Estimating Crop Nutritional Status Using Smart Apps to Support Nitrogen Fertilization. A Case Study on Paddy Rice

 ,
,
Abstract
:1. Introduction
- Evaluating the system’s capability to estimate N nutritional status and to support N management through dedicated field experiments;
- Proposing an effective procedure to derive calibration curves for indirect methods for PNC estimates, and demonstrating its suitability in a case study with PocketN and 43 rice cultivars widely grown in Europe. This procedure would allow extending the diagnostic system to production districts where other varieties are grown.
2. Materials and Methods
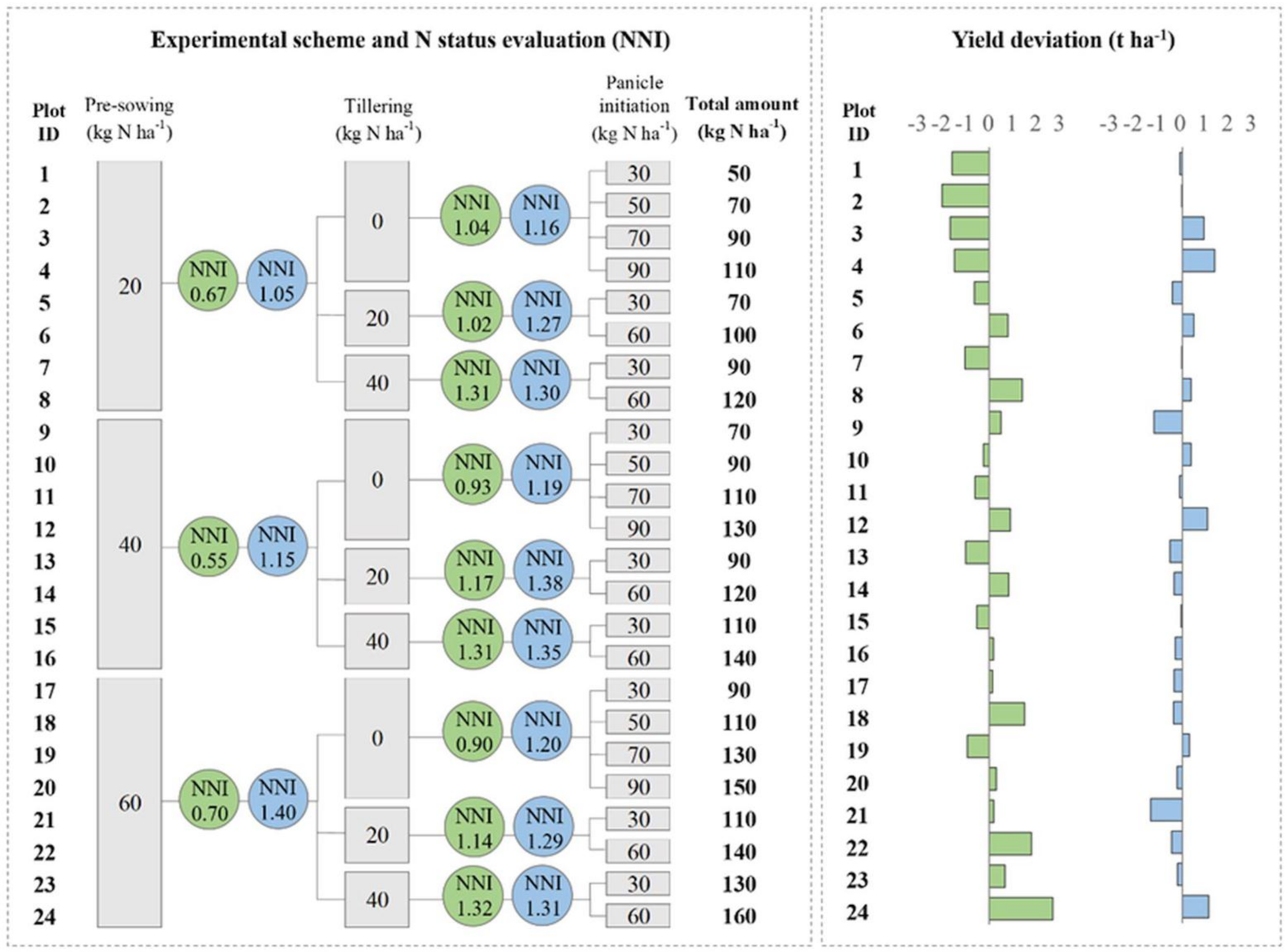
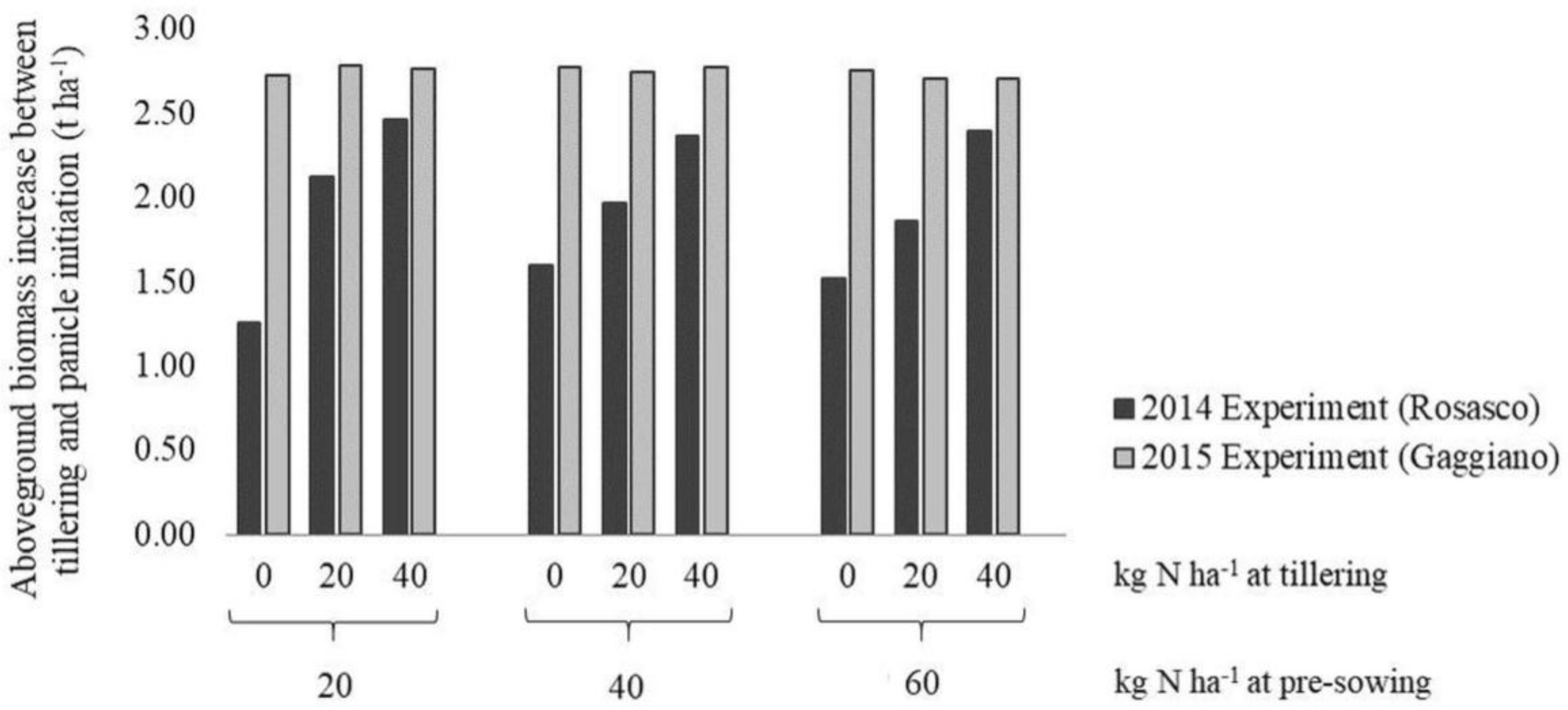
2.1. Experimental Design and Field Measurements to Develop and Test the Smart App-Based Diagnostic System for Supporting N Fertilization
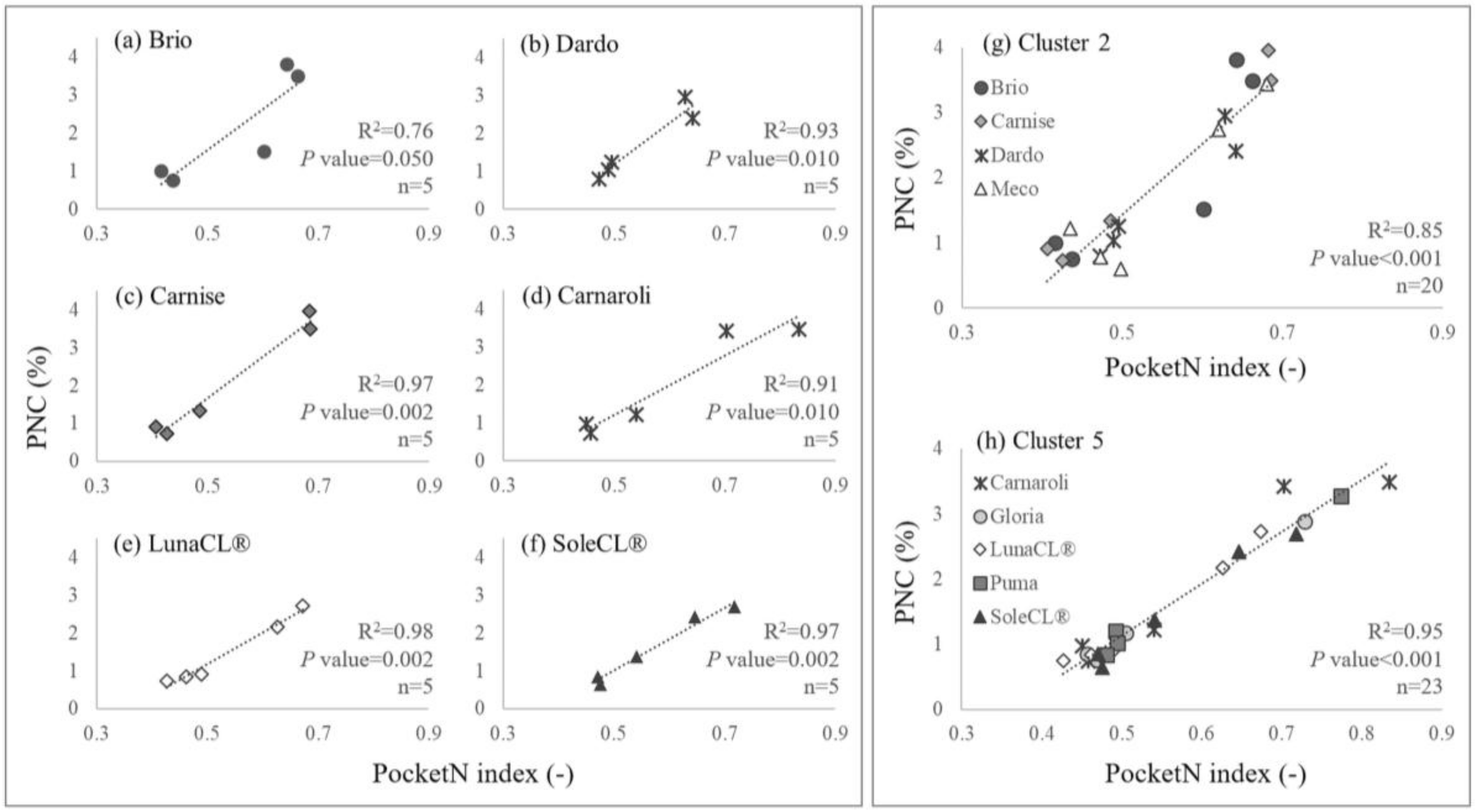
2.2. Definition of Calibration Curves for PocketN
3. Results
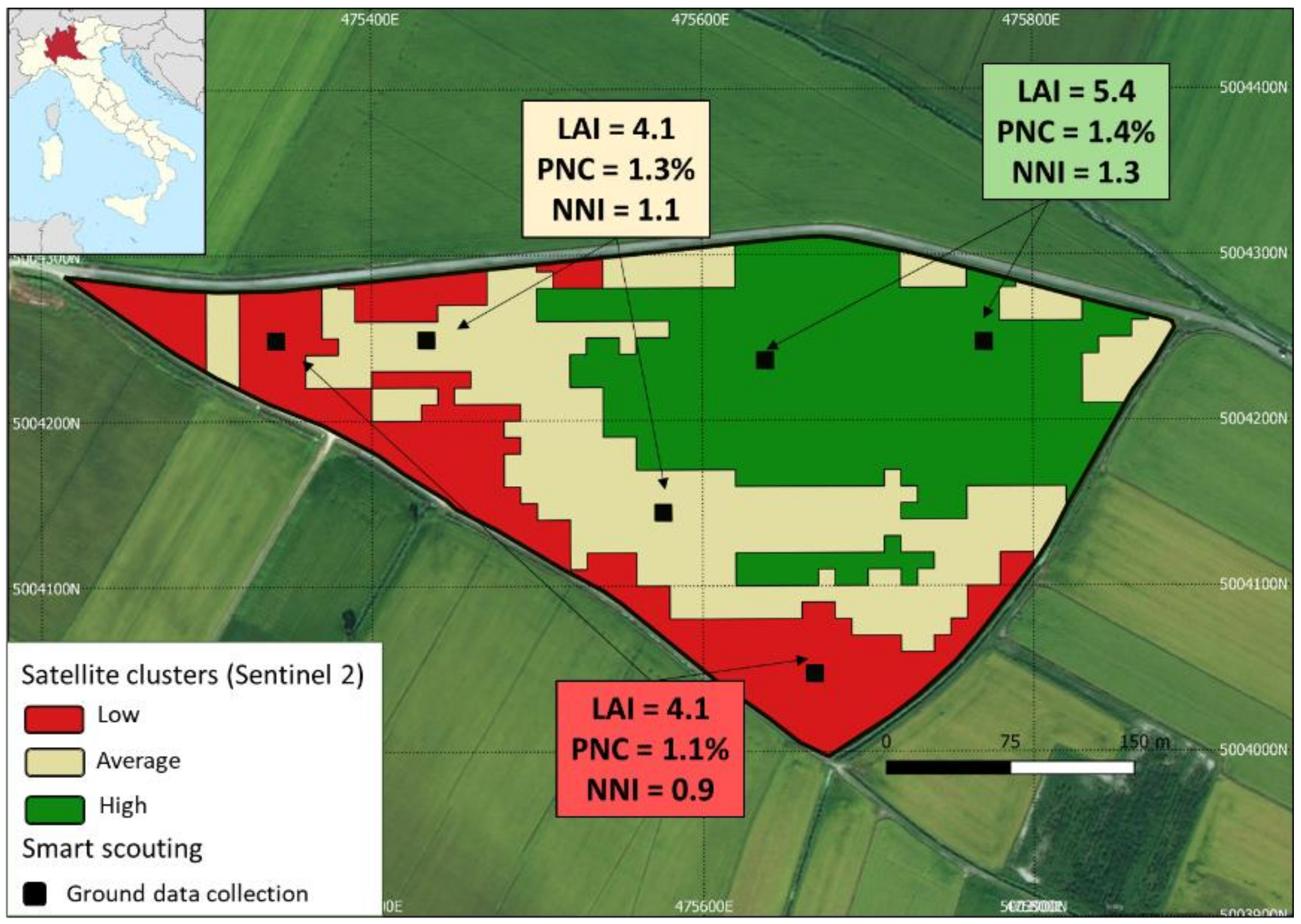
3.1. Evaluation of N Nutritional Status Via Smart Apps
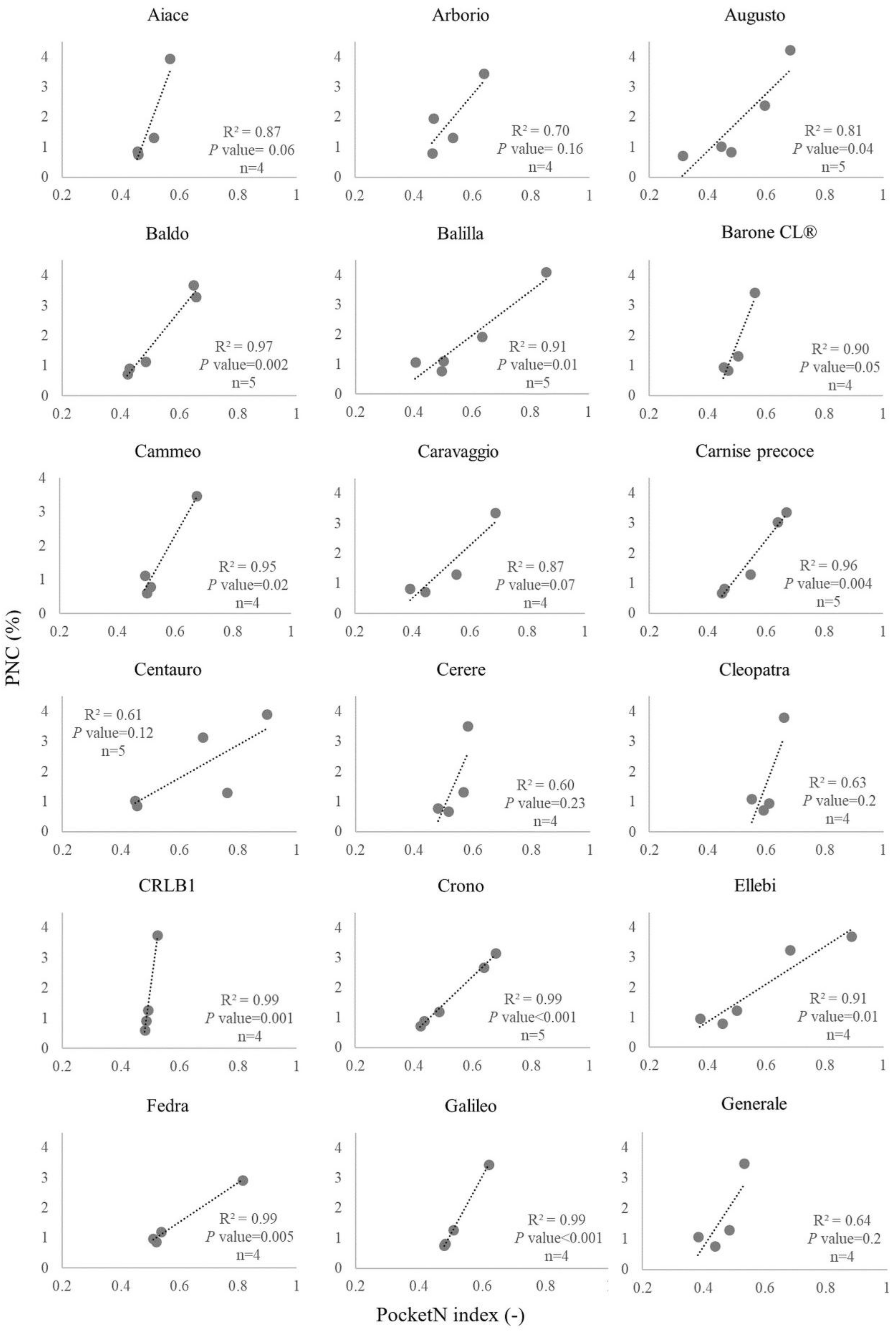
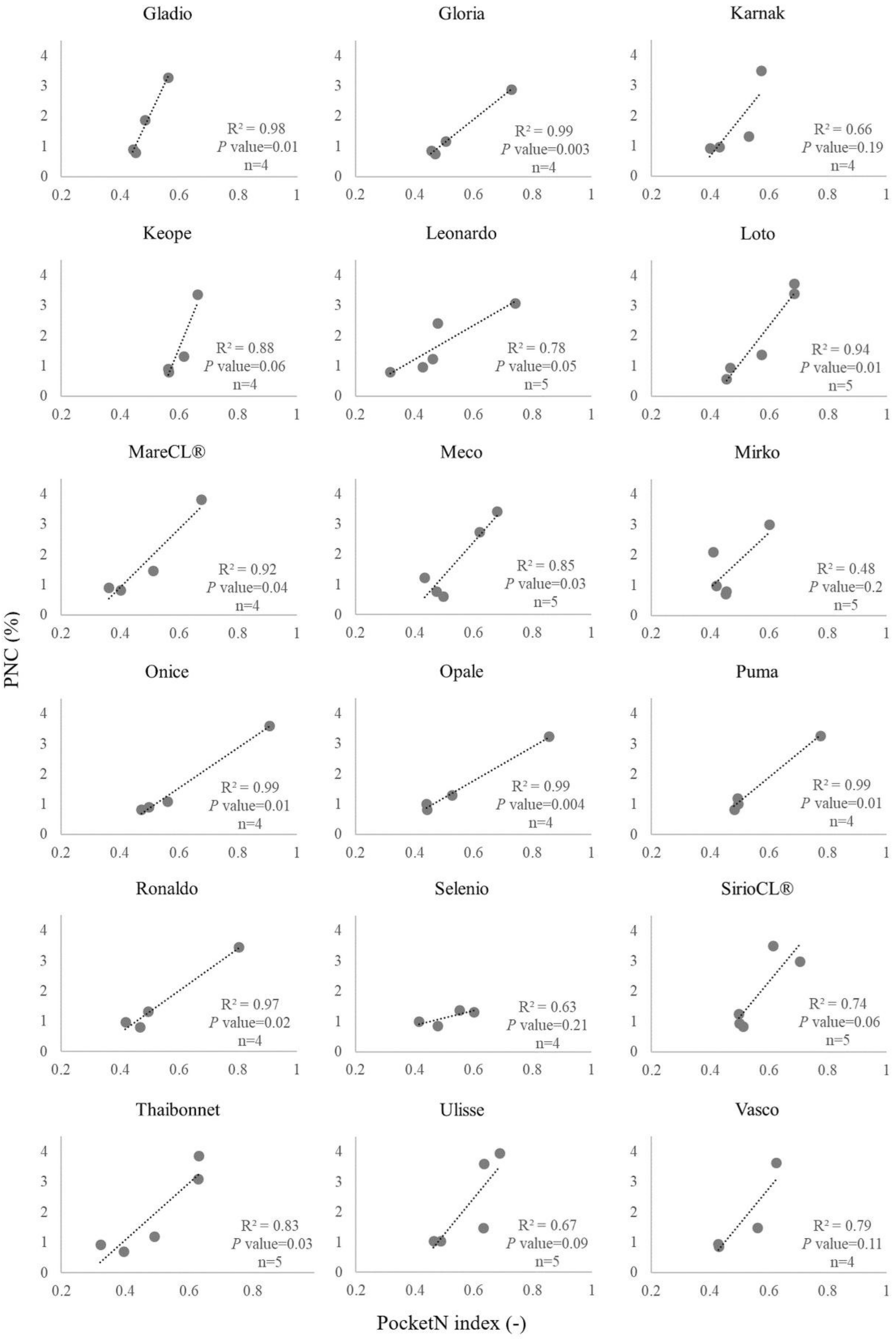
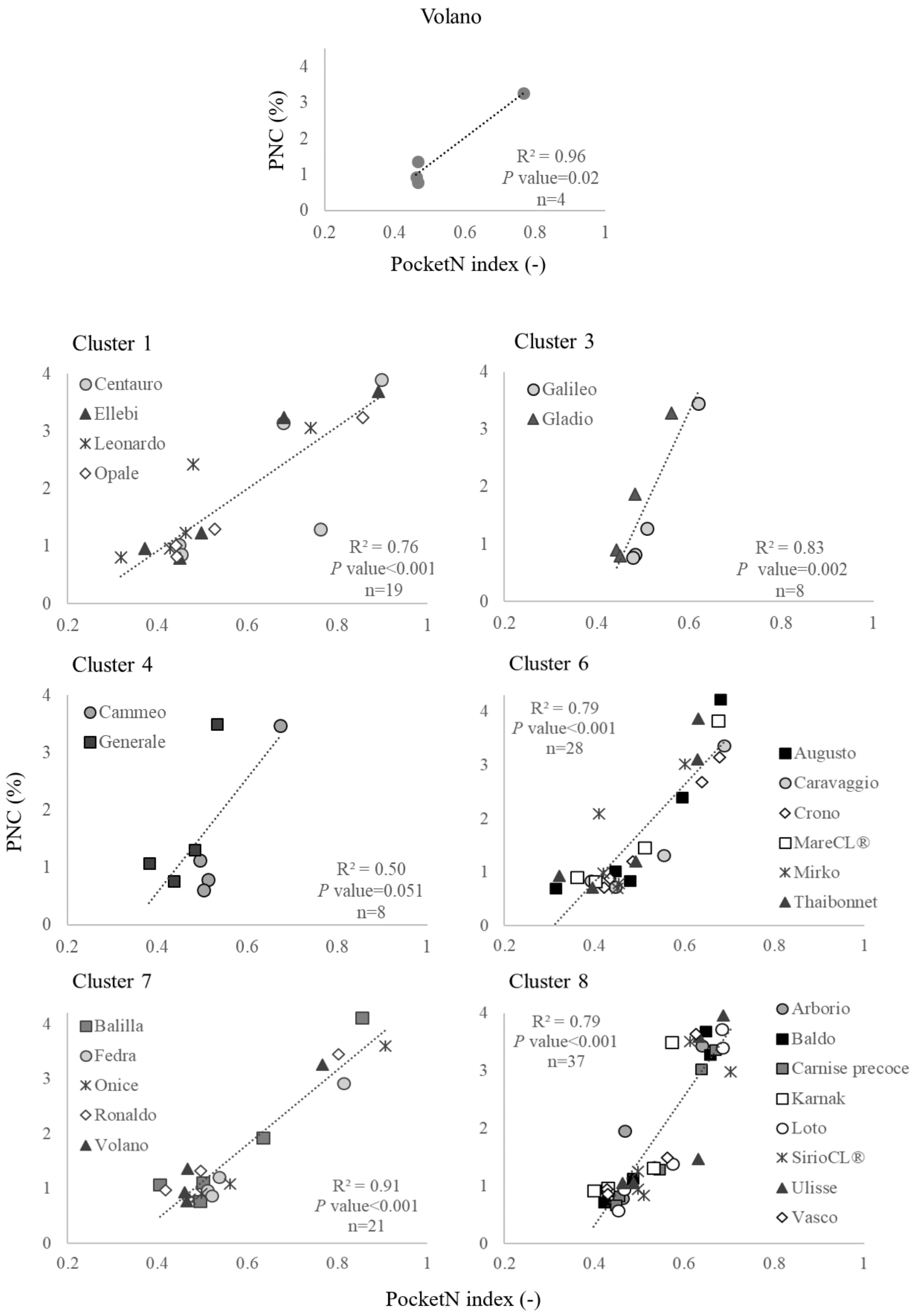
3.2. Calibration Curves for PocketN
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, N.D.; Gerber, J.S.; Johnston, M.; Deepak, K.R.; Ramankutty, N.; Foley, J.A. Closing yield gaps through nutrient and water management. Nature 2012, 490, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Goulding, K.; Jarvis, S.; Whitmore, A. Optimizing nutrient management for farm systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 667–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komarek, A.M.; Drogue, S.; Chenoune, R.; Hawkins, J.; Msangi, S.; Belhouchette, H.; Flichman, G. Agricultural household effects of fertilizer price changes for smallholder farmers in central Malawi. Agric. Syst. 2017, 154, 168–178. [Google Scholar] [CrossRef]
