Anticancer Activity of Amb4269951, a Choline Transporter-Like Protein 1 Inhibitor, in Human Glioma Cells



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
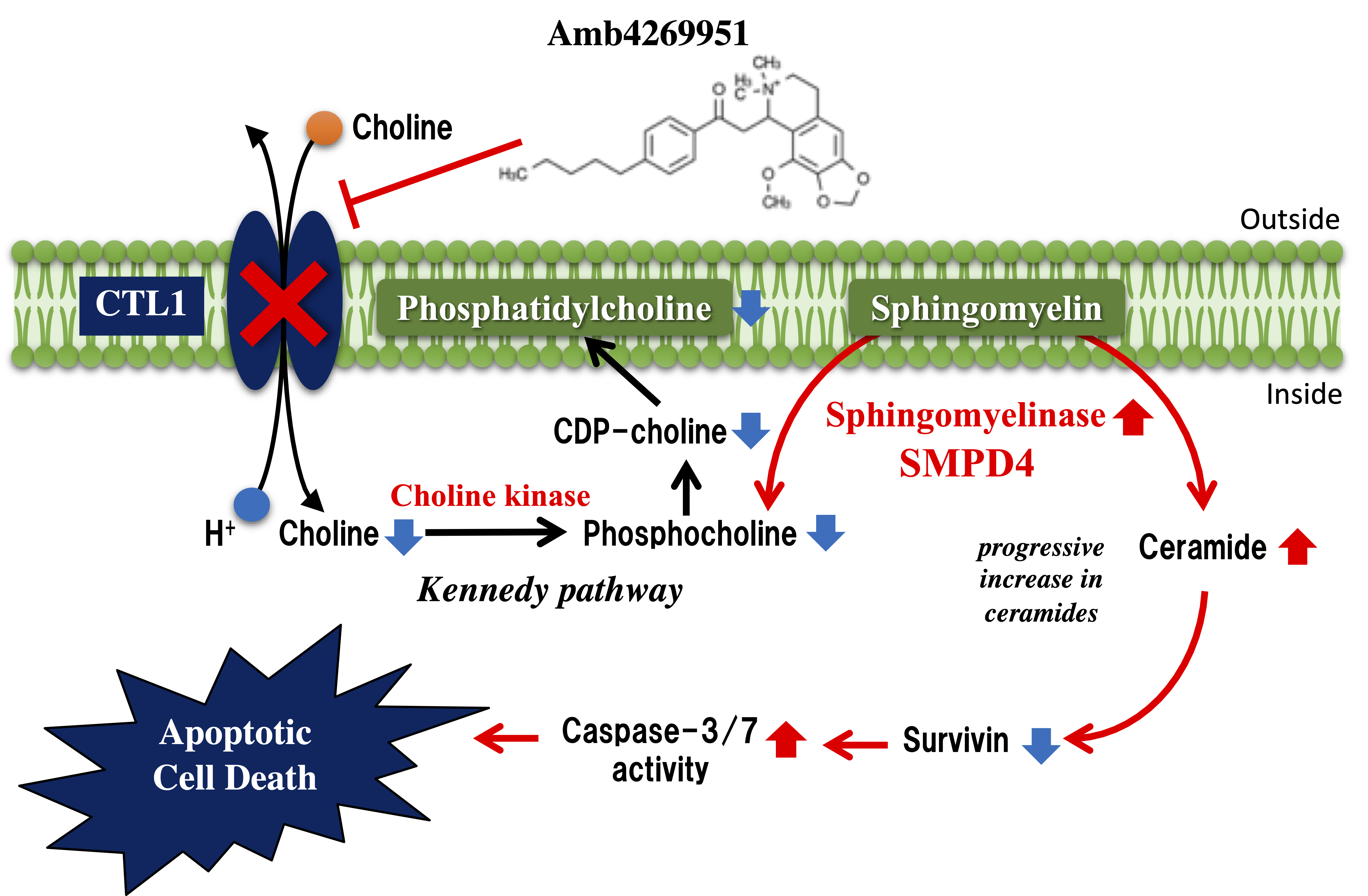
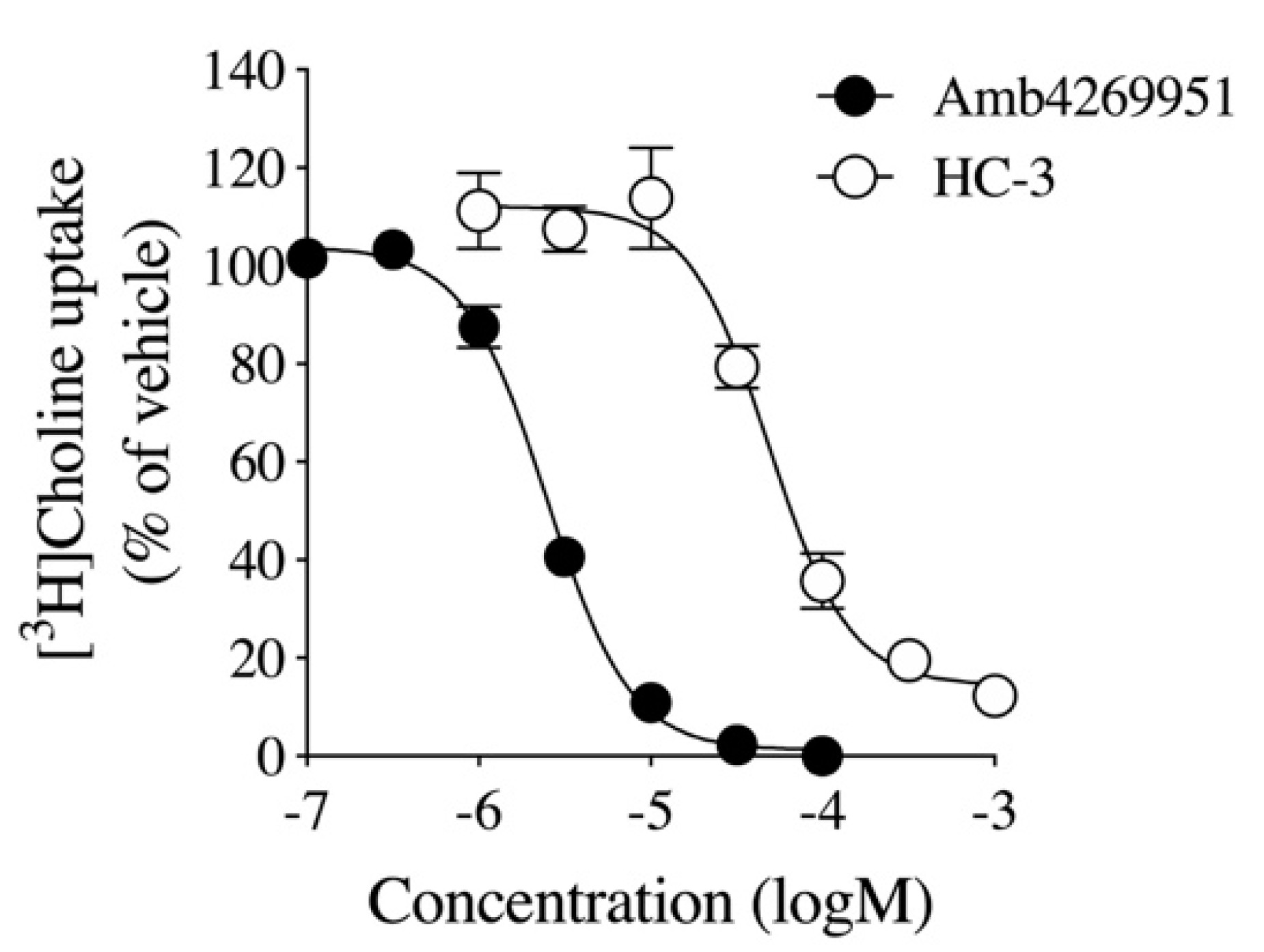
2.1. Effect of Amb4269951 and Hemicholinium-3 (HC-3) on [3H]Choline Uptake
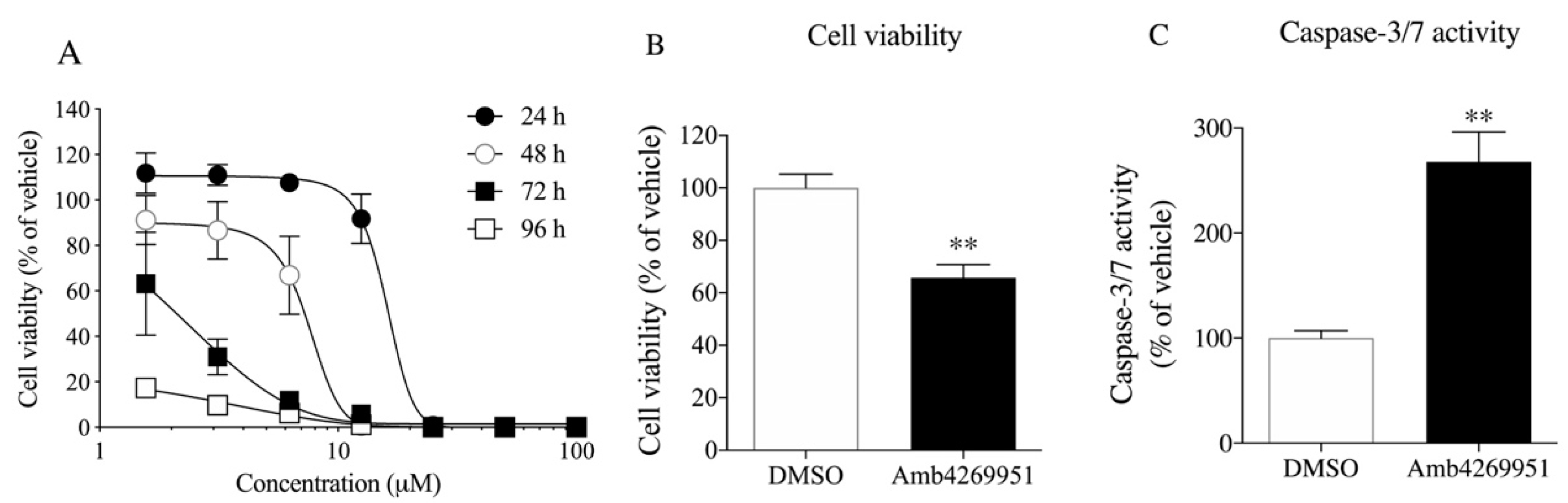
2.2. Effects of Amb4269951 on Cell Viability and Caspase-3/7 Activity
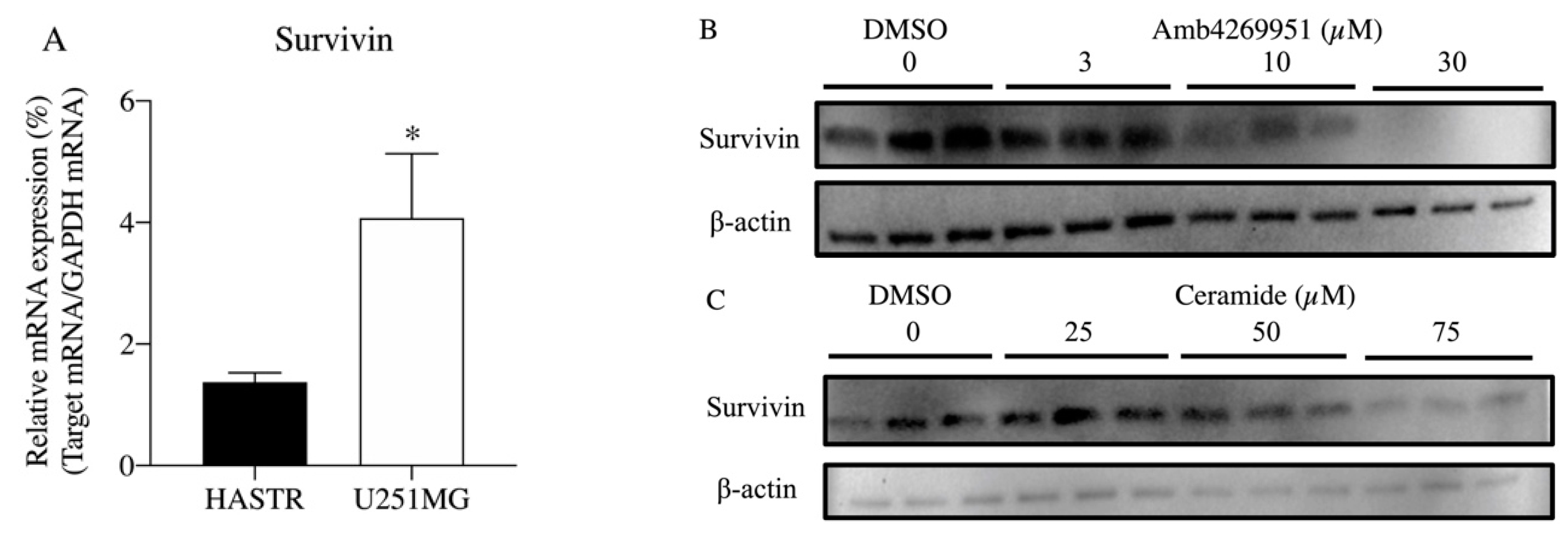
2.3. Effect of Amb4269951 on the Expression of Sphingomyelinases
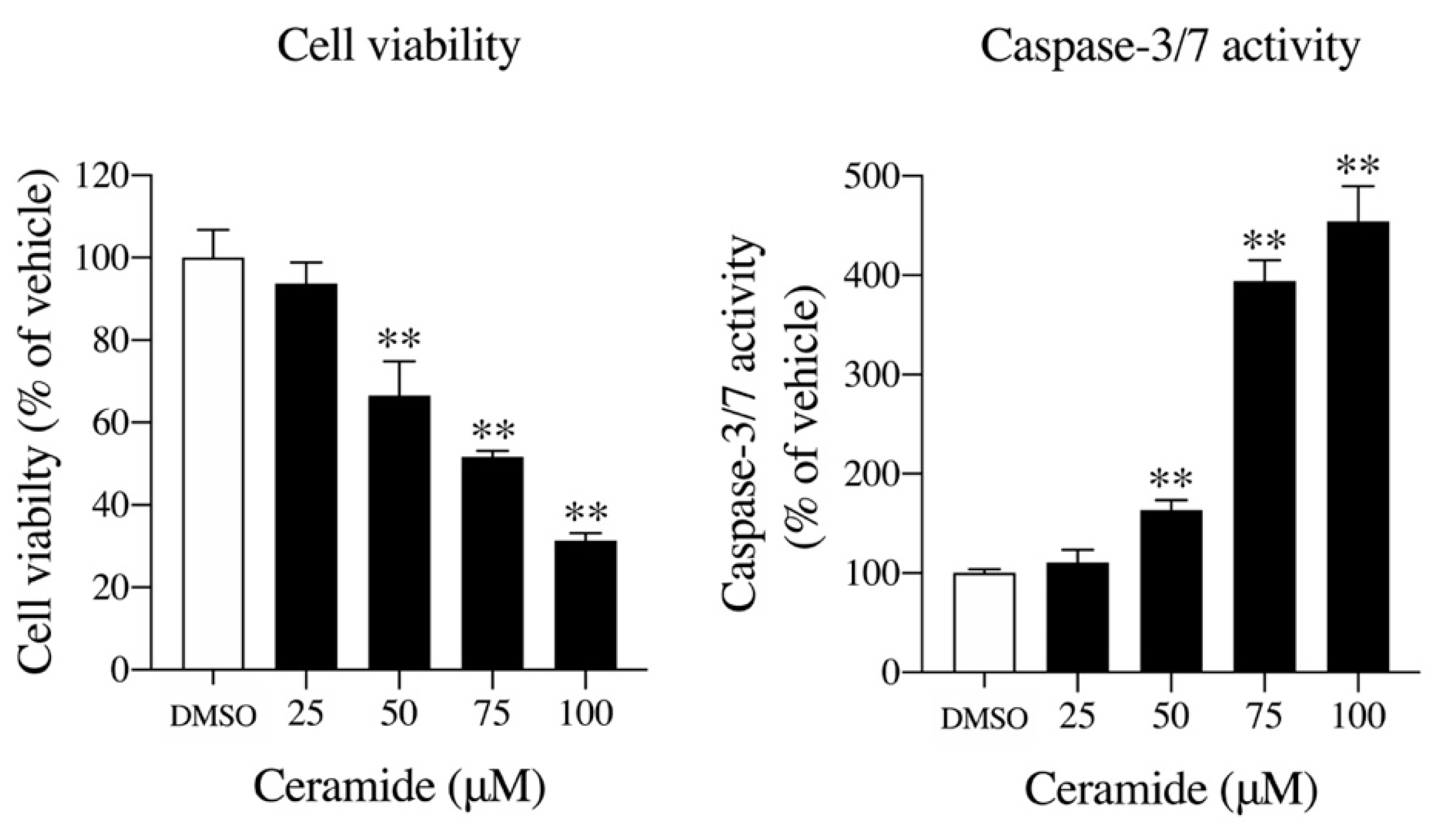
2.4. Effects of Ceramide on Cell Viability and Caspase-3/7 Activity
2.5. Effect of Amb4269951 and Ceramide on the Expression of Survivin
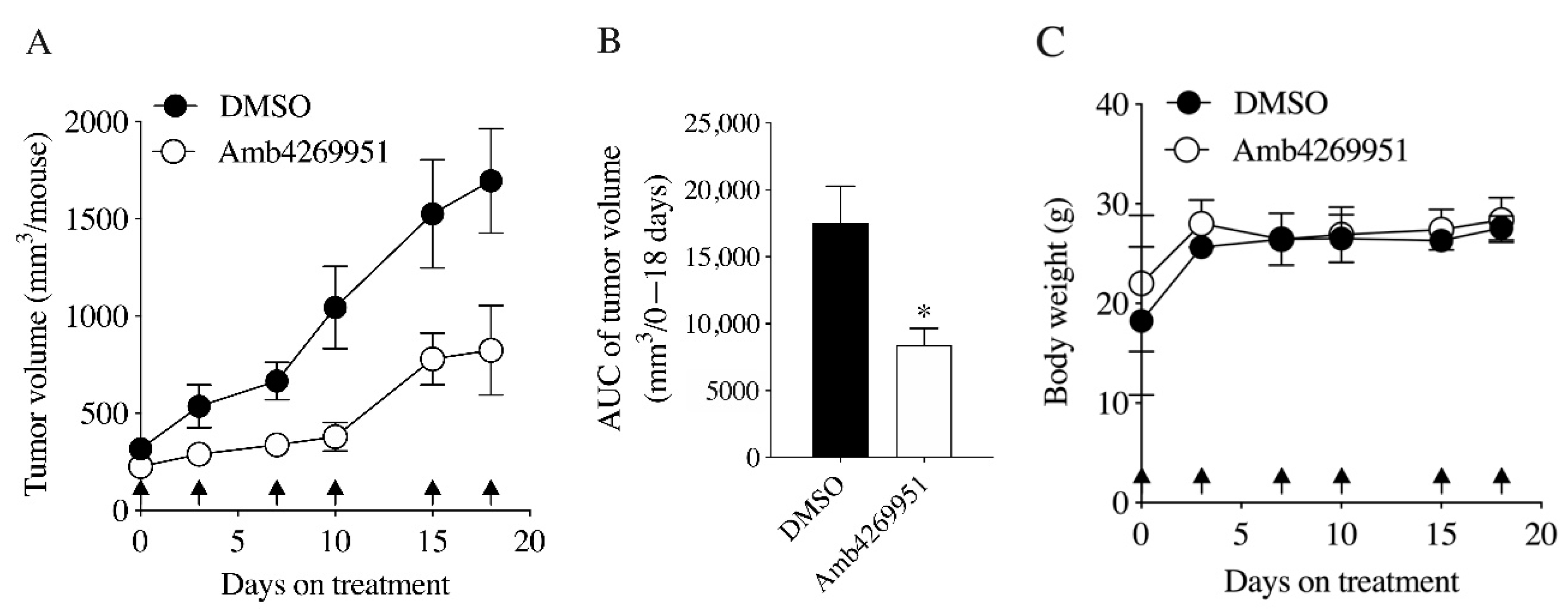
2.6. Effects of Amb4269951 in U251MG Cell Xenograft Model Mice
3. Discussion
4. Materials and Methods
4.1. Cell Culture
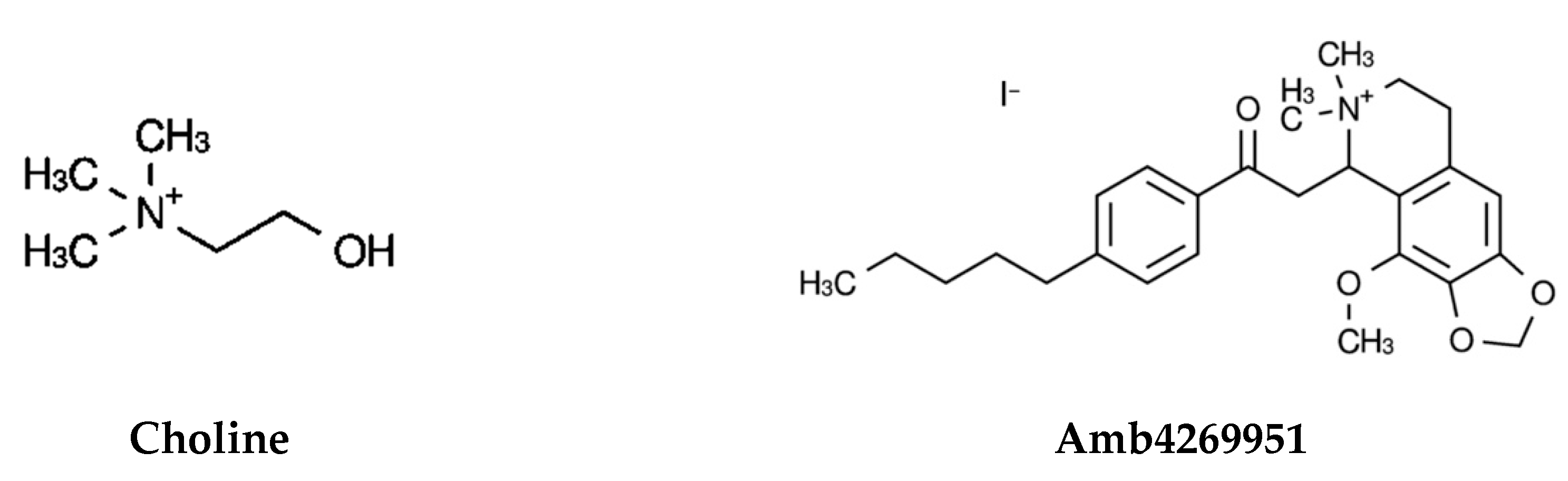
4.2. Isoquinoline Derivative Amb4269951
4.3. [3H]Choline Uptake in U251MG Cells
4.4. Cell Viability Assay
4.5. Measurements of Caspase-3/7 Activity
4.6. Quantification of mRNA by qPCR
4.7. Western Blot Analysis
4.8. Mouse Xenograft Model
