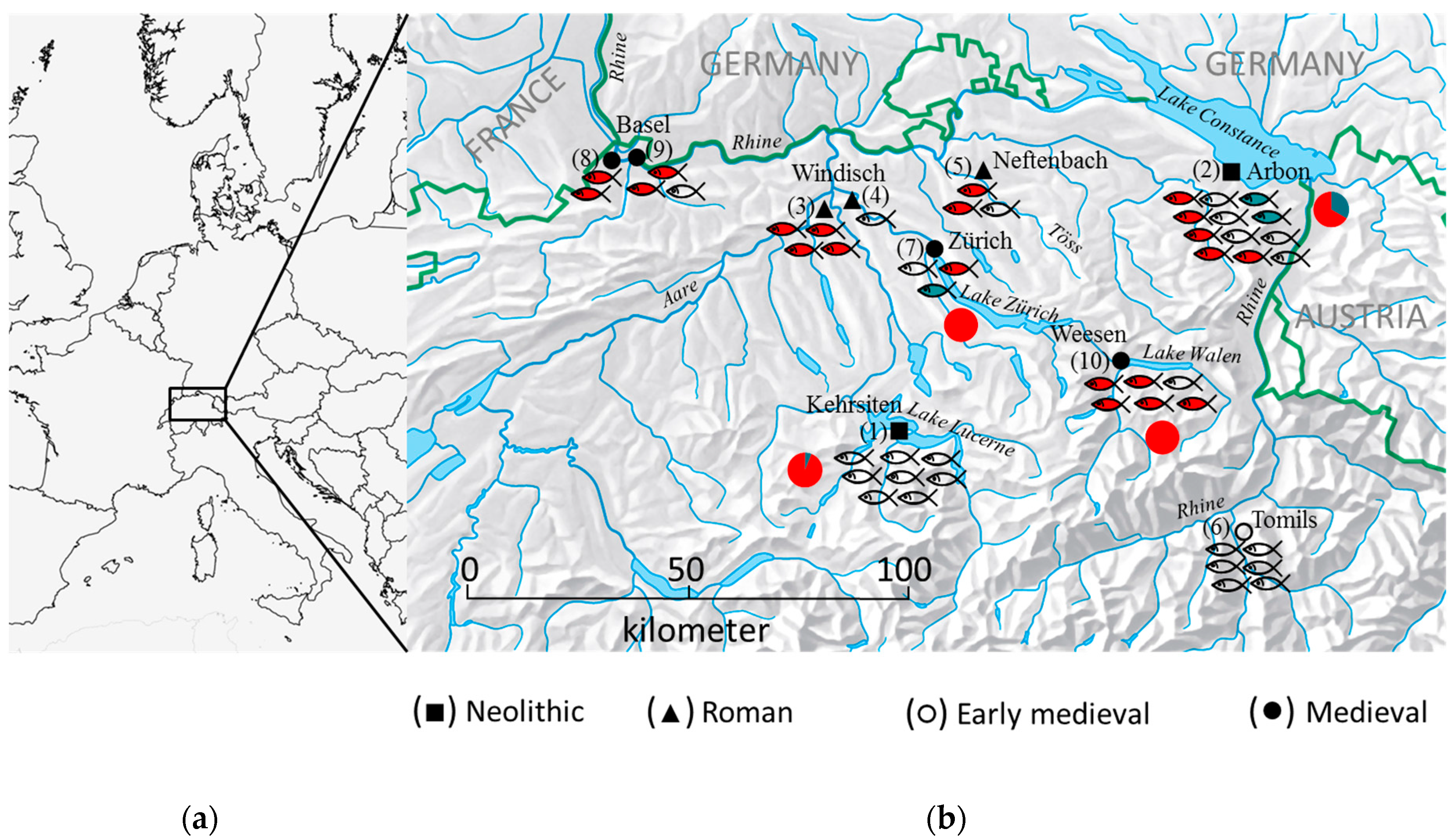
2.2. Archaeological Sites
2.3. Neolithic Period
2.3.1. Stansstad-Kehrsiten
2.3.2. Arbon-Bleiche 3
2.4. Roman Era
2.5. Medieval Period
2.6. Methods
2.6.1. DNA Extraction
2.6.2. Primer Design, PCR, and Sequencing
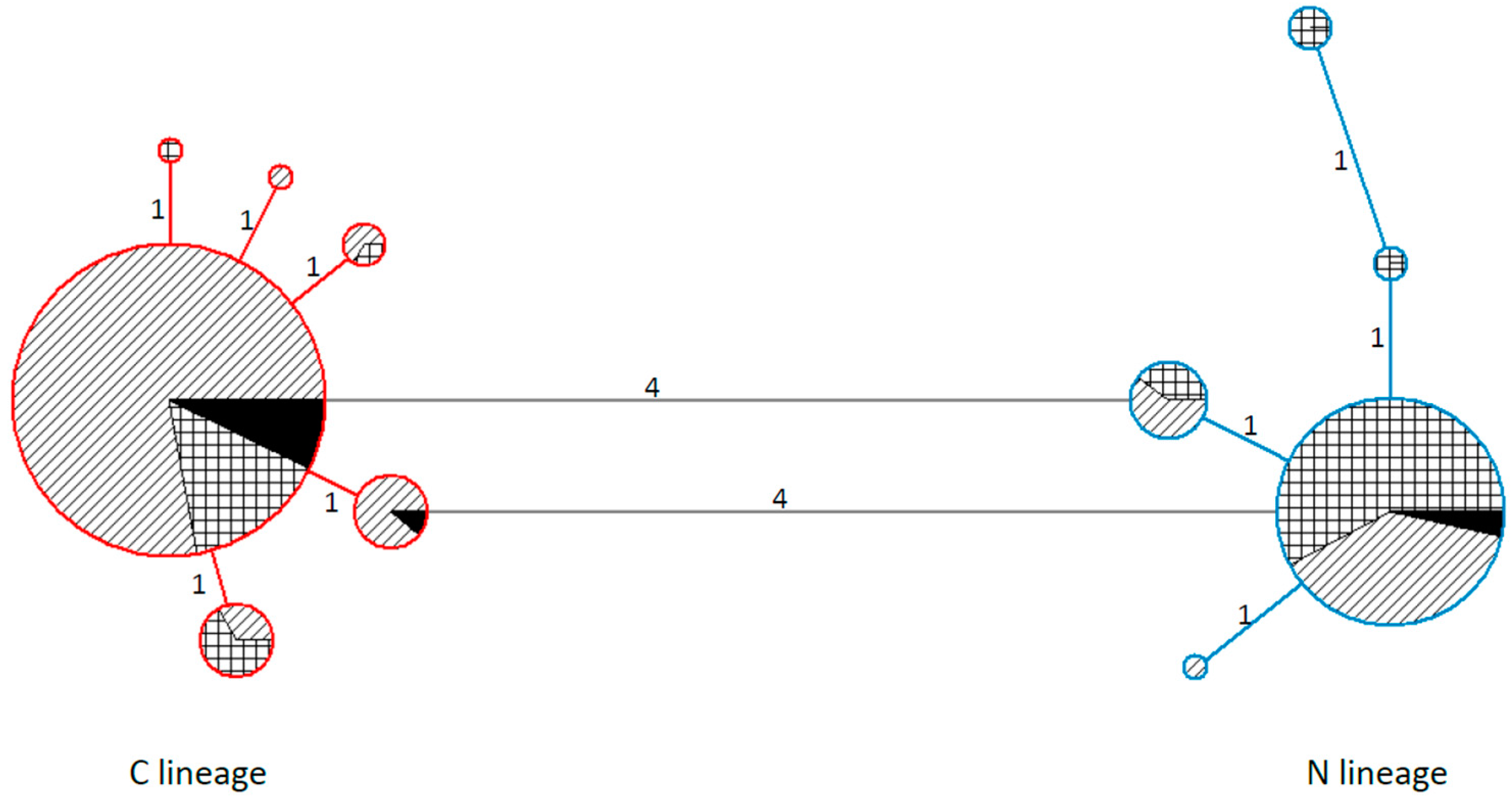
3. Results and Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bernatchez, L.; Renaut, S.; Whiteley, A.R.; Derome, N.; Jeukens, J.; Landry, L.; Lu, G.; Nolte, A.W.; Ostbye, K.; Rogers, S.M.; et al. On the origin of species: Insights from the ecological genomics of lake whitefish. Philos. Trans. R. Soc. B 2010, 365, 1783–1800. [Google Scholar] [CrossRef] [PubMed]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Publications Kottelat: Cornol, Switzerland, 2007. [Google Scholar]
- Schulz, M.; Freyhof, J.; Saint-Laurent, R.; Østbye, K.; Mehner, T.; Bernatchez, L. Evidence for independent origin of two spring-spawning ciscoes (salmoniformes: Coregonidae) in germany. J. Fish Biol. 2006, 68, 119–135. [Google Scholar] [CrossRef]
- Bernatchez, L. Ecological theory of adaptive radiation: An empirical assessment from coregonine fishes (salmoniformes). In Evolution Illuminated: Salmon and Their Relatives; Hendry, A.P., Stearns, S.C., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 175–207. [Google Scholar]
- Hudson, A.G.; Vonlanthen, P.; Mueller, R.; Seehausen, O. Review: The geography of speciation and adaptive radiation in coregonines. In Biology and Management of Coregonid Fishes—2005; Jankun, M., Brzuzan, P., Hliwa, P., Luczynski, M., Eds.; Schweizerbart Wissenschaftsverlage: Stuttgart, Germany, 2007; Volume 60, pp. 111–146. [Google Scholar]
- Østbye, K.; Amundsen, P.A.; Bernatchez, L.; Klemetsen, A.; Knudsen, R.; Kristoffersen, R.; Næsje, T.F.; Hindar, K. Parallel evolution of ecomorphological traits in the european whitefish Coregonus lavaretus (L.) species complex during postglacial times. Mol. Ecol. 2006, 15, 3983–4001. [Google Scholar] [CrossRef] [PubMed]
- Douglas, M.R.; Brunner, P.C.; Bernatchez, L. Do assemblages of Coregonus (teleostei: Salmoniformes) in the central alpine region of europe represent species flocks? Mol. Ecol. 1999, 8, 589–603. [Google Scholar] [CrossRef]
