α-Glucosidase Inhibitory Activity of Cycloartane-Type Triterpenes Isolated from Indonesian Stingless Bee Propolis and Their Structure–Activity Relationship


Abstract
:1. Introduction
2. Results
2.1. Extracts and Fractions Obtained from T. sapiens Propolis
2.2. Total Phenolic Content and Antioxidant Activity of T. sapiens Propolis
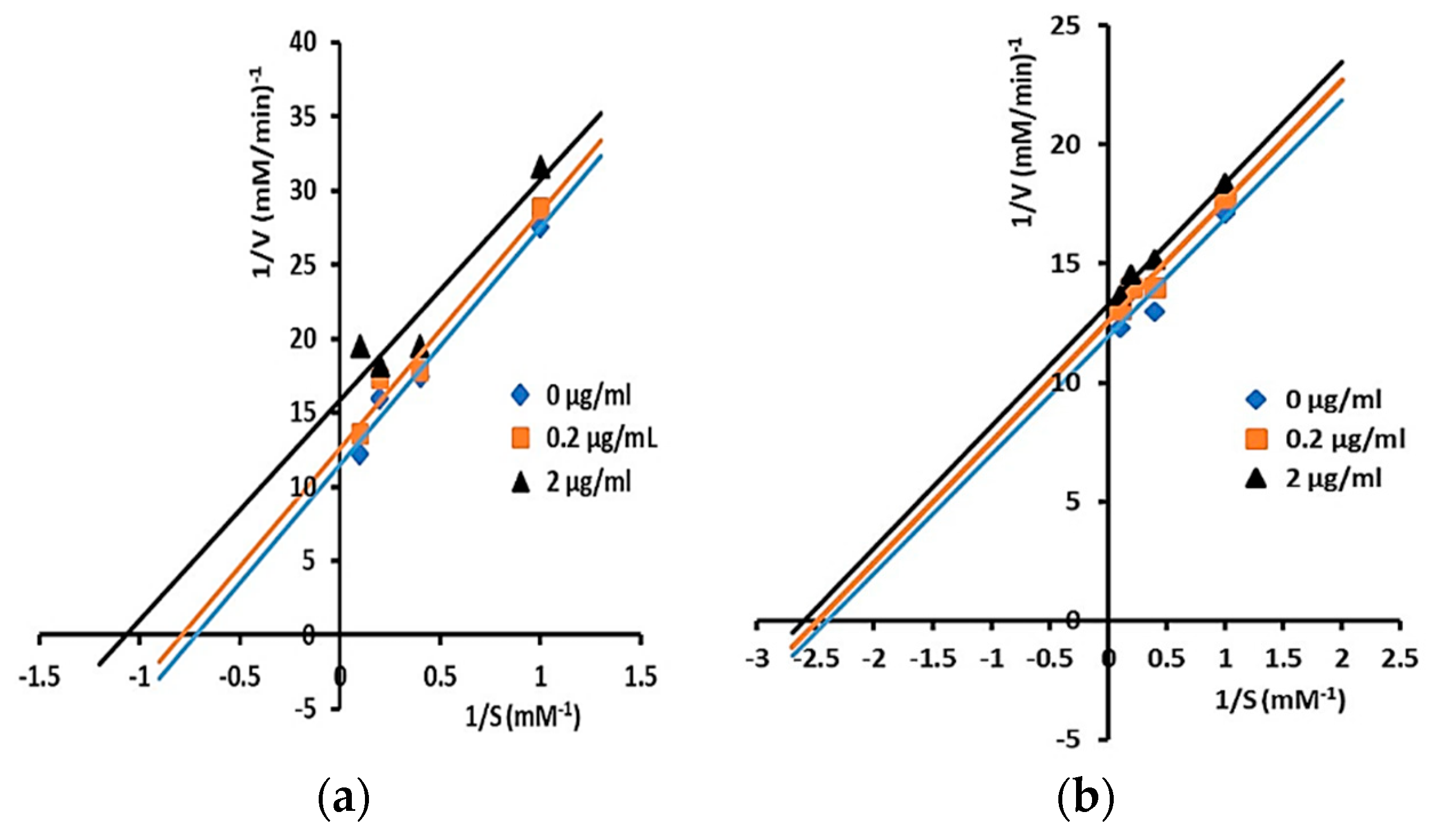
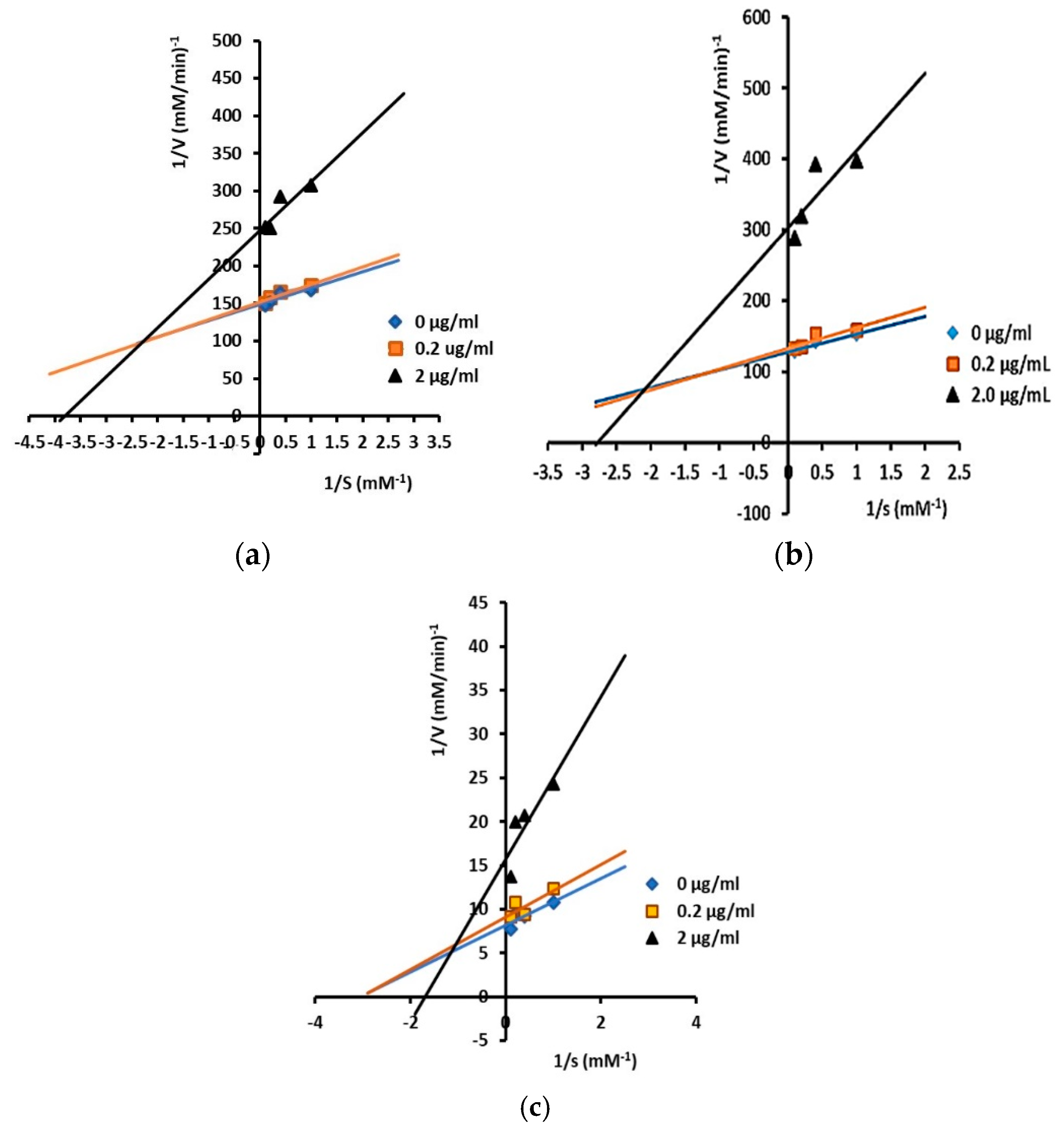