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K. WHO Classification of Tumors of the Central Nervous System, Revised, 4th ed.; IARC Press: Lyon, France, 2016. [Google Scholar]
- Narita, Y.; Shibui, S. Trends and outcomes in the treatment of gliomas based on data during 2001-2004 from the Brain Tumor Registry of Japan. Neurol. Med. Chir. (Tokyo) 2015, 55, 286–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacinto, F.V.; Esteller, M. MGMT hypermethylation: A prognostic foe, a predictive friend. DNA Repair 2007, 6, 1155–1160. [Google Scholar] [CrossRef] [PubMed]
- Hegi, M.E.; Diserens, A.C.; Gorlia, T.; Hamou, M.F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roger, S.; Hegi, M.E.; Mason, W.P.; Van Den Bent, M.J.; Taphoorn, M.J.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Barbara, F.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on viability in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Hedtke, V.; Bakovic, M. Choline transport for phospholipid synthesis: An emerging role of choline transporter-like protein 1. Exp. Biol. Med. (Maywood) 2019, 244, 655–662. [Google Scholar] [CrossRef] [Green Version]
- Inazu, M. Choline transporter-like proteins CTLs/SLC44 family as a novel molecular target for cancer therapy. Biopharm. Drug Dispos. 2014, 35, 431–449. [Google Scholar] [CrossRef]
- Michel, V.; Yuan, Z.; Ramsubir, S.; Bakovic, M. Choline transport for phospholipid synthesis. Exp. Biol. Med. (Maywood) 2006, 31, 490–504. [Google Scholar] [CrossRef]
- Giovannini, E.; Lazzeri, P.; Milano, A.; Gaeta, M.C.; Ciarmiello, A. Clinical applications of choline PET/CT in brain tumors. Curr. Pharm. Des. 2015, 21, 121–127. [Google Scholar] [CrossRef]
- Hara, T.; Kondo, T.; Kosaka, N. Use of 18F-choline and 11C-choline as contrast agents in positron emission tomography imaging-guided stereotactic biopsy sampling of gliomas. J. Neurosurg. 2003, 99, 474–479. [Google Scholar] [CrossRef]
- Nishiyama, R.; Nagashima, F.; Iwao, B.; Kawai, Y.; Inoue, K.; Midori, A.; Yamanaka, T.; Uchino, H.; Inazu, M. Identification and functional analysis of choline transporter in tongue cancer: A novel molecular target for tongue cancer therapy. J. Pharmacol. Sci. 2016, 131, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, F.; Nishiyama, R.; Iwao, B.; Kawai, Y.; Ishii, C.; Yamanaka, T.; Uchino, H.; Inazu, M. Molecular