Larvicidal and Enzymatic Inhibition Effects of Annona Muricata Seed Extract and Main Constituent Annonacin against Aedes Aegypti and Aedes Albopictus (Diptera: Culicidae)

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Seed Collection and Obtaining the Ethanolic Extract of Annona muricata
2.2. Obtaining the Acetogenin-Rich Fraction of A. muricata Seeds
2.3. Analysis of the Acetogenin-Rich Fraction by HPLC and Separation of Its Main Constituent
2.4. Collection and Maintenance of Ae. aegypti and Ae. albopictus Mosquitoes
2.5. Larvicidal Assay
2.6. Enzymatic Effect of the Acetogenin-Rich Fraction and Annonacin of A. muricata on the Mosquito Larvae
2.7. Statistical Analysis
3. Results and Discussion
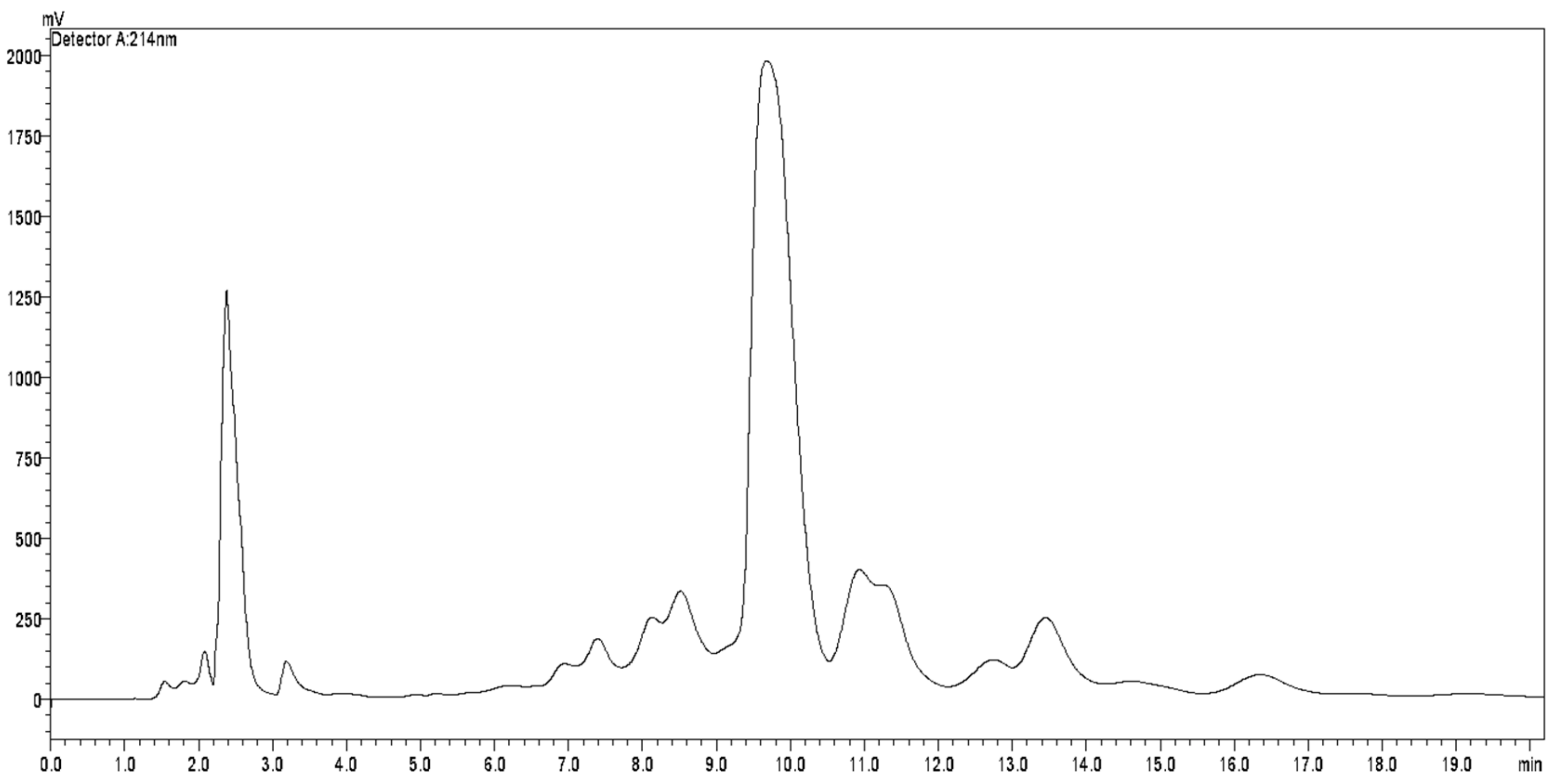
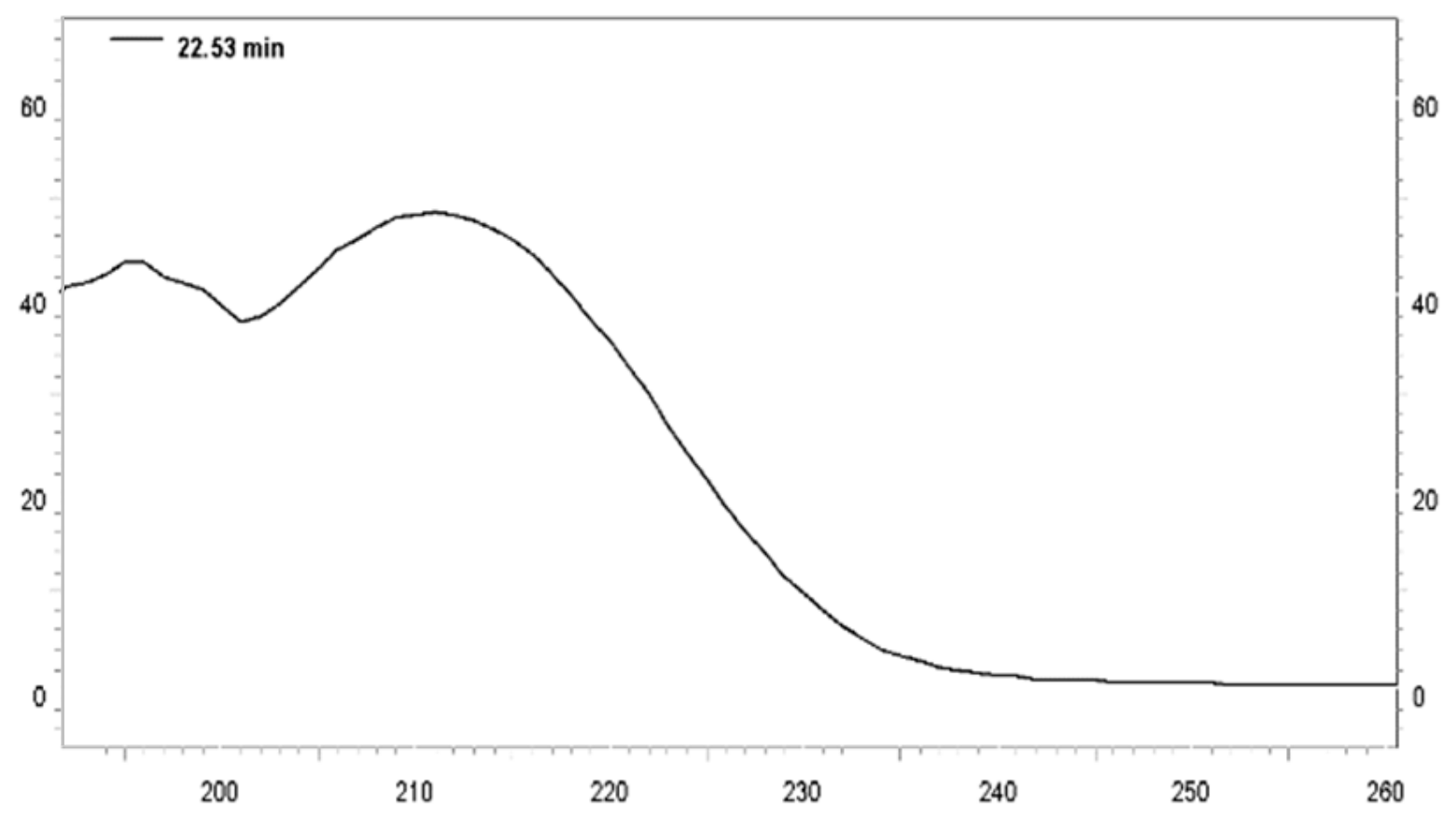
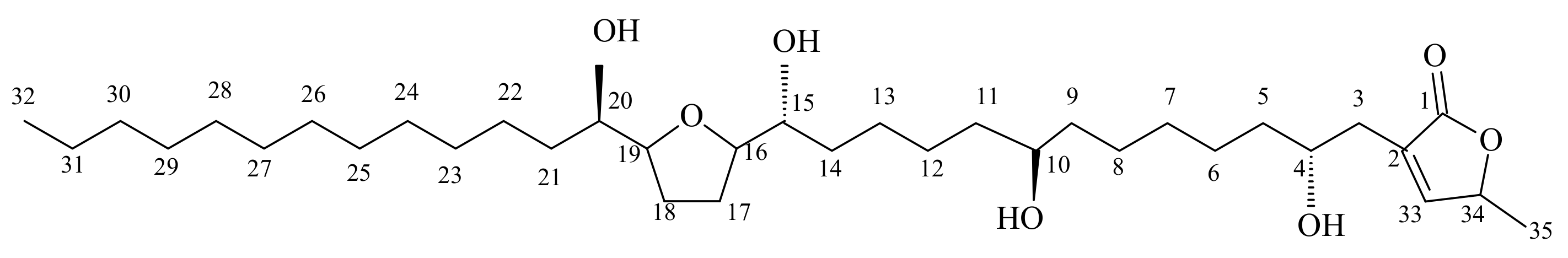
3.1. Analysis of the Constituents of the Acetogenin-Rich Fraction (ACERF)
3.2. Efficacy of the ACERF and Annonacin of A. muricata Seeds on Ae. aegypti and Ae. albopictus
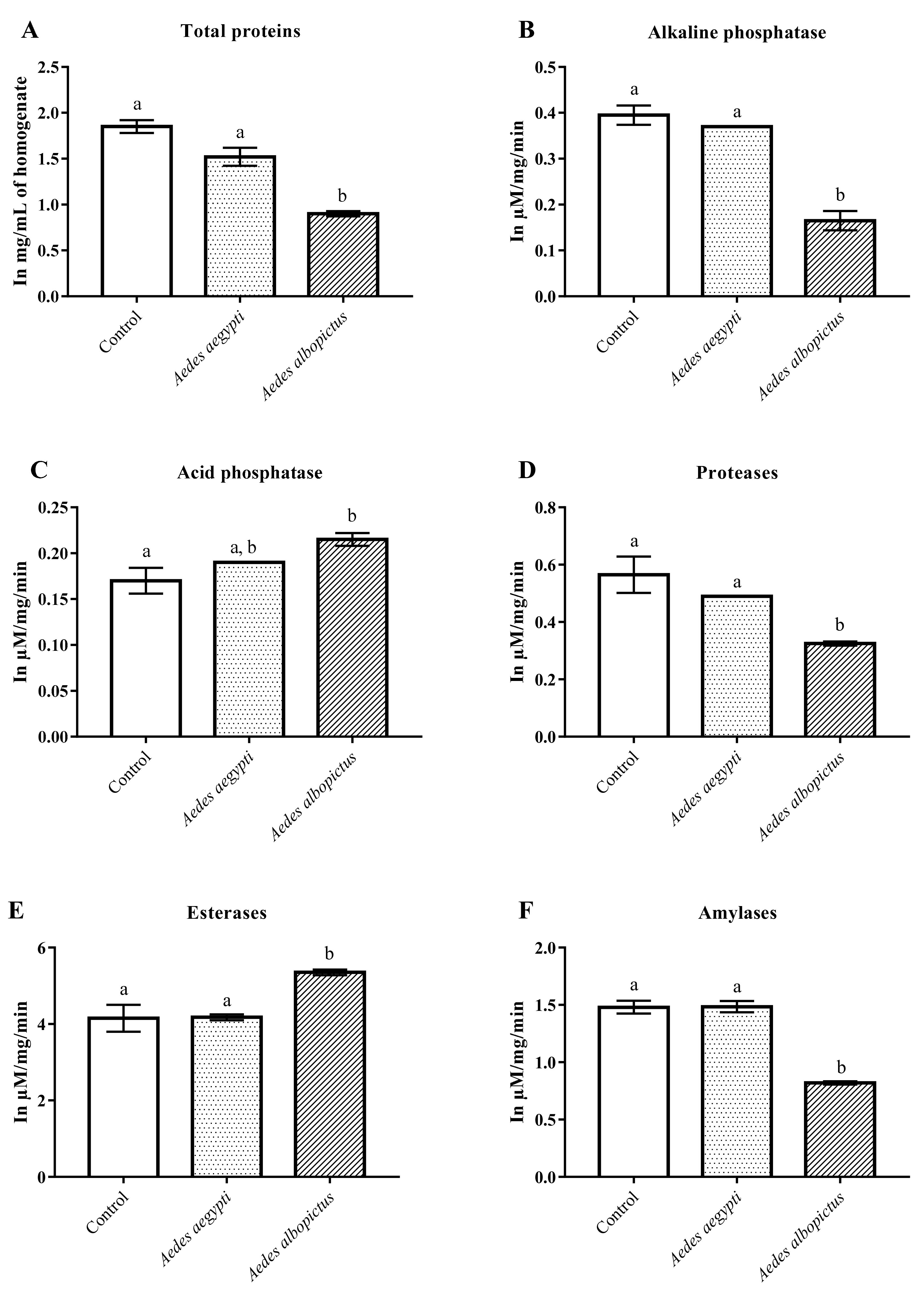
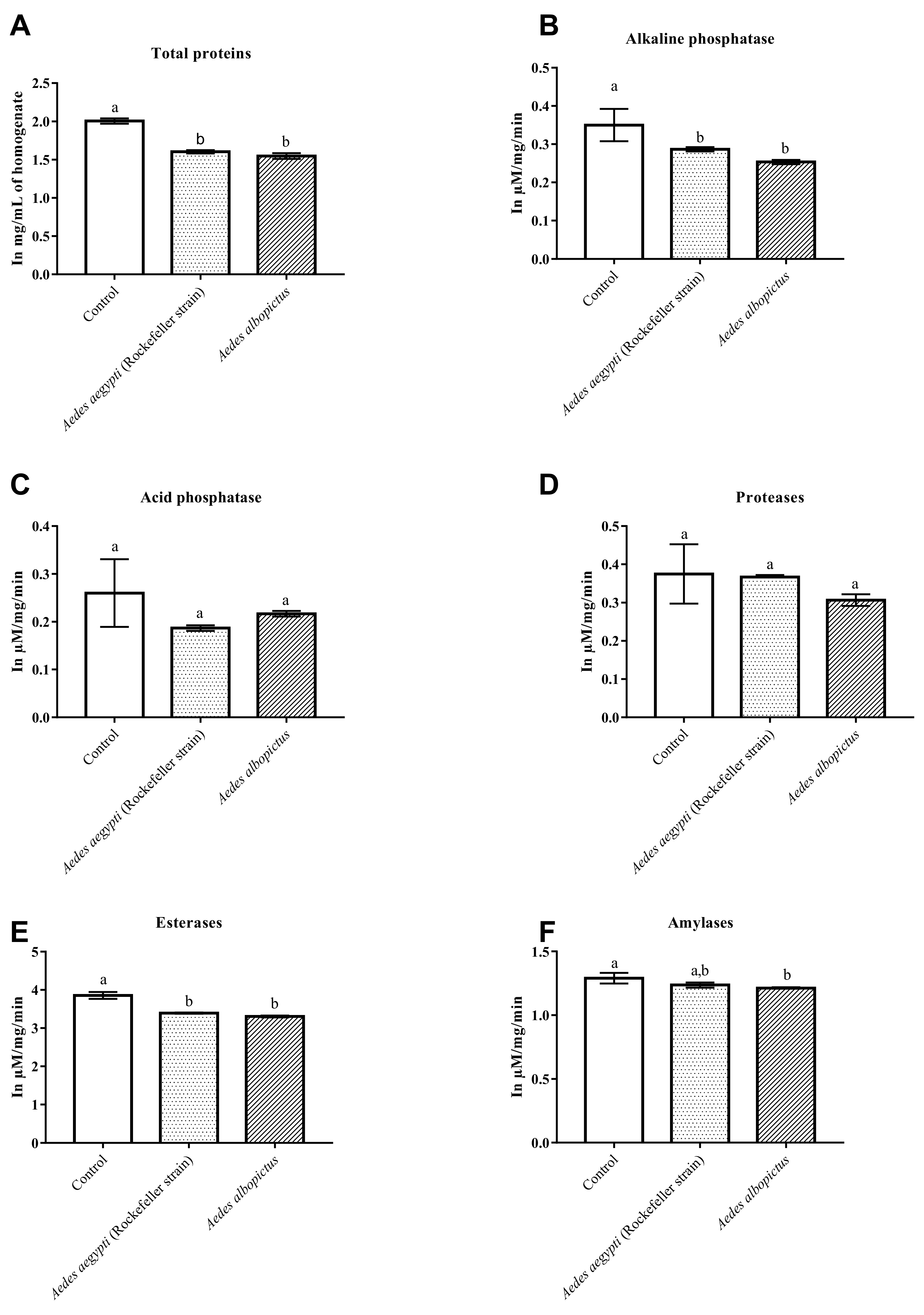
3.3. Changes in the Enzymatic Level of Ae. aegypti and Ae. albopictus Treated with ACERF and Annonacin