2.3. Inhibitory Activity and Kinetic Inhibition of Cycloartane-Type Triterpenes Isolated from T. sapiens Bee Propolis on α-Glucosidases
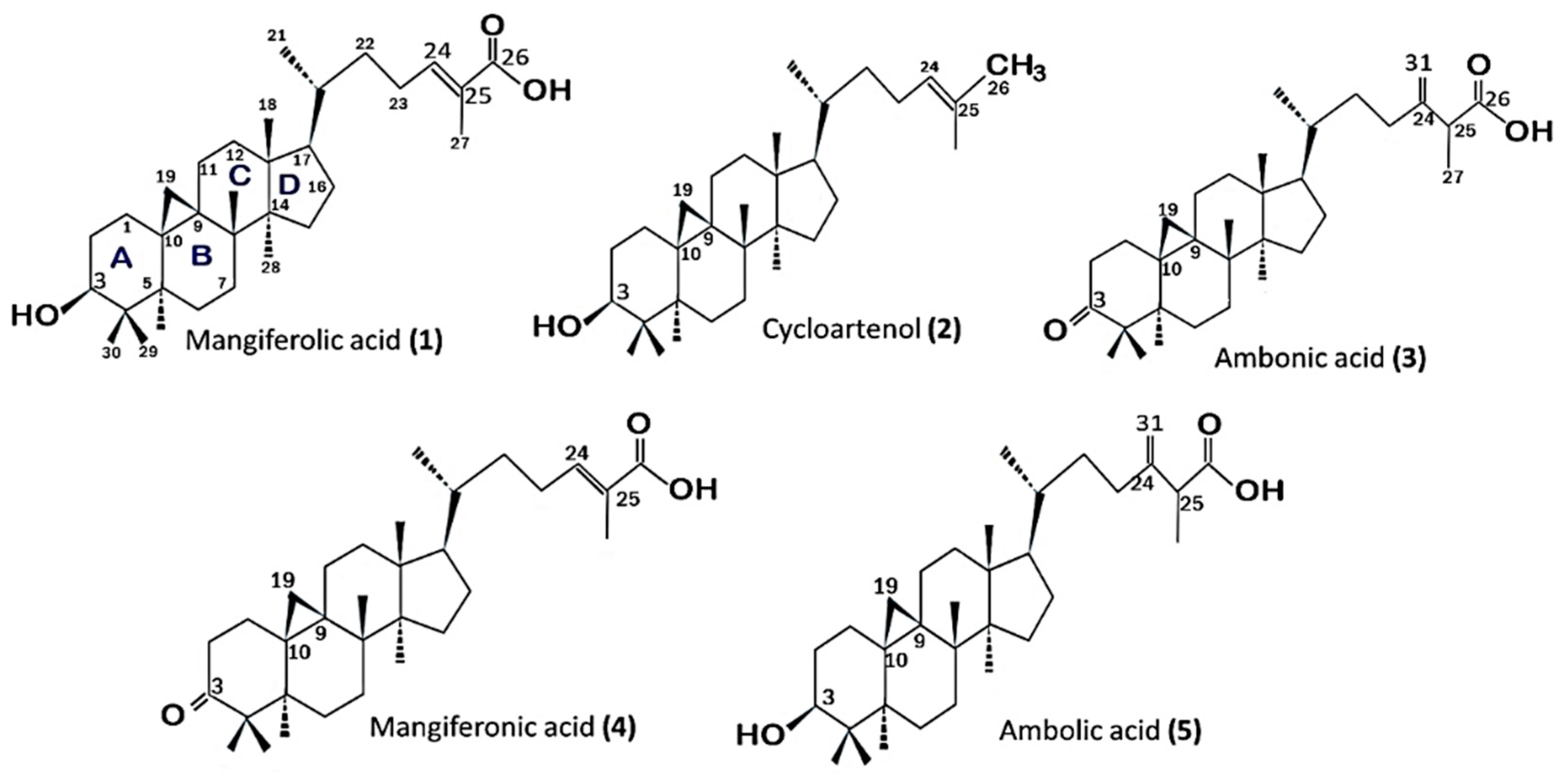
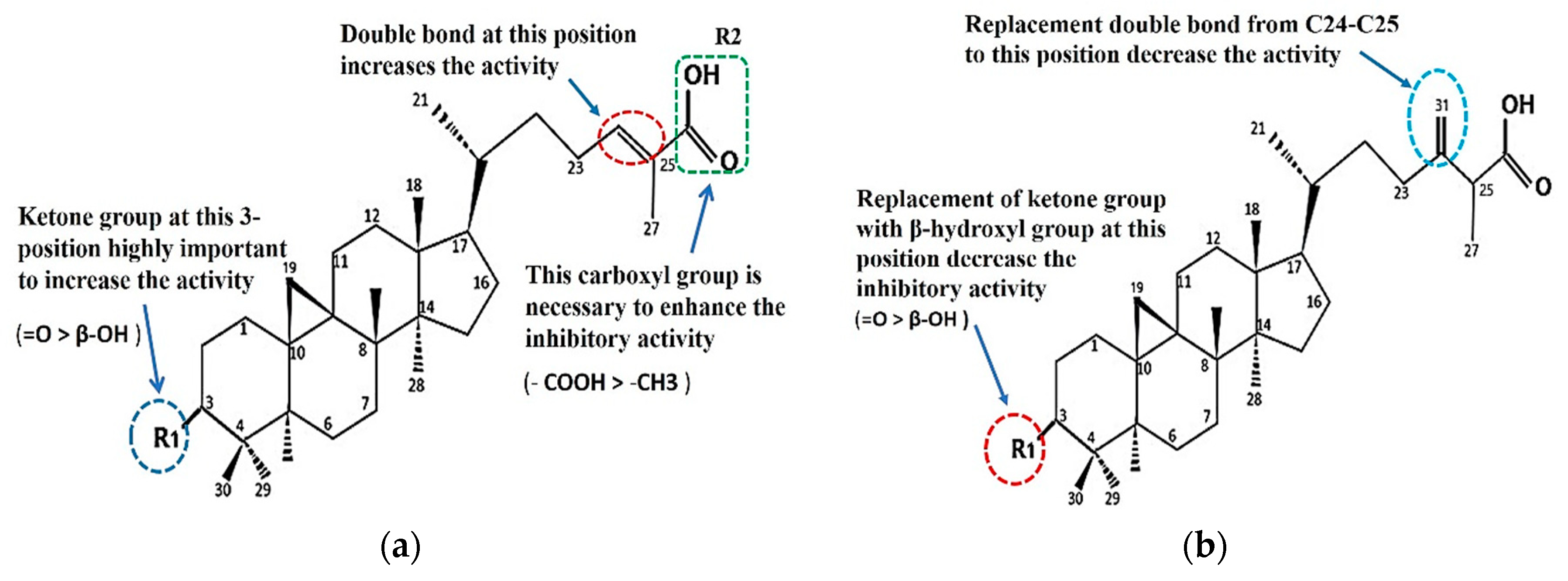
2.4. Structure–Activity Relationship of the Cycloartane-Type Triterpenes Isolated from T. sapiens Propolis and Their Inhibitory Activity against α-Glucosidase
3. Discussion
3.1. Phenolic Content and Antioxidant Properties of T. sapiens Propolis
3.2. Inhibitory Activity and Kinetic Inhibition of Cycloartane-Type Triterpenes Isolated from T. sapiens Bee Propolis on α-Glucosidases
3.3. Structure–Activity Relationship of the Cycloartane-Type Triterpenes Isolated from T. sapiens Propolis and Their Inhibitory Activity against α-Glucosidase
4. Materials and Methods
4.1. Sample Preparation
4.2. Total Phenolic Content
4.3. DPPH Free Radical Scavenging Activity
4.4. Inhibitory Activity Assay for Yeast α-Glucosidase
4.5. Inhibitory Activity Assay for Rat Intestinal α-Glucosidase
4.6. Determination of Inhibitory Type by Kinetic Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Diabetes Mellitus. WHO. Available online: https://www.who.int/ mediacentre/factsheets/fs138/en/ (accessed on 18 April 2019).
- International Diabetes Federation. IDF Diabetes Atlas, 8th ed.; International Diabetes Federation: Brussels, Belgium, 2017; pp. 14–24. Available online: http://www.diabetesatlas.org (accessed on 18 April 2019).