and Functional Characterization of Choline Transporter-Like Proteins in Esophageal Cancer Cells and Potential Therapeutic Targets. Biomol. Ther. (Seoul) 2018, 26, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Saiki, I.; Yara, M.; Yamanaka, T.; Uchino, H.; Inazu, M. Functional Expression of Choline Transporter-Like Protein 1 in LNCaP Prostate Cancer Cells: A Novel Molecular Target. Biomol. Ther. (Seoul) 2020, 28, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, C.; Inazu, M.; Saiki, I.; Yara, M.; Hara, N.; Yamanaka, T.; Uchino, H. Functional analysis of [methyl-(3)H]choline uptake in glioblastoma cells: Influence of anti-cancer and central nervous system drugs. Biochem. Pharmacol. 2014, 88, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Inazu, M.; Watanabe, S.; Nishijima, N.; Hirai, K.; Hase, A.; Shibata, K.; Gido, R.; Yamanaka, T. Development of new therapeutic drugs for glioma targeting choline transporter-like protein 1. Proc. 93rd Annu. Meet. Jpn. Pharmacol. Soc. 2020, 93, 3-P-348. [Google Scholar] [CrossRef] [Green Version]
- Kouji, H.; Inazu, M.; Yamada, T.; Tajima, H.; Aoki, T.; Matsumiya, T. Molecular and functional characterization of choline transporter in human colon carcinoma HT-29 cells. Arch. Biochem. Biophys. 2009, 483, 90–98. [Google Scholar] [CrossRef]
- Ogretmen, B.; Hannun, Y.A. Biologically active sphingolipids in cancer pathogenesis and treatment. Nat. Rev. Cancer 2004, 4, 604–616. [Google Scholar] [CrossRef]
- Goni, F.M.; Alonso, A. Sphingomyelinases: Enzymology and membrane activity. FEBS Lett. 2002, 531, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Kolesnick, R.N.; Kronke, M. Regulation of ceramide production and apoptosis. Annu. Rev. Physiol. 1998, 60, 643–665. [Google Scholar] [CrossRef]
- Nganga, R.; Oleinik, N.; Ogretmen, B. Mechanisms of Ceramide-Dependent Cancer Cell Death. Adv. Cancer Res. 2018, 140, 1–25. [Google Scholar] [CrossRef]
- Garg, H.; Suri, P.; Gupta, J.C.; Talwar, G.P.; Dubey, S. Survivin: A unique target for tumor therapy. Cancer Cell Int. 2016, 23, 49. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; Bhujwalla, Z.M.; Glunde, K. Targeting Phospholipid Metabolism in Cancer. Front. Oncol. 2016, 27, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glunde, K.; Ackerstaff, E.; Mori, N.; Jacobs, M.A.; Bhujwalla, Z.M. Choline phospholipid metabolism in cancer: Consequences for molecular pharmaceutical interventions. Mol. Pharm. 