- Praebel, K.; Knudsen, R.; Siwertsson, A.; Karhunen, M.; Kahilainen, K.K.; Ovaskainen, O.; Ostbye, K.; Peruzzi, S.; Fevolden, S.E.; Amundsen, P.A. Ecological speciation in postglacial european whitefish: Rapid adaptive radiations into the littoral, pelagic, and profundal lake habitats. Ecol. Evol. 2013, 3, 4970–4986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmann, P. Monographie der schweizerischen Koregonen. Beitrag zum Problem der Entstehung neuer Arten. Spezieller Teil. Schweiz Z. Hydrobiol. 1950, 12, 340–491. [Google Scholar]
- Vonlanthen, P.; Bittner, D.; Hudson, A.G.; Young, K.A.; Muller, R.; Lundsgaard-Hansen, B.; Roy, D.; Di Piazza, S.; Largiader, C.R.; Seehausen, O. Eutrophication causes speciation reversal in whitefish adaptive radiations. Nature 2012, 482, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Douglas, M.R.; Brunner, P.C. Biodiversity of central alpine Coregonus (salmoniformes): Impact of one-hundred years of management. Ecol. Appl. 2002, 12, 154–172. [Google Scholar] [CrossRef]
- Hudson, A.G.; Vonlanthen, P.; Seehausen, O. Rapid parallel adaptive radiations from a single hybridogenic ancestral population. Proc. R. Soc. B 2011, 278, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Hudson, A.G.; Lundsgaard-Hansen, B.; Lucek, K.; Vonlanthen, P.; Seehausen, O. Managing cryptic biodiversity: Fine-scale intralacustrine speciation along a benthic gradient in alpine whitefish (Coregonus spp.). Evol. Appl. 2017, 10, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Østbye, K.; Bernatchez, L.; Næsje, T.F.; Himberg, K.J.M.; Hindar, K. Evolutionary history of the european whitefish Coregonus lavaretus (L.) species complex as inferred from mtdna phylogeography and gill-raker numbers. Mol. Ecol. 2005, 14, 4371–4387. [Google Scholar] [CrossRef] [PubMed]
- Alexander, T.J.; Vonlanthen, P.; Seehausen, O. Does eutrophication-driven evolution change aquatic ecosystems? Philos. Trans. R. Soc. B 2017, 372, 20160041. [Google Scholar] [CrossRef] [PubMed]
- Ruhlé, C.; Kindle, T. Morphological comparison of river-spawning whitefish of the alpine rhine with the whitefish of lake constance. Pol. Arch. Hydrobiol. 1992, 39, 403–408. [Google Scholar]
- Rick, T.C.; Lockwood, R. Integrating paleobiology, archeology, and history to inform biological conservation. Conserv. Biol. 2013, 27, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Hüster Plogmann, H. Fischfang und Kleintierbeute. Ergebnisse der Untersuchung aus den Schlämmproben. In Die Jungsteinzeitliche Seeufersiedlung Arbon—Bleiche 3. Umwelt und Wirtschaft. Archäologie im Thurgau 12; Jacomet, S., Leuzinger, U., Schibler, J., Eds.; Amt für Archäologie des Kantons Thurgau: Frauenfeld, Switzerland, 2004; pp. 253–276. [Google Scholar]