- Huang, H.; Nguyen Thi Thu, T.; He, X.; Gravot, A.; Bernillon, S.; Ballini, E.; Morel, J.-B. Increase of fungal pathogenicity and role of plant glutamine in nitrogen-induced susceptibility (NIS) to rice blast. Front. Plant Sci. 2017, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wu, L.; Ding, Y.; Yao, X.; Wu, X.; Weng, F.; Li, G.; Liu, Z.; Tang, S.; Ding, C.; et al. Nitrogen fertilizer application affects lodging resistance by altering secondary cell wall synthesis in japonica rice (Oryza sativa). J. Plant Res. 2017, 130, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.; Buresh, R.J.; Wang, Z.; Zhang, H.; Liu, L.; Yang, J.; Zhang, J. Roots and shoots traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates applications. Field Crop. Res. 2015, 175, 47–55. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Struik, P. Diverse concepts of breeding for nitrogen use efficiency. A review. Agron. Sustain. Dev. 2017, 37, 50. [Google Scholar] [CrossRef] [Green Version]
- Stafford, J.V.; Lark, R.M.; Bolam, H.C. Using Yield Maps to Regionalize Fields into Potential Management Units. In Precision Agriculture; Robert, P.C., Rush, R.H., Larson, W.E., Eds.; ASA/CSSA/SSSA: Madison, WI, USA, 1999; pp. 225–237. [Google Scholar]
- Koch, B.; Khosla, R.; Frasier, W.M.; Westfall, D.G.; Inman, D. Site specific management: Economic feasibility of variable-rate nitrogen application utilizing site-specific management zones. Agron. J. 2004, 96, 1572–1580. [Google Scholar] [CrossRef]