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fujiwara, G.M.; Annies, V.; De Oliveira, C.F.; Lara, R.A.; Gabriel, M.M.; Betim, F.C.; Nadal, J.M.; Farago, P.V.; Dias, J.F.; Miguel, O.G.; et al. Evaluation of larvicidal activity and ecotoxicity of linalool, methyl cinnamate and methyl cinnamate/linalool in combination against Aedes aegypti. Ecotoxicol. Environ. Saf. 2017, 139, 238–244. [Google Scholar] [CrossRef]
- De Oliveira, A.P.S.; Silva, L.L.D.S.; Lima, T.D.A.; Pontual, E.V.; Santos, N.D.D.L.; Coelho, L.C.B.B.; Navarro, D.M.D.A.F.; Zingali, R.B.; Napoleão, T.H.; Paiva, P.M.G. Biotechnological value of Moringa oleifera seed cake as source of insecticidal lectin against Aedes aegypti. Process. Biochem. 2016, 51, 1683–1690. [Google Scholar] [CrossRef]
- Pezzi, M.; Zamberlan, F.; Chicca, M.; Leis, M. Evaluation of larvicidal activity of esters of 4-mercapto-2-butenoic acid against Aedes albopictus (Diptera: Culicidae). Saudi J. Boil. Sci. 2018. [Google Scholar] [CrossRef]
- Martins, V.E.P.; Martins, M.G.; De Araújo, J.M.P.; Silva, L.O.R.; Monteiro, H.A.D.O.; Castro, F.C.; Vasconcelos, P.F.D.C.; Guedes, M.I.F. First report of Aedes (Stegomyia) albopictus in the state of Ceará, Brazil. Rev. Saúde Pública 2006, 40, 737–739. [Google Scholar] [CrossRef] [PubMed]