- Yin, Z.; Zhang, W.; Feng, F.; Zhang, Y.; Kang, W. α-Glucosidase inhibitors isolated from medicinal plants. Food Sci. Hum. Wellness 2014, 3, 136–174. [Google Scholar] [CrossRef]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of α-glucosidase, α-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Cao, J.; Duan, L.; Tang, Y.; Zhao, Y. Protein tyrosine phosphatase 1B (PTP1B) and α-glucosidase inhibitory activities of Schisandra chinensis (Turcz.) Baill. J. Funct. Foods 2014, 9, 264–270. [Google Scholar] [CrossRef]
- Nyemb, J.N.; Tchinda, A.T.; Talla, E.; Nanga, E.B.; Ngoudjou, D.T. A New Cerebroside from Vitellaria paradoxa (Sapotaceae) and its Bioactivities. Nat. Prod. Chem. Res. 2018, 6, 1–9. [Google Scholar] [CrossRef]
- Nhiem, N.X.; Kiem, P.V.; Minh, C.V.; Ban, N.K.; Cuong, N.X.; Ha, L.M.; Tai, B.H.; Quang, T.H.; Tung, N.H.; Kim, Y.H. Cucurbitane-type triterpene glycosides from the fruits of Momordica charantia. Magn. Reson. Chem. 2010, 48, 392–396. [Google Scholar]
- Marcucci, M.C. Propolis: Chemical composition, biological properties and therapeutic activity. Apidologie 1995, 26, 83–99. [Google Scholar] [CrossRef]
- Burdock, G.A. Review of the biological properties and toxicity of bee propolis (propolis). Food Chem. Toxicol. 1998, 36, 347–363. [Google Scholar] [CrossRef]
- Falcão, S.I.; Vale, N.; Cos, P.; Gomes, P.; Freire, C.; Maes, L.; Vilas-Boas, M. In Vitro Evaluation of Portuguese Propolis and Floral Sources for Antiprotozoal, Antibacterial and Antifungal Activity. Phytother. Res. 2014, 28, 437–443. [Google Scholar] [CrossRef]
- Boisard, S.; Le Ray, A.-M.; Landreau, A.; Kempf, M.; Cassisa, V.; Flurin, C.; Richomme, P. Antifungal and Antibacterial Metabolites from a French Poplar Type Propolis. Evid. Based Complement. Altern. Med. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Awale, S.; Tezuka, Y.; Kadota, S. Cytotoxic Constituents of Propolis from Myanmar and Their Structure-Activity Relationship. Biol. Pharm. Bull. 2009, 32, 2075–2078. [Google Scholar] [CrossRef] [Green Version]
- Elnakady, Y.A.; Rushdi, A.I.; Franke, R.; Abutaha, N.; Ebaid, H.; Baabbad, M.; Omar, M.O.M.; Al Ghamdi, A.A. Characteristics, chemical compositions and biological activities of propolis from Al-Bahah, Saudi Arabia. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Can, Z.; Yıldız, O.; Șahİn, H.; Asadov, A.; Kolaylı, S. Phenolic profile and antioxidant potential of propolis from Azerbaijan. Mellifera 2015, 15, 16–28. [Google Scholar]
- Rimbach, G.; Fischer, A.; Schloesser, A.; Jerz, G.; Ikuta, N.; Ishida, Y.; Matsuzawa, R.; Matsugo, S.; Huebbe, P.; Terao, K. Anti-Inflammatory Properties of Brazilian Green Propolis Encapsulated in a γ-Cyclodextrin Complex in Mice Fed a Western-Type Diet. Int. J. Mol. Sci. 2017, 18, 1141. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, H.M.; Anees, M.M.; Van Dyke, T.E. Propolis Improves Periodontal Status and Glycemic Control in Patients with Type 2 Diabetes Mellitus and Chronic Periodontitis: A Randomized Clinical Trial. J. Periodontol. 2016, 87, 1418–1426. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Chen, M.; Shou, Q.; Li, Y.; Hu, F. Biological Activities of Chinese Propolis and Brazilian Propolis on Streptozotocin-Induced Type 1 Diabetes Mellitus in Rats. Evid. Based Complement. Altern. Med. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pujirahayu, N.; Suzuki, T.; Katayama, T. Cycloartane-Type Triterpenes and Botanical Origin of Propolis of Stingless Indonesian Bee Tetragonula sapiens. Plants 2019, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Li, Y.; Zhang, Q.; Wei, X.; Peng, A.; Chen, L.; Wei, Y. Triterpene acids isolated from Lagerstroemia speciosa leaves as α-glucosidase inhibitors. Phytother. Res. 2009, 23, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Genet, C.; Strehle, A.; Schmidt, C.; Boudjelal, G.; Lobstein, A.; Schoonjans, K.; Souchet, M.; Auwerx, J.; Saladin, R.; Wagner, A. Structure−Activity Relationship Study of Betulinic Acid, A Novel and Selective TGR5 Agonist, and Its Synthetic Derivatives: Potential Impact in Diabetes. J. Med. Chem. 2010, 53, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xing, Y.; Wen, C.; Yu, X.; Sun, W.; Xiu, Z.; Dong, Y. Pentacyclic triterpenes as α-glucosidase and α-amylase inhibitors: Structure-activity relationships and the synergism with acarbose. Bioorg. Med. Chem. Lett. 2017, 27, 5065–5070. [Google Scholar] [CrossRef] [PubMed]
- Sancheti, S.; Sancheti, S.; Bafna, M.; Seo, S.-Y. 2,4,6-Trihydroxybenzaldehyde as a potent antidiabetic agent alleviates postprandial hyperglycemia in normal and diabetic rats. Med. Chem. Res. 2011, 20, 1181–1187. [Google Scholar] [CrossRef]
- Shibano, M.; Kakutani, K.; Taniguchi, M.; Yasuda, M.; Baba, K. Antioxidant constituents in the dayflower (Commelina communis L.) and their α-glucosidase-inhibitory activity. J. Nat. Med. 2008, 62, 349. [Google Scholar] [CrossRef] [PubMed]
- Dewi, R.T.; Tachibana, S.; Darmawan, A. Effect on α-glucosidase inhibition and antioxidant activities of butyrolactone derivatives from Aspergillus terreus MC751. Med. Chem. Res. 2014, 23, 454–460. [Google Scholar] [CrossRef]
- Anjaneyulu, V.; Satyanarayana, P.; Viswanadham, K.N.; Jyothi, V.G.; Rao, K.N.; Radhika, P. Triterpenoids from Mangifera indica. Phytochemistry 1999, 50, 1229–1236. [Google Scholar] [CrossRef]
- Freitas, M.O.; Ponte, F.A.F.; Lima, M.A.S.; Silveira, E.R. Flavonoids and triterpenes from the nest of the stingless bee Trigona spinipes. J. Braz. Chem. Soc. 2008, 19, 532–535. [Google Scholar] [CrossRef]
- Kardar, M.N.; Zhang, T.; Coxon, G.D.; Watson, D.G.; Fearnley, J.; Seidel, V. Characterisation of triterpenes and new phenolic lipids in Cameroonian propolis. Phytochemistry 2014, 106, 156–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobedo-Martínez, C.; Concepción Lozada, M.; Hernández-Ortega, S.; Villarreal, M.L.; Gnecco, D.; Enríquez, R.G.; Reynolds, W. 1H and 13C NMR characterization of new cycloartane triterpenes from Mangifera indica: Cycloartane triterpenes from Mangifera indica. Magn. Reson. Chem. 2012, 50, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Mangunwidjaja, D.; Sunarti, T.; Suparno, O.; Setiyono, A. Investigating the Antioxidant and Anticytotoxic Activities of Propolis Collected from Five Regions of Indonesia and Their Abilities to Induce Apoptosis. Emir. J. Food Agric. 2014, 26, 390–398. [Google Scholar] [CrossRef]
- Miguel, M.C. Chemical and biological properties of propolis from the western countries of the mediterranean basin and Portugal. Int. J. Pharm. Pharm. Sci. 2013, 5, 403–409. [Google Scholar]
- Sforcin, J.M.; Bankova, V. Propolis: Is there a potential for the development of new drugs? J. Ethnopharmacol. 2011, 133, 253–260. [Google Scholar] [CrossRef]