- Raun, W.R.; Solie, J.B.; Stone, M.L.; Martin, K.L.; Freeman, K.W.; Mullen, R.W.; Zhang, H.; Schepers, J.S.; Johnson, G.V. Optical sensor-based algorithm for crop nitrogen fertilization. Commun. Soil Sci. Plan. 2005, 36, 2759–2781. [Google Scholar] [CrossRef]
- Basso, B.; Dumont, B.; Cammarano, D.; Pezzuolo, A.; Marinello, F.; Sartori, L. Environmental and economic benefits of variable rate nitrogen fertilization in a nitrate vulnerable zone. Sci. Total Environ. 2016, 545–546, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Casa, R.; Pelosi, F.; Pascucci, S.; Fontana, F.; Castaldi, F.; Pignatti, S.; Pepe, M. Early stage variable rate nitrogen fertilization of silage maize driven by multi-temporal clustering of archive satellite data. Adv. Anim. Biosci. 2017, 8, 288–292. [Google Scholar] [CrossRef]
- Schwalbert, R.A.; Amado, T.J.C.; Reimche, G.B.; Gebert, F. Fine-tuning of wheat (Triticum aestivum, L.) variable nitrogen rate by combining crop sensing and management zones approaches in southern Brazil. Precis. Agric. 2019, 20, 56–77. [Google Scholar] [CrossRef]
- Salette, J.; Lemaire, G. Sur la Variation de la Teneur en Azote des Graminées Fourragères Pendant Leur Croissance: Formulation D’une Loi de Diluition, Série III. ed.; Compte Rendus de l’académie des Sciences: Paris, France, 1981; Volume 292, pp. 875–878. [Google Scholar]
- Ata-Ul-Karim, S.T.; Zhu, Y.; Liu, X.; Cao, Q.; Tian, Y.; Cao, W. Comparison of different critical nitrogen curves for nitrogen diagnosis in rice. Sci. Rep. 2017, 7, 42679. [Google Scholar] [CrossRef] [PubMed]
- Justes, E.; Mary, B.; Meynard, J.M.; Machet, J.M.; Thelier-huche, L. Determination of a critical nitrogen dilution curve for winter wheat crops. Ann. Bot. 1994, 74, 397–407. [Google Scholar] [CrossRef]