- Michel, C.; Bleicher, N.; Brombacher, C.; Hüster Plogmann, H.; Ismail-Meyer, K.; Rehazek, A. Pfahlbauten am Vierwaldstättersee-der steinzeitliche Siedlungsplatz in Kehrsiten. Archäol. Schweiz 2012, 35, 56–71. [Google Scholar]
- Hüster Plogmann, H. Befunde und Fundkomplexe-der steingebaute Gutshof: Knochenreste aus Schlämmproben. In Der Römische Gutshof in Neftenbach. Monographien der Kantonsarchäologie Zürich 31, Band 1; Kantonsarchäologie Zürich, Ed.; Fotorotar AG: Egg/Zürich, Switzerland, 1999; pp. 413–414. [Google Scholar]
- Hüster Plogmann, H. Von Leckerbissen und Schädlingen-die Untersuchung der Kleintierreste. In Zur Frühzeit von Vindonissa. Auswertung der Holzbauten der Grabung Windisch-Breite 1996–1998. Veröffentlichungen der Gesellschaft pro Vindonissa XVIII; Kantonsarchäologie Aargau, Ed. Kantonsarchäologie Aargau: Brugg, Switzerland, 2003; pp. 231–243. [Google Scholar]
- Zaugg, B.; Stucki, P.; Pedroli, J.-C.; Kirchhofer, A. Pisces—Atlas Fauna Helvetica 7; CSCF: Chaumont, France, 2003. [Google Scholar]
- Häberle, S.; Fuller, B.T.; Nehlich, O.; Van Neer, W.; Schibler, J.; Hüster Plogmann, H. Inter- and intraspecies variability in stable isotope ratio values of archaeological freshwater fish remains from switzerland (11th–19th centuries ad). Environ. Archaeol. 2016, 21, 119–132. [Google Scholar] [CrossRef]
- Hüster Plogmann, H. (Ed.) Fisch und Fischer aus zwei Jahrtausenden. Eine Fischereiwirtschaftliche Zeitreise durch die Nordwestschweiz. Forschungen in Augst 39; Römermuseum Augst: Augst, Switzerland, 2006. [Google Scholar]
- Müller, L. Die Fischerei im Spätmittelalterlichen Basel. Lizentiazsarbeit; Universität Basel: Basel, Switzerland, 1989. [Google Scholar]
- Chassaing, O.; Desse-Berset, N.; Hanni, C.; Hughes, S.; Berrebi, P. Phylogeography of the european sturgeon (acipenser sturio): A critically endangered species. Mol. Phylogenet. Evol. 2016, 94, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Grealy, A.; Douglass, K.; Haile, J.; Bruwer, C.; Gough, C.; Bunce, M. Tropical ancient DNA from bulk archaeological fish bone reveals the subsistence practices of a historic coastal community in southwest madagascar. J. Archaeol. Sci. 2016, 75, 82–88. [Google Scholar] [CrossRef]