- Dayan, G.H.; Rivera, D.M.; Deseda, C.; Costa, M.S.; Morales-Ramírez, J.O.; Rey, L.C.; Dietze, R.; Luz, K.; Rivas, E.; Zambrano, B.; et al. Efficacy of a Tetravalent Dengue Vaccine in Children in Latin America. N. Engl. J. Med. 2015, 372, 113–123. [Google Scholar]
- Karunaratne, S.; Weeraratne, T.; Perera, M.; Surendran, S. Insecticide resistance and, efficacy of space spraying and larviciding in the control of dengue vectors Aedes aegypti and Aedes albopictus in Sri Lanka. Pestic. Biochem. Physiol. 2013, 107, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, V.C.; O Scherer, P.; Falcão, S.S.; Alencar, J.; Cunha, S.P.; Rodrigues, I.M.; Pinheiro, N.L. Diversidade de criadouros e tipos de imóveis freqüentados por Aedes albopictus e Aedes aegypti. Rev. Saúde Pública 2006, 40, 1106–1111. [Google Scholar] [CrossRef]
- Rueda, A.G.; Otero, A.L.C.; Duque, J.E.; Kouznetsov, V.V. Synthesis of new α-amino nitriles with insecticidal action on Aedes aegypti (Diptera: Culicidae). Rev. Bras. Èntomol. 2018, 62, 112–118. [Google Scholar] [CrossRef]
- Braga, I.A.; Valle, D. Aedes aegypti: Inseticidas, mecanismos de ação e resistência. Epidemiol. Serviços Saúde 2007, 16, 279–293. [Google Scholar] [CrossRef]