- Lemaire, G.; Jeuffroy, M.H.; Gastal, F. Diagnosis tool for plant and crop N status in vegetative stage. Theory and practices for crop N management. Eur. J. Agron. 2008, 28, 614–624. [Google Scholar] [CrossRef]
- Chen, P. A comparison of two approaches for estimating the wheat nitrogen nutrition index using remote sensing. Remote Sens. 2015, 7, 4527–4548. [Google Scholar] [CrossRef]
- Huang, S.; Miao, Y.; Zhao, G.; Yuan, F.; Ma, X.; Tan, C.; Yu, W.; Gnyp, M.; Lenz-Wiedemann, V.; Rascher, U.; et al. Satellite remote sensing-based in-season diagnosis of rice nitrogen status in Northeast China. Remote Sens. 2015, 7, 10646–10667. [Google Scholar] [CrossRef]
- Morisette, J.T.; Baret, F.; Privette, J.L.; Myneni, R.B.; Nickeson, J.E.; Garrigues, S.; Shabanov, N.V.; Weiss, M.; Fernandes, R.A.; Leblanc, S.G.; et al. Validation of global moderate-resolution LAI products: A framework proposed within the CEOS land product validation subgroup. IEEE Trans. Geosci. Remote 2006, 44, 1804–1814. [Google Scholar] [CrossRef]
- Muñoz-Huerta, R.F.; Guevara-Gonzalez, R.G.; Contreras-Medina, L.M.; Torres-Pacheco, I.; Prado-Olivarez, J.; Ocampo-Velazquez, R.V. A review of methods for sensing the nitrogen status in plants: Advantages, disadvantages and recent advances. Sensors 2013, 13, 10823–10843. [Google Scholar] [CrossRef] [PubMed]
- Orlando, F.; Movedi, E.; Coduto, D.; Parisi, S.; Brancadoro, L.; Pagani, V.; Guarneri, T.; Confalonieri, R. Estimating LAI in vineyard using the PocketLAI smart-app. Sensors 2016, 16, 2004. [Google Scholar] [CrossRef] [PubMed]