- Grier, C.; Flanigan, K.; Winters, M.; Jordan, L.G.; Lukowski, S.; Kemp, B.M. Using ancient DNA identification and osteometric measures of archaeological pacific salmon vertebrae for reconstructing salmon fisheries and site seasonality at dionisio point, british columbia. J. Archaeol. Sci. 2013, 40, 544–555. [Google Scholar] [CrossRef]
- Ólafsdóttir, G.Á.; Westfall, K.M.; Edvardsson, R.; Pálsson, S. Historical DNA reveals the demographic history of atlantic cod (gadus morhua) in medieval and early modern iceland. Proc. R. Soc. B 2014, 281. [Google Scholar] [CrossRef]
- Speller, C.F.; Hauser, L.; Lepofsky, D.; Moore, J.; Rodrigues, A.T.; Moss, M.L.; McKechnie, I.; Yang, D.Y. High potential for using DNA from ancient herring bones to inform modern fisheries management and conservation. PLoS ONE 2012, 7, e51122. [Google Scholar] [CrossRef] [PubMed]
- Splendiani, A.; Fioravanti, T.; Giovannotti, M.; Negri, A.; Ruggeri, P.; Olivieri, L.; Nisi Cerioni, P.; Lorenzoni, M.; Caputo Barucchi, V. The effects of paleoclimatic events on mediterranean trout: Preliminary evidences from ancient DNA. PLoS ONE 2016, 11, e0157975. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.; Arndt, U.; Lippold, S.; Benecke, N.; Debus, L.; King, T.; Matsumura, S. Tracing the first steps of american sturgeon pioneers in europe. BMC Evol. Biol. 2008, 8, 221. [Google Scholar] [CrossRef] [PubMed]
- Oleksiak, M.F. Genomic approaches with natural fish populations. J. Fish Biol. 2010, 76, 1067–1093. [Google Scholar] [CrossRef] [PubMed]
- Macqueen, D.J.; Primmer, C.R.; Houston, R.D.; Nowak, B.F.; Bernatchez, L.; Bergseth, S.; Davidson, W.S.; Gallardo-Escarate, C.; Goldammer, T.; Guiguen, Y.; et al. Functional annotation of all salmonid genomes (faasg): An international initiative supporting future salmonid research, conservation and aquaculture. BMC Genom. 2017, 18, 484. [Google Scholar] [CrossRef] [PubMed]
- Zohar, I.; Belmaker, M. Size does matter: Methodological comments on sieve size and species richness in fishbone assemblages. J. Archaeol. Sci. 2005, 32, 635–641. [Google Scholar] [CrossRef]
- Leuzinger, U. Die Jungsteinzeitliche Seeufersiedlung Arbon—Bleiche 3. Befunde. Archäologie im Thurgau. Band 9; Amt für Archäologie des Kantons Thurgau: Frauenfeld, Switzerland, 2000; Volume 9. [Google Scholar]