- Park, H.-M.; Kim, J.; Chang, K.-S.; Kim, B.-S.; Yang, Y.-J.; Kim, G.-H.; Shin, S.-C.; Park, I.-K. Larvicidal activity of Myrtaceae essential oils and their components against Aedes aegypti, acute toxicity on Daphnia magna, and aqueous residue. J. Med. Èntomol. 2011, 48, 405–410. [Google Scholar] [CrossRef]
- Ali, S.I.; Gopalakrishnan, B.; Venkatesalu, V. Chicory (Cichorium intybus) and wormwood (Artemisia absinthium) extracts exhibit strong larvicidal activity against mosquito vectors of malaria, dengue fever, and filariasis. Parasitol. Int. 2018, 67, 781–786. [Google Scholar] [CrossRef]
- Mendes, L.A.; Martins, G.F.; Valbon, W.R.; De-Souza, T.D.S.; Menini, L.; Ferreira, A.; Ferreira, M.F.C. Larvicidal effect of essential oils from Brazilian cultivars of guava on Aedes aegypti L. Ind. Crops Prod. 2017, 108, 684–689. [Google Scholar] [CrossRef]
- Oliveira, A.E.M.F.M.; Bezerra, D.C.; Duarte, J.L.; Cruz, R.A.S.; Souto, R.N.P.; Ferreira, R.M.A.; Nogueira, J.; Conceição, E.C.; Leitão, S.; Bizzo, H.R.; et al. Essential oil from Pterodon emarginatus as a promising natural raw material for larvicidal nanoemulsions against a tropical disease vector. Sustain. Chem. 2017, 6, 1–9. [Google Scholar] [CrossRef]