- Nutini, F.; Confalonieri, R.; Crema, A.; Movedi, E.; Paleari, L.; Stavrakoudis, D.; Boschetti, M. An operational workflow to assess rice nutritional status based on satellite imagery and smartphone apps. Comput. Electron. Agric. 2018, 154, 80–92. [Google Scholar] [CrossRef]
- Alam, M.M.; Ladha, J.K.; Khan, S.R.; Foyjunessa; Harun-ur-Rashid; Khan, A.H.; Buresh, R.J. Leaf color chart for managing nitrogen fertilizer in lowland rice in Bangladesh. Agron. J. 2005, 97, 949–959. [Google Scholar] [CrossRef]
- Confalonieri, R.; Debellini, C.; Pirondini, M.; Possenti, P.; Bergamini, L.; Barlassina, G.; Bartoli, A.; Agostoni, E.G.; Appiani, M.; Babazadeh, L.; et al. A new approach for determining rice critical nitrogen concentration. J. Agric. Sci. 2011, 149, 633–638. [Google Scholar] [CrossRef]
- Peng, S.; Garcia, F.V.; Laza, R.C.; Sanico, A.L.; Visperas, R.M.; Cassman, K.G. Increased N-use efficiency using a chlorophyll meter on high-yielding irrigated rice. Field Crop. Res. 1996, 47, 243–252. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M.; Baret, F. Review of methods for in situ leaf area index determination. Part I. Theories, sensors and hemispherical photography. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Stroppiana, D.; Boschetti, M.; Confalonieri, R.; Bocchi, S.; Brivio, P.A. Evaluation of LAI-2000 for leaf area index monitoring in paddy rice. Field Crop. Res. 2006, 99, 167–170. [Google Scholar] [CrossRef]
- Cerovic, Z.G.; Masdoumier, G.; Ghozlen, N.B.; Latouche, G. A new optical leaf-clip meter for simultaneous nondestructive assessment of leaf chlorophyll and epidermal flavonoids. Physiol. Plant. 2012, 146, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Giunta, F.; Motzo, R.; Deidda, M. SPAD readings and associated leaf traits in durum wheat, barley and triticale cultivars. Euphytica 2002, 125, 197–205. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, Y.; Qin, Y.; Chen, Y.; Fan, M. Establishing dynamic thresholds for potato nitrogen status diagnosis with the SPAD chlorophyll meter. J. Integr. Agr. 2015, 14, 190–195. [Google Scholar] [CrossRef]
- Ravier, C.; Quemada, M.; Jeuffroy, M.-H. Use of a chlorophyll meter to assess nitrogen nutrition index during the growth cycle in winter wheat. Field Crop. Res. 2016, 214, 73–82. [Google Scholar] [CrossRef]
- Ata-Ul-Karim, S.T.; Cao, Q.; Zhu, Y.; Tang, L.; Rehmani, M.I.A.; Cao, W. Non-destructive assessment of plant nitrogen parameters using leaf chlorophyll measurements in rice. Front. Plant Sci. 2016, 7, 1829. [Google Scholar] [CrossRef] [PubMed]
- Confalonieri, R.; Foi, M.; Casa, R.; Aquaro, S.; Tona, E.; Peterle, M.; Boldini, A.; De Carli, G.; Ferrari, A.; Finotto, G.; et al. Development of an app for estimating leaf area index using a smartphone. Trueness and precision determination and comparison with other indirect methods. Comput. Electron. Agric. 2013, 96, 67–74. [Google Scholar] [CrossRef]
- Confalonieri, R.; Paleari, L.; Movedi, E.; Pagani, V.; Orlando, F.; Foi, M.; Barbieri, M.; Pesenti, M.; Cairati, O.; La Sala, M.S.; et al. Improving in vivo plant nitrogen content estimates from digital images: Trueness and precision of a new approach as compared to other methods and commercial devices. Biosyst. Eng. 2015, 135, 21–30. [Google Scholar] [CrossRef]
- Baruth, B.; Van den Berg, M.; Niemeyer, S. MARS Bulletin, 2014, Volume 22, n.13. Available online: publications.jrc.ec.europa.eu/repository/bitstream/JRC88391/marsbulletin_vol22_no13.pdf (accessed on 23 February 2019).