- Hagendorn, A. Zur Frühzeit von Vindonissa. Auswertung der Holzbauten der Grabung Windisch-Breite 1996–1998. Veröffentlichungen der Gesellschaft pro Vindonissa XVIII; Kantonsarchäologie Aargau: Brugg, Switzerland, 2003. [Google Scholar]
- Hüster Plogmann, H. Befunde und Fundkomplexe-der steingebaute Gutshof: Knochen aus Schlämmproben. In Der römische Gutshof in Neftenbach. Monographien der Kantonsarchäologie Zürich 31, Band 1; Kantonsarchäologie Zürich, Ed.; Fotorotar AG: Egg/Zürich, Switzerland, 1999; pp. 264–265. [Google Scholar]
- Rychener, J. Der Römische Gutshof in Neftenbach. Monographien der Kantonsarchäologie Zürich 31; Fotorotar AG: Egg/Zürich, Switzerland, 1999. [Google Scholar]
- Caduff, B.; Hüster Plogmann, H.; Diaz Tabernero, J.; Durst, M. Zum Frühmittelalterlichen Speisezettel in Tumegl/Tomils, Sogn Murez. In Jahresberichte des Archäologischen Dienstes Graubünden und der Denkmalpflege Graubünden 2002; Archäologischer Dienst Graubünden, Ed.; Archäologischer Dienst Graubünden: Haldenstein/Chur, Switzerland, 2003; pp. 96–115. [Google Scholar]
- Hüster Plogmann, H. Die frühmittelalterlichen Speisereste. In Zum frühmittelalterlichen Speisezettel in Tumegl/Tomils, Sogn Murezi. Jahresberichte des Archäologischen Dienstes Graubünden und der Denkmalpflege Graubünden 2002; Archäologischer Dienst Graubünden, Ed.; Archäologischer Dienst Graubünden: Haldenstein/Chur, Switzerland, 2003; pp. 100–113. [Google Scholar]
- Jecklin-Tischhauser, U. Die Kirchenanlage Sogn Murezi in Tomils (GR). Kirchliches und herrschaftliches Zentrum im frühmittelalterlichen Churrätien. Ph.D. Thesis, University of Zürich, Zürich, Switzerland, July 2017. [Google Scholar]
- Ohnsorg, P. Zwischen Limmat und Fraumünster. Neue Untersuchungen zur Uferzone am Zürcher Stadthausquai und zur Fraumünster-Abtei. In Stadt Zürich Archäologie und Denkmalpflege 2008–2010; Stadt Zürich: Zürich, Switzerland, 2011; pp. 1–65. [Google Scholar]
- Brombacher, C.; Helmig, G.; Hüster Plogmann, H.; Klee, M.; Rentzel, P.; Rodel, S.; Veszeli, M. Und was davon übrig bleibt—Untersuchungen an einem mittelalterlichen Latrinenschacht an der Bäumleingasse 14 (1992/20). In Archäologische Bodenforschung des Kantons Basel-Stadt. Jahresbericht 1998; Archäologische Bodenforschung des Kantons Basel-Stadt, Ed.; Archäologische Bodenforschung des Kantons Basel-Stadt: Basel, Switzerland, 1999; pp. 93–132. [Google Scholar]