- Piplani, M.; Bhagwat, D.P.; Singhvi, G.; Sankaranarayanan, M.; Balana-Fouce, R.; Vats, T.; Chander, S. Plant-based larvicidal agents: An overview from 2000 to 2018. Exp. Parasitol. 2019, 199, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Ramanibai, R.; Parthiban, E.; Boothapandi, M. Effect of seed kernel aqueous extract from Annona squamosa against three mosquito vectors and its impact on non-target aquatic organisms. Asian Pac. J. Trop. Dis. 2016, 6, 741–745. [Google Scholar] [CrossRef]
- Ribeiro, L.P.; Vendramim, J.D.; Bicalho, K.U.; Andrade, M.S.; Fernandes, J.B.; Moral, R.A.; Demétrio, C.G.B. Annona mucosa Jacq. (Annonaceae): A promising source of bioactive compounds against Sitophilus zeamais Mots. (Coleoptera: Curculionidae). J. Stored Prod. Res. 2013, 55, 6–14. [Google Scholar] [CrossRef] [Green Version]
- Bento, E.B.; Júnior, F.E.B.; De-Oliveira, D.R.; Fernandes, C.N.; De-Araújo-Delmondes, G.; Cesário, F.R.A.S.; Rodrigues, K.S.; Sales, V.S.; Figueiredo, F.R.S.D.N.; Lemos, I.C.S.; et al. Antiulcerogenic activity of the hydroalcoholic extract of leaves of Annona muricata Linnaeus in mice. Saudi J. Biol. Sci. 2018, 25, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Avula, B.; Bae, J.-Y.; Majrashi, T.; Wu, T.-Y.; Wang, Y.-H.; Wang, M.; Ali, Z.; Wu, Y.-C.; Khan, I.A. Targeted and non-targeted analysis of annonaceous alkaloids and acetogenins from Asimina and Annona species using UHPLC-QToF-MS. J. Pharm. Biomed. Anal. 2018, 159, 548–566. [Google Scholar] [CrossRef] [PubMed]