- Baruth, B.; Van den Berg, M.; Niemeyer, S. MARS Bulletin, 2015, Volume 23, n.8. Available online: publications.jrc.ec.europa.eu/repository/bitstream/JRC98793/lb-am-15-008-en-n.pdf (accessed on 23 February 2019).
- Lee, J.-H.; Kang, C.-S.; Roh, A.-S.; Park, K.Y.; Lee, H.J. Assessment of N topdressing rate at panicle initiation stage with chlorophyll meter-based diagnosis in rice. J. Crop Sci. Biotechnol. 2009, 12, 195–200. [Google Scholar] [CrossRef]
- Karcher, D.E.; Richardson, M.D. Quantifying turfgrass color using digital image analysis. Crop Sci. 2003, 43, 943–951. [Google Scholar] [CrossRef]
- Warren-Wilson, J. Estimation of foliage denseness and foliage angle by inclined point quadrats. Aust. J. Bot. 1963, 11, 95–105. [Google Scholar] [CrossRef]
- Baret, F.; de Solan, B.; Lopez-Lozano, R.; Ma, K.; Weiss, M. GAI estimates of row crops from downward looking digital photos taken perpendicular to rows at 57.5° zenith angle: Theoretical considerations based on 3D architecture models and application to wheat crops. Agric. For. Meteorol. 2010, 150, 1393–1401. [Google Scholar] [CrossRef]
- Gomez, K.A. Techniques for Field Experiments with Rice: Layout, Sampling, Sources of Error; International Rice Research Institute: Los Baños, Philippines, 1972; 46p. [Google Scholar]
- Confalonieri, R.; Stroppiana, D.; Boschetti, M.; Gusberti, D.; Bocchi, S.; Acutis, M. Analysis of rice sample size variability due to development stage, nitrogen fertilization, sowing technique and variety using the visual jackknife. Field Crop. Res. 2006, 97, 135–141. [Google Scholar] [CrossRef]
- Williams, J.R.; Jones, C.A.; Kiniry, J.R.; Spanel, D.A. The EPIC crop growth model. Trans. ASAE 1989, 32, 497–511. [Google Scholar] [CrossRef]
- Greenwood, D.J.; Lemaire, G.; Gosse, G.; Cruz, P.; Draycott, A.; Neeteson, J.J. Decline in percentage N of C3 and C4 crops with increasing plant mass. Ann. Bot. 1990, 66, 425–436. [Google Scholar] [CrossRef]
- MacQueen, J.B. Some Methods for Classification and Analysis of Multivariate Observations. In Proceedings of the 5th Berkeley Symposium on Mathematical Statistics and Probability, Berkeley, CA, USA; University of California Press: Berkeley, CA, USA, 1967; Volume 1, pp. 281–297. [Google Scholar]
- Zhao, B.; Liu, Z.; Ata-Ul-Karim, S.T.; Xiao, J.; Liu, Z.; Qi, A.; Ning, D.; Nan, J.; Duan, A. Rapid and nondestructive estimation of the nitrogen nutrition index in winter barley using chlorophyll measurements. Field Crop. Res. 2016, 185, 59–68. [Google Scholar] [CrossRef]
- Ziadi, N.; Brassard, M.; Bélanger, G.; Cambouris, A.N.; Tremblay, N.; Nolin, M.C.; Claessens, A.; Parent, L.E. Critical nitrogen curve and nitrogen nutrition index for corn in eastern Canada. Agron. J. 2008, 100, 271–276. [Google Scholar] [CrossRef]
- Ata-Ul-Karim, S.T.; Liu, X.; Lu, Z.; Yuan, Z.; Zhu, Y.; Cao, W. In-season estimation of rice grain yield using critical nitrogen dilution curve. Field Crop. Res. 2016, 195, 1–8. [Google Scholar] [CrossRef]
- Gaines, T.P.; Gaines, S.T. Soil texture effect on nitrate leaching in soil percolates. Commun. Soil Sci. Plant Anal. 2008, 25, 2561–2570. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Davis, J.G.; Malzer, G.L.; Copeland, P.J.; Lamb, J.A.; Robert, P.C.; Bruulsema, T.W. Using yield variability to characterize spatial crop response to applied N. In Proceedings of the 3rd International Conference Precision Agriculture, Minneapolis, MN, USA, 23–26 June 1996; American Society of Agronomy: Madison, WI, USA, 1996; pp. 513–519. [Google Scholar]
- Chen, Q.; Tian, Y.; Yao, X.; Cao, W.; Zhu, Y. Comparison of five different nitrogen dressing methods to optimize rice growth. Plant Prod. Sci. 2014, 17, 66–80. [Google Scholar] [CrossRef]
- Sheehy, J.E.; Mnzava, M.; Cassman, K.G.; Mitchell, P.L.; Pablico, P.; Robles, R.P.; Samonte, H.P.; Lales, J.S.; Ferrer, A.B. Temporal origin of nitrogen in the grain of irrigated rice in the dry season: The outcome or recycling, cycling, senescence and competition studied using a 15N-point placement technique. Field Crop. Res. 2004, 89, 337–348. [Google Scholar] [CrossRef]
- Markwell, J.; Osterman, J.C.; Mitchell, J.L. Calibration of the Minolta SPAD-502 leaf chlorophyll meter. Photosynth. Res. 1995, 46, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M.; Hunt, E.R., Jr.; Mutters, R.G. Assessment of leaf color chart observations for estimating maize chlorophyll content by analysis of digital photographs. Agron. J. 2014, 108, 822–829. [Google Scholar] [CrossRef]