- Häberle, S.; Schibler, J.; Van Neer, W.; Hüster Plogmann, H. Fischknochen als Indikatoren für Gewässerzustand und menschliche Fischselektion. Eine zusammenfassende Auswertung mittelalterlicher und neuzeitlicher Fischreste aus dem Rheineinzugsgebiet der Schweiz. Archäol. Korresp. 2015, 45, 417–437. [Google Scholar]
- Homberger, V. Ein neu entdecktes spätrömisches Kastell bei Weesen SG. Jahrb. Archäol. Schweiz 2008, 91, 141–149. [Google Scholar]
- Straumann, S. 2008/3, Münsterplatz 19, Museum der Kulturen: Ausgrabungen und Funde im Jahr 2009: Münsterhügel. In Archäologische Bodenforschung des Kantons Basel-Stadt. Jahresbericht 2009; Archäologische Bodenforschung des Kantons Basel-Stadt, Ed.; Archäologische Bodenforschung des Kantons Basel-Stadt: Basel, Switzerland, 2010; pp. 32–34. [Google Scholar]
- Jacomet, S.; Ebersbach, R.; Akeret, Ö.; Antolín, F.; Baum, T.; Bogaard, A.; Brombacher, C.; Bleicher, N.K.; Heitz-Weniger, A.; Hüster-Plogmann, H.; et al. On-site data cast doubts on the hypothesis of shifting cultivation in the late neolithic (c. 4300–2400 cal. BC): Landscape management as an alternative paradigm. Holocene 2016, 26, 1858–1874. [Google Scholar] [CrossRef]
- Menotti, F. Wetland Archaeology and Beyond: Theory and Practice; Oxford Universtity Press: Oxford, UK, 2012. [Google Scholar]
- Schibler, J. Zooarchaeological results from neolithic and bronze age wetland and dryland sites in the central alpine foreland: Economic, ecologic and taphonomic relevance. In Oxford Handbook of Archaeozoology; Albarella, U., Rizzetto, M., Russ, H., Vickers, K., Viner-Daniels, S., Eds.; Oxford University Press: Oxford, UK, 2017; pp. 83–98. [Google Scholar]
- Elsner, J.; Hofreiter, M.; Schibler, J.; Schlumbaum, A. Ancient mtdna diversity reveals specific population development of wild horses in switzerland after the last glacial maximum. PLoS ONE 2017, 12, e0177458. [Google Scholar] [CrossRef] [PubMed]
- Schibler, J.; Elsner, J.; Schlumbaum, A. Incorporation of aurochs into a cattle herd in neolithic europe: Single event or breeding? Sci. Rep. 2014, 4, 5798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binladen, J.; Gilbert, M.T.; Campos, P.F.; Willerslev, E. 5′-tailed sequencing primers improve sequencing quality of pcr products. BioTechniques 2007, 42, 174, 176. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.-J.; Macaulay, V.; Richards, M. Median networks: Speedy construction and greedy reduction, one simulation, and two case studies from human mtdna. Mol. Phylogenet. Evol. 2000, 16, 8–28. [Google Scholar] [CrossRef] [PubMed]