- Coria-Téllez, A.N.; Montalvo-Gónzalez, E.; Yahia, E.M.; Obledo-Vázquez, E.N. Annona muricata: A comprehensive review on its traditional medicinal uses, phytochemicals, pharmacological activities, mechanisms of action and toxicity. Arab. J. Chem. 2016, 11, 662–691. [Google Scholar] [CrossRef]
- Roduan, M.R.M.; Hamid, R.A.; Sulaiman, H.; Mohtarrudin, N. Annona muricata leaves extracts prevent DMBA/TPA-induced skin tumorigenesis via modulating antioxidants enzymes system in ICR mice. Biomed. Pharmacother. 2017, 94, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Liu, J.; Sun, X.; Zhu, W.; Yang, F.; Felczak, L.; Dou, Q.P.; Zhou, K. Novel Annonaceous acetogenins from Graviola (Annona muricata) fruits with strong anti-proliferative activity. Tetrahedron Lett. 2017, 58, 1895–1899. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Guidelines for Laboratory and Field Testing of Mosquito Larvicides. Available online: https://apps.who.int/iris/bitstream/handle/10665/69101/WHO_CDS_WHOPES_GCDPP_2005.13.pdf;jsessionid=ACA4BF09EDE97BB1D53DF249203894E0?sequence=1 (accessed on 5 April 2018).