- Hirasawa, T.; Ozawa, S.; Taylaran, R.; Ookawa, T. Varietal differences in photosynthetic rates in rice plants, with special reference to nitrogen content of leaves. Plant Prod. Sci. 2010, 13, 53–57. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Cultivar | Calibration Curve Parameters a | R2 | p-Value | ID | Cultivar | Calibration Curve Parameters a | R2 | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| a | b | a | b | ||||||||
| 1 | Aiace | 26.55 | −11.53 | 0.87 | 0.060 | 23 | Gladio | 21.00 | −8.47 | 0.98 | 0.010 |
| 2 | Arborio | 11.62 | −4.23 | 0.70 | 0.160 | 24 | Gloria | 7.78 | −2.78 | 0.99 | 0.003 |
| 3 | Augusto | 9.54 | −2.95 | 0.81 | 0.040 | 25 | Karnak | 12.17 | −4.21 | 0.66 | 0.190 |
| 4 | Baldo | 12.08 | −4.44 | 0.97 | 0.002 | 26 | Keope | 23.70 | −12.64 | 0.88 | 0.060 |
| 5 | Balilla | 7.45 | −2.51 | 0.91 | 0.010 | 27 | Leonardo | 5.63 | −1.04 | 0.78 | 0.050 |
| 6 | BaroneCL® | 24.67 | −10.62 | 0.90 | 0.050 | 28 | Loto | 12.25 | −4.98 | 0.94 | 0.010 |
| 7 | Brio | 10.70 | −3.79 | 0.76 | 0.050 | 29 | LunaCL® | 8.32 | −2.97 | 0.98 | 0.002 |
| 8 | Cammeo | 15.21 | −6.81 | 0.95 | 0.020 | 30 | MareCL® | 9.63 | −2.94 | 0.92 | 0.040 |
| 9 | Caravaggio | 8.75 | −2.99 | 0.87 | 0.070 | 31 | Meco | 11.15 | −4.28 | 0.85 | 0.030 |
| 10 | Carnaroli | 7.83 | −2.71 | 0.91 | 0.010 | 32 | Mirko | 9.00 | −2.69 | 0.48 | 0.200 |
| 11 | Carnise | 10.95 | −3.80 | 0.97 | 0.002 | 33 | Onice | 6.61 | −2.43 | 0.99 | 0.010 |
| 12 | Carnise precoce | 12.26 | −4.92 | 0.96 | 0.004 | 34 | Opale | 5.61 | −1.59 | 0.99 | 0.004 |
| 13 | Centauro | 5.50 | −1.52 | 0.61 | 0.120 | 35 | Puma | 7.93 | −2.86 | 0.99 | 0.010 |
| 14 | Cerere | 22.00 | −10.23 | 0.60 | 0.230 | 36 | Ronaldo | 6.94 | −2.15 | 0.97 | 0.020 |
| 15 | Cleopatra | 25.39 | −13.64 | 0.63 | 0.200 | 37 | Selenio | 2.40 | −0.09 | 0.63 | 0.210 |
| 16 | CRLB1 | 71.52 | −33.83 | 0.99 | 0.001 | 38 | SirioCL® | 11.71 | −4.70 | 0.74 | 0.060 |
| 17 | Crono | 9.32 | −3.23 | 0.99 | <0.001 | 39 | SoleCL® | 8.34 | −3.16 | 0.97 | 0.002 |
| 18 | Dardo | 10.94 | −4.28 | 0.93 | 0.010 | 40 | Thaibonnet | 9.41 | −2.69 | 0.83 | 0.030 |
| 19 | Ellebi | 6.29 | −1.65 | 0.91 | 0.010 | 41 | Ulisse | 11.91 | −4.69 | 0.67 | 0.090 |
| 20 | Fedra | 6.52 | −2.39 | 0.99 | 0.005 | 42 | Vasco | 11.74 | −4.28 | 0.79 | 0.110 |
| 21 | Galileo | 19.20 | −8.46 | 0.99 | <0.001 | 43 | Volano | 7.39 | −2.41 | 0.96 | 0.020 |
| 22 | Generale | 15.36 | −5.38 | 0.64 | 0.200 | ||||||
| Cluster | Cultivars a | Calibration Curve Parameters b | R2 | p-Value | |
|---|---|---|---|---|---|
| a | b | ||||
| 1 | Centauro, Ellebi, Leonardo, Opale | 5.42 | −1.24 | 0.76 | <0.001 |
| 2 | Brio, Carnise, Dardo, Meco | 10.90 | −4.02 | 0.85 | <0.001 |
| 3 | Galileo, Gladio | 17.22 | −7.03 | 0.83 | 0.002 |
| 4 | Cammeo, Generale | 9.97 | −3.43 | 0.50 | 0.051 |
| 5 | Carnaroli, Gloria, LunaCL®, Puma, SoleCL® | 7.99 | −2.87 | 0.95 | <0.001 |
| 6 | Augusto, Caravaggio, Crono, MareCL®, Mirko, Thaibonnet | 9.04 | −2.79 | 0.79 | <0.001 |
| 7 | Balilla, Fedra, Onice, Ronaldo, Volano | 6.77 | −2.25 | 0.91 | <0.001 |
| 8 | Arborio, Baldo, Carnise Precoce, Karnak, Loto, SirioCL®, Ulisse, Vasco | 11.25 | −4.19 | 0.79 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paleari, L.; Movedi, E.; Vesely, F.M.; Thoelke, W.; Tartarini, S.; Foi, M.; Boschetti, M.; Nutini, F.; Confalonieri, R. Estimating Crop Nutritional Status Using Smart Apps to Support Nitrogen Fertilization. A Case Study on Paddy Rice. Sensors 2019, 19, 981. https://0-doi-org.brum.beds.ac.uk/10.3390/s19040981
Paleari L, Movedi E, Vesely FM, Thoelke W, Tartarini S, Foi M, Boschetti M, Nutini F, Confalonieri R. Estimating Crop Nutritional Status Using Smart Apps to Support Nitrogen Fertilization. A Case Study on Paddy Rice. Sensors. 2019; 19(4):981. https://0-doi-org.brum.beds.ac.uk/10.3390/s19040981
Chicago/Turabian StylePaleari, Livia, Ermes Movedi, Fosco M. Vesely, William Thoelke, Sofia Tartarini, Marco Foi, Mirco Boschetti, Francesco Nutini, and Roberto Confalonieri. 2019. "Estimating Crop Nutritional Status Using Smart Apps to Support Nitrogen Fertilization. A Case Study on Paddy Rice" Sensors 19, no. 4: 981. https://0-doi-org.brum.beds.ac.uk/10.3390/s19040981