- Popović, D.; Panagiotopoulou, H.; Baca, M.; Stefaniak, K.; Mackiewicz, P.; Makowiecki, D.; King, T.L.; Gruchota, J.; Weglenski, P.; Stankovic, A. The history of sturgeon in the baltic sea. J. Biogeogr. 2014, 41, 1590–1602. [Google Scholar] [CrossRef]
- Schlumbaum, A.; Edwards, C.J. Ancient DNA reasearch on wetland archaeological evidence. In The Oxford Handbook of Wetland Archaeology; Menotti, F., O′Sullivan, A., Eds.; Oxford University Press: Oxford, UK, 2013; pp. 569–583. [Google Scholar]
- Elsner, J.; Schibler, J.; Hofreiter, M.; Schlumbaum, A. Burial condition is the most important factor for mtdna pcr amplification success in palaeolithic equid remains from the alpine foreland. Archaeol. Anthropol. Sci. 2015, 7, 505–515. [Google Scholar] [CrossRef]
- Pruvost, M.; Schwarz, R.; Correia, V.B.; Champlot, S.; Braguier, S.; Morel, N.; Fernandez-Jalvo, Y.; Grange, T.; Geigl, E.M. Freshly excavated fossil bones are best for amplification of ancient DNA. Proc. Natl. Acad. Sci. USA 2007, 104, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Giguet-Covex, C.; Pansu, J.; Arnaud, F.; Rey, P.J.; Griggo, C.; Gielly, L.; Domaizon, I.; Coissac, E.; David, F.; Choler, P.; et al. Long livestock farming history and human landscape shaping revealed by lake sediment DNA. Nat. Commun. 2014, 5, 3211. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Archaeological Site | City/Canton | Dating | Code Figure 1 | Elements | Site Type | Depositional Context | Reference |
|---|---|---|---|---|---|---|---|
| Stansstad-Kehrsiten | Kehrsiten/NW | 3500–3400 BC | 1 | 4 scales 4 vertebrae | Lake-shore settlement | Layer samples | [19] |
| Arbon Bleiche 3 | Arbon/TG | 3384–3370 BC | 2 | 4 vertebrae 4 scales 1 4 vertebrae 1 | Lake-shore settlement | Cultural layers | [36] |
| Breite | Windisch/AG | 1st century AD | 3 | 2 scales 2 vertebrae | dryland | Barrel pits, pits | [21,37] |
| Römerblick | Windisch/AG | 1st century AD | 4 | 1 vertebra | dryland | Kitchen floor of a peristyle house | [21,37] |
| Neftenbach | Neftenbach/ZH | 3th/4th century AD | 5 | 3 vertebrae | dryland | Cesspits | [38,39] |
| Tomils | Tomils/GR | 7th century AD | 6 | 6 vertebrae | dryland | Floor insulation structure | [40,41,42] |