- Suryawanshi, R.K.; Patil, C.D.; Borase, H.P.; Narkhede, C.P.; Salunke, B.K.; Patil, S.V. Mosquito larvicidal and pupaecidal potential of prodigiosin from Serratia marcescens and understanding its mechanism of action. Pestic. Biochem. Physiol. 2015, 123, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Rieser, M.J.; Gu, Z.-M.; Fang, X.-P.; Zeng, L.; Wood, K.V.; McLaughlin, J.L. Five Novel Mono-tetrahydrofuran Ring Acetogenins from the Seeds of Annona muricata. J. Nat. Prod. 1996, 59, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Mohankumar, T.K.; Shivanna, K.S.; Achuttan, V.V. Screening of Methanolic Plant Extracts against Larvae of Aedes aegypti and Anopheles stephensi in Mysore. J. Arthropod Borne Dis. 2016, 10, 303–314. [Google Scholar] [PubMed]
- Kempraj, V.; Bhat, S.K. Acute and reproductive toxicity of Annona squamosa to Aedes albopictus. Pestic. Biochem. Physiol. 2011, 100, 82–86. [Google Scholar] [CrossRef]
- Deomena, M.; Navarro, D.; DePaula, J.; Luna, J.; Ferreiradelima, M.; Santana, A. Larvicidal activities against Aedes aegypti of some Brazilian medicinal plants. Bioresour. Technol. 2007, 98, 2549–2556. [Google Scholar] [CrossRef]
- Cavalca, P.A.M.; Lolis, M.I.G.D.A.; Reis, B.; Bonato, C.M. Homeopathic and larvicide effect of Eucalyptus cinerea essential oil against Aedes aegypti. Braz. Arch. Boil. Technol. 2010, 53, 835–843. [Google Scholar] [CrossRef] [Green Version]
- Warikoo, R.; Ray, A.; Sandhu, J.K.; Samal, R.; Wahab, N.; Kumar, S. Larvicidal and irritant activities of hexane leaf extracts of Citrus sinensis against dengue vector Aedes aegypti L. Asian Pac. J. Trop. Biomed. 2012, 2, 152–155. [Google Scholar] [CrossRef] [Green Version]
- De Morais, S.M.; Facundo, V.A.; Bertini, L.M.; Cavalcanti, E.S.B.; Júnior, J.F.D.A.; Ferreira, S.A.; De Brito, E.S.; Neto, M.A.D.S. Chemical composition and larvicidal activity of essential oils from Piper species. Biochem. Syst. Ecol. 2007, 35, 670–675. [Google Scholar] [CrossRef]
- Kumar, S.; Nair, G.; Singh, A.P.; Batra, S.; Wahab, N.; Warikoo, R. Evaluation of the larvicidal efficiency of stem, roots and leaves of the weed, Parthenium hysterophorus (Family: Asteraceae) against Aedes aegypti L. Asian Pac. J. Trop. Dis. 2012, 2, 395–400. [Google Scholar] [CrossRef]
- Subramaniam, J.; Kovendan, K.; Kumar, P.M.; Murugan, K.; Walton, W. Mosquito larvicidal activity of Aloe vera (Family: Liliaceae) leaf extract and Bacillus sphaericus, against Chikungunya vector, Aedes aegypti. Saudi J. Boil. Sci. 2012, 19, 503–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahuman, A.A.; Gopalakrishnan, G.; Venkatesan, P.; Geetha, K. Larvicidal activity of some Euphorbiaceae plant extracts against Aedes aegypti and Culex quinquefasciatus (Diptera: Culicidae). Parasitol. Res. 2008, 102, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Bonneau, N.; Schmitz-Afonso, I.; Brunelle, A.; Touboul, D.; Champy, P. Method development for quantification of the environmental neurotoxin annonacin in Rat plasma by UPLC–MS/MS and application to a pharmacokinetic study. J. Chromatogr. B 2015, 1004, 46–52. [Google Scholar] [CrossRef] [PubMed]
- De Mendonça, F.A.; Da Silva, K.; Dos Santos, K.; Júnior, K.R.; Sant’Ana, A. Activities of some Brazilian plants against larvae of the mosquito Aedes aegypti. Fitoterapia 2005, 76, 629–636. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Zeng, Q.Y.; Shi, G.; Oberlies, N.H.; Zhao, G.; Njoku, C.J.; Mclaughlin, J.L. Comparative SAR evaluations of annonaceous acetogenins for pesticidal acitivity. J. Pest Sci. 1997, 49, 372–378. [Google Scholar] [CrossRef]