- Shai, L.J.; Masoko, P.; Mokgotho, M.P.; Magano, S.R.; Mogale, A.M.; Boaduo, N.; Eloff, J.N. Yeast alpha glucosidase inhibitory and antioxidant activities of six medicinal plants collected in Phalaborwa, South Africa. S. Afr. J. Bot. 2010, 76, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Hamid, K.; Alqahtani, A.; Kim, M.-S.; Cho, J.-L.; Cui, P.H.; Li, C.G.; Groundwater, P.W.; Li, G.Q. Tetracyclic Triterpenoids in Herbal Medicines and their Activities in Diabetes and its Complications. Curr. Top. Med. Chem. 2015, 15, 2406–2430. [Google Scholar] [CrossRef] [PubMed]
- Aichour, S.; Haba, H.; Benkhaled, M.; Harakat, D.; Lavaud, C. Terpenoids and other constituents from Euphorbia bupleuroides. Phytochem. Lett. 2014, 10, 198–203. [Google Scholar] [CrossRef]
- Suzuki, T.; Matsuno, F.; Katayama, T.; Andrianto, D. α-Glucosidase Inhibition and DPPH free radical—Scavenging activities of underutilized fruits from Java Island, Indonesia. J. For. Biomass Util Soc. 2017, 12, 59–62. [Google Scholar]
- Kim, K.Y.; Nam, K.A.; Kurihara, H.; Kim, S.M. Potent α-glucosidase inhibitors purified from the red alga Grateloupia elliptica. Phytochemistry 2008, 69, 2820–2825. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.-N.; Shin, J.-G.; Jang, H.-D. Antioxidant and antidiabetic activity of Dangyuja (Citrus grandis Osbeck) extract treated with Aspergillus saitoi. Food Chem. 2009, 117, 35–41. [Google Scholar] [CrossRef]
- Noor, A.; Bansal, V.S.; Vijayalakshmi, M.A. Current update on anti-diabetic biomolecules from key traditional Indian medicinal plants. Curr. Sci. 2013, 104, 721–727. [Google Scholar]
- Honda, M.; Hara, Y. Inhibition of Rat Small Intestinal Sucrase and α-Glucosidase Activities by Tea Polyphenols. Biosci. Biotechnol. Biochem. 1993, 57, 123–124. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Jeong, Y.-K.; Wang, M.-H.; Lee, W.-Y.; Rhee, H.-I. Inhibitory effect of pine extract on α-glucosidase activity and postprandial hyperglycemia. Nutrition 2005, 21, 756–761. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Lerdsuwankij, O.; Poputtachai, U.; Minipun, A.; Suparpprom, C. Inhibitory Activity of Cinnamon Bark Species and their Combination Effect with Acarbose against Intestinal α-glucosidase and Pancreatic α-amylase. Plant Foods Hum. Nutr. 2011, 66, 143–148. [Google Scholar] [CrossRef]
- Harrison, D.E.; Strong, R.; Allison, D.B.; Ames, B.N.; Astle, C.M.; Atamna, H.; Fernandez, E.; Flurkey, K.; Javors, M.A.; Nadon, N.L.; et al. Acarbose, 17-α-estradiol, and nordihydroguaiaretic acid extend mouse lifespan preferentially in males. Aging Cell 2014, 13, 273–282. [Google Scholar] [CrossRef]
- Misbah, H.; Aziz, A.A.; Aminudin, N. Antidiabetic and antioxidant properties of Ficus deltoidea fruit extracts and fractions. BMC Complement. Altern. Med. 2013, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, R.R.; Tipton, F.K. Assessment of Enzyme Inhibition: A Review with Examples from the Development of Monoamine Oxidase and Cholinesterase Inhibitory Drugs. Molecules 2017, 22, 1192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, G.; Beta, T.; Dong, J. Inhibitory Properties of Aqueous Ethanol Extracts of Propolis on Alpha-Glucosidase. Evid. Based Complement. Altern. Med. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghadyale, V.; Takalikar, S.; Haldavnekar, V.; Arvindekar, A. Effective Control of Postprandial Glucose Level through Inhibition of Intestinal Alpha Glucosidase by Cymbopogon martinii (Roxb.). Evid. Based Complement. Altern. Med. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.X.; Le, T.C.; Do, T.N.V.; Le, T.H.; Nguyen, N.T.; Nguyen, M.T.T. α-Glucosidase inhibitors from the bark of Mangifera mekongensis. Chem. Cent. J. 2016, 10, 1–6. [Google Scholar] [CrossRef]