| Fraumünsterstrasse | Zürich/ZH | 1010–1160 AD | 7 | 3 vertebrae | dryland | Occupation layer, filling of a fireplace | [43] |
| Bäumleingasse 14 | Basel/BS | 13th century AD | 8 | 2 vertebrae | dryland | Cesspits | [44] |
| Weesen Rosengärten | Weesen/SG | 14th century AD | 10 | 6 vertebrae | dryland | Waste trench | [45,46] |
| Museum der Kulturen, Im Schürhof | Basel/BS | 15th/16th century AD | 9 | 3 vertebrae | dryland | Cesspits | [45,47] |
| Primer Pairs | Coordinates Reference Sequence NC002646 | Nucleotide Sequence (5′–3′) | Annealing Temperature (°C) | Primer Length (bps) | Amplicon Length (bps) | Target Region Contains |
|---|---|---|---|---|---|---|
| CORb1F | 15,794–15,818 | TCAACATAAGTGATTTTAAGCCCTC | 54–55 | 25 | 90–91 | SNP1 |
| CORb1R | 15,864–15,883 | AGAACGGTTCCGTTGGTGAT | 20 | |||
| CORb1F | 15,794–15,818 | TCAACATAAGTGATTTTAAGCCCTC | 54 | 25 | 134–135 | SNP1 and SNP2 |
| COR1R | 15,908–15,927 | GCCCGTGTTAGTTGGAGGTT | 20 | |||
| CORc1F | 15,853–15,874 | AGACTCGGATAATCACCAACGG | 54–55 | 22 | 80 | SNP2 |
| CORc1R | 15,914–15,932 | ACGGAGCCCGTGTTAGTTG | 19 | |||
| CORa2F | 16,447–16,466 | TGTCAAACCCCAAAACCAGG | 54–55 | 20 | 78 | SNP3 |
| CORa2R | 16,502–16,524 | TGTCGGTGCCAAAGTTTGTTAAT | 23 | |||
| COR3F | 16,690–16,709 | TTGGCACCGACAACCCTATC | 54–55 | 20 | 86 | SNP4 |
| COR3R | 15–38 | ACAGCTTCAGTGTTATGCTTTAGT | 24 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso, J.D.G.; Häberle, S.; Plogmann, H.H.; Schibler, J.; Schlumbaum, A. Millennia-Long Co-Existence of Two Major European Whitefish (Coregonus spp.) Lineages in Switzerland Inferred from Ancient Mitochondrial DNA. Diversity 2017, 9, 34. https://0-doi-org.brum.beds.ac.uk/10.3390/d9030034
Alonso JDG, Häberle S, Plogmann HH, Schibler J, Schlumbaum A. Millennia-Long Co-Existence of Two Major European Whitefish (Coregonus spp.) Lineages in Switzerland Inferred from Ancient Mitochondrial DNA. Diversity. 2017; 9(3):34. https://0-doi-org.brum.beds.ac.uk/10.3390/d9030034
Chicago/Turabian StyleAlonso, José David Granado, Simone Häberle, Heidemarie Hüster Plogmann, Jörg Schibler, and Angela Schlumbaum. 2017. "Millennia-Long Co-Existence of Two Major European Whitefish (Coregonus spp.) Lineages in Switzerland Inferred from Ancient Mitochondrial DNA" Diversity 9, no. 3: 34. https://0-doi-org.brum.beds.ac.uk/10.3390/d9030034