- Rodríguez-Cavallo, E.; Guarnizo-Méndez, J.; Yépez-Terrill, A.; Cárdenas-Rivero, A.; Diaz-Castillo, F.; Méndez-Cuadro, D. Protein carbonylation is a mediator in larvicidal mechanisms of Tabernaemontana cymosa ethanolic extract. J. King Saud Univ. Sci. 2018. [Google Scholar] [CrossRef]
- Costa, M.D.S.; De Paula, S.O.; Martins, G.F.; Zanuncio, J.C.; Santana, A.E.G.; Serrão, J.E. Modes of action of squamocin in the anal papillae of Aedes aegypti larvae. Physiol. Mol. Plant Pathol. 2018, 101, 172–177. [Google Scholar] [CrossRef]
- Santos, L.M.; Nascimento, J.S.; Santos, M.A.; Marriel, N.B.; Bezerra-Silva, P.C.; Rocha, S.K.; Silva, A.G.; Correia, M.T.; Paiva, P.M.; Martins, G.F.; et al. Fatty acid-rich volatile oil from Syagrus coronata seeds has larvicidal and oviposition-deterrent activities against Aedes aegypti. Physiol. Mol. Plant Pathol. 2017, 100, 35–40. [Google Scholar] [CrossRef]
- Wong, C.-L.; Ahmad, R.; Yu, K.-X.; Jantan, I. Larvicidal activity, inhibition effect on development, histopathological alteration and morphological aberration induced by seaweed extracts in Aedes aegypti (Diptera: Culicidae). Asian Pac. J. Trop. Med. 2015, 8, 1006–1012. [Google Scholar] [Green Version]
- Koodalingam, A.; Mullainadhan, P.; Arumugam, M. Effects of extract of soapnut Sapindus emarginatus on esterases and phosphatases of the vector mosquito, Aedes aegypti (Diptera: Culicidae). Acta Trop. 2011, 118, 27–36. [Google Scholar] [CrossRef]
- Shamsi, T.N.; Parveen, R.; Ahmad, A.; Samal, R.R.; Kumar, S.; Fatima, S. Inhibition of gut proteases and development of dengue vector, Aedes aegypti by Allium sativum protease inhibitor. Acta Ecol. Sin. 2018, 38, 325–328. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mosquito Strains | LC50 (CL at 95%) | LC90 (CL at 95%) |
|---|---|---|
| Ae. aegypti | 12.41 (9.85–14.86) | 30.21 (26.17–36.52) |
| Ae. albopictus | 3.41 (2.83–4.00) | 6.17 (5.37–7.43) |
| Ae. aegypti (Rockefeller strain) | 26.75 (21.36–34.00) | 54.46 (44.78–71.01) |
| Mosquito Strains | LC50 (CL at 95%) | LC90 (CL at 95%) |
|---|---|---|
| Ae. aegypti (Rockefeller strain) | 2.65 (1.87–3.64) | 4.83 (3.80–7.35) |
| Ae. albopictus | 8.34 (7.10–10.26) | 16.30 (13.56–21.13) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, A.M.; Silva, A.A.S.; Pinto, C.C.C.; Santos, D.L.d.; Freitas, J.C.C.d.; Martins, V.E.P.; Morais, S.M.d. Larvicidal and Enzymatic Inhibition Effects of Annona Muricata Seed Extract and Main Constituent Annonacin against Aedes Aegypti and Aedes Albopictus (Diptera: Culicidae). Pharmaceuticals 2019, 12, 112. https://0-doi-org.brum.beds.ac.uk/10.3390/ph12030112
Rodrigues AM, Silva AAS, Pinto CCC, Santos DLd, Freitas JCCd, Martins VEP, Morais SMd. Larvicidal and Enzymatic Inhibition Effects of Annona Muricata Seed Extract and Main Constituent Annonacin against Aedes Aegypti and Aedes Albopictus (Diptera: Culicidae). Pharmaceuticals. 2019; 12(3):112. https://0-doi-org.brum.beds.ac.uk/10.3390/ph12030112
Chicago/Turabian StyleRodrigues, Alzeir Machado, Antonio Adailson Sousa Silva, Cleonilda Claita Carneiro Pinto, Dayanne Lima dos Santos, José Claudio Carneiro de Freitas, Victor Emanuel Pessoa Martins, and Selene Maia de Morais. 2019. "Larvicidal and Enzymatic Inhibition Effects of Annona Muricata Seed Extract and Main Constituent Annonacin against Aedes Aegypti and Aedes Albopictus (Diptera: Culicidae)" Pharmaceuticals 12, no. 3: 112. https://0-doi-org.brum.beds.ac.uk/10.3390/ph12030112