- Suzuki, T.; Eto, K.; Kubota, Y.; Katayama, T.; Pankasemsuk, T. Antioxidative catechol lignans/neolignans isolated from defatted kernel of Jatropha curcas. J. Wood Sci. 2016, 62, 339–348. [Google Scholar] [CrossRef]
- Sivasothy, Y.; Loo, K.Y.; Leong, K.H.; Litaudon, M.; Awang, K. A potent α-glucosidase inhibitor from Myristica cinnamomea King. Phytochemistry 2016, 122, 265–269. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Extract & Fractions | Weigh/Yield (g) | % |
|---|---|---|---|
| 1. | Raw propolis of P1 | 10.00 | |
| Ethanol Extract of Propolis (EEP) of P1 | 5.66 | 56.6 | |
| Diethyl ether fr. of EEP P1 | 4.87 | 86.0 | |
| Ethyl acetate (EtOAc) fr. of EEP P1 | 0.09 | 1.59 | |
| Aqueous fr. of EEP P1 | 0.56 | 9.89 | |
| 2. | Raw propolis of P2 | 15.00 | |
| EEP of P2 | 9.97 | 66.5 | |
| Diethyl ether fr. of EEP P2 | 7.35 | 73.7 | |
| Ethyl acetate (EtOAc) fr. of EEP P2 | 0.51 | 5.11 | |
| Aqueous fr. of EEP P2 | 0.57 | 5.71 |
| Sample | Phenolic Acid (GAE) (µg/mg) a | Antioxidant Activity IC50 (µg/mL) a |
|---|---|---|
| P1 ether Fraction | 16.7 ± 0.218 | 64.9 ± 4.30 |
| P1 EtOAc fraction | 21.5 ± 0.271 | 25.0 ± 3.03 |
| P1 aqueous fraction | 24.4 ± 0.612 | 19.7 ± 0.229 |
| P2 ether Fraction | 32.4 ± 0.311 | 25.4 ± 3.78 |
| P2 EtOAc Fraction | 37.2 ± 0.468 | 32.44 ± 6.32 |
| P2 aqueous fraction | 13.9 ± 0.198 | 84.3 ± 9.71 |
| Trolox | - | 6.76 ± 0.395 |
| Compounds | DPPH | |
|---|---|---|
| IC50 (µg/mL) a | IC50 (µM) a | |
| Mangiferolic acid (1) | 12.38 ± 3.0 | 27.11 ± 6.57 |
| Cycloartenol (2) | 120.21 ± 11.2 | 282.18 ± 26.3 |
| Ambonic acid (3) | 42.45 ± 4.5 | 90.57 ± 9.60 |
| Mangiferonic acid (4) | 17.16 ± 2.98 | 37.74 ± 6.55 |
| Ambolic acid (5) | 132.30 ± 13.9 | 281.30 ± 29.5 |
| Trolox b | 7.17 ± 0.13 | 28.62 ± 0.52 |
| Compounds | Yield (mg) | IC50 | Inhibition Mode | |
|---|---|---|---|---|
| S. cerevisiae (µM) a | Rat Small Intestine (µM) a | |||
| Mangiferolic acid (1) | 20.6 | 5.52 ± 0.04 | ND | Uncompetitive |
| Cycloartenol (2) | 5.9 | 10.72 ± 0.28 | ND | Uncompetitive |
| Ambonic acid (3) | 42.4 | 3.01 ± 1.26 | ND | Mixed inhibition |
| Mangiferonic acid (4) | 68.5 | 2.46 ± 0.70 | ND | Mixed inhibition |
| Ambolic acid (5) | 24.8 | 4.31 ± 0.04 | ND | Mixed inhibition |
| (–)-Epicatechin c | 1991.1 ± 89.9 | ND | ||
| Acarbose c | ND b | 208.95 ± 0.96 | ||
| Voglibose c | ND | 78.57 ± 1.27 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pujirahayu, N.; Bhattacharjya, D.K.; Suzuki, T.; Katayama, T. α-Glucosidase Inhibitory Activity of Cycloartane-Type Triterpenes Isolated from Indonesian Stingless Bee Propolis and Their Structure–Activity Relationship. Pharmaceuticals 2019, 12, 102. https://0-doi-org.brum.beds.ac.uk/10.3390/ph12030102
Pujirahayu N, Bhattacharjya DK, Suzuki T, Katayama T. α-Glucosidase Inhibitory Activity of Cycloartane-Type Triterpenes Isolated from Indonesian Stingless Bee Propolis and Their Structure–Activity Relationship. Pharmaceuticals. 2019; 12(3):102. https://0-doi-org.brum.beds.ac.uk/10.3390/ph12030102
Chicago/Turabian StylePujirahayu, Niken, Debu Kumar Bhattacharjya, Toshisada Suzuki, and Takeshi Katayama. 2019. "α-Glucosidase Inhibitory Activity of Cycloartane-Type Triterpenes Isolated from Indonesian Stingless Bee Propolis and Their Structure–Activity Relationship" Pharmaceuticals 12, no. 3: 102. https://0-doi-org.brum.beds.ac.uk/10.3390/ph12030102