2006, 3, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Glunde, K.; Bhujwalla, Z.M.; Ronen, S.M. Choline metabolism in malignant transformation. Nat. Rev. Cancer 2011, 11, 835–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.H.; Liu, W.L.; Cao, H.; Wen, C.; Chen, L.; Jiang, G. O6-methylguanine DNA methyltransferase as a promising target for the treatment of temozolomide-resistant gliomas. Cell Death Dis. 2013, 4, e876. [Google Scholar] [CrossRef] [Green Version]
- Inazu, M.; Yamada, T.; Kubota, N.; Yamanaka, T. Functional expression of choline transporter-like protein 1 (CTL1) in small cell lung carcinoma cells: A target molecule for lung cancer therapy. Pharmacol. Res. 2013, 76, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Ogretmen, B. Sphingolipid metabolism in cancer signalling and therapy. Nat. Rev. Cancer 2018, 18, 33–50. [Google Scholar] [CrossRef]
- Morad, S.A.; Cabot, M.C. Ceramide-orchestrated signalling in cancer cells. Nat. Rev. Cancer 2018, 13, 51–65. [Google Scholar] [CrossRef]
- Lin, I.L.; Chou, H.L.; Lee, J.C.; Chen, F.W.; Fong, Y.; Chang, W.C.; Huang, H.W.; Wu, C.Y.; Chang, W.T.; Wang, H.D.; et al. The antiproliferative effect of C2-ceramide on lung cancer cells through apoptosis by inhibiting Akt and NFκB. Cancer Cell Int. 2014, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Ryland, L.; Yang, J.; Liao, A.; Aliaga, C.; Watts, R.; Tan, S.F.; Kaiser, J.; Shanmugavelandy, S.S.; Rogers, A.; et al. Targeting of survivin by nanoliposomal ceramide induces complete remission in a rat model of NK-LGL. leukemia. Blood 2010, 116, 4192–4201. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Ambrosini, G.; Chu, E.Y.; Plescia, J.; Tognin, S.; Marchisio, P.C.; Altieri, D.C. Control of apoptosis and mitotic spindle checkpoint by survivin. Nature 1998, 396, 580–584. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, C.; Song, Y.; Zhang, J.; Xu, J. Prognostic role of survivin in patients with glioma. Medicine 2018, 97, e0571. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.H.; Yang, L.; Chen, J.X.; Li, Q.R.; Zhu, L.R.; Xu, Q.F.; Huang, G.H.; Zhang, Z.X.; Xiang, Y.; Du, L.; et al. Bortezomib inhibits growth and sensitizes glioma to temozolomide (TMZ) via down-regulating the FOXM1-Survivin axis. Cancer Commun. 2019, 39, 81. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Zhuang, J.; He, L. Carboxamide derivatives induce apoptosis in the U251 glioma cell line. Oncol. Lett. 2019, 18, 1409–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Wang, Y.; Song, T.; Xin, T.; Zheng, Z.; Zhong, P.; Zhang, X. Silencing of survivin using YM155 inhibits invasion and suppresses proliferation in glioma cells. Cell Biochem. Biophys. 2015, 71, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Furihata, T.; Ito, R.; Kamiichi, A.; Saito, K.; Chiba, K. Establishment and characterization of a new conditionally immortalized human astrocyte cell line. J. Neurochem. 2016, 136, 92–105. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, S.; Nishijima, N.; Hirai, K.; Shibata, K.; Hase, A.; Yamanaka, T.; Inazu, M. Anticancer Activity of Amb4269951, a Choline Transporter-Like Protein 1 Inhibitor, in Human Glioma Cells. Pharmaceuticals 2020, 13, 104. https://0-doi-org.brum.beds.ac.uk/10.3390/ph13050104
Watanabe S, Nishijima N, Hirai K, Shibata K, Hase A, Yamanaka T, Inazu M. Anticancer Activity of Amb4269951, a Choline Transporter-Like Protein 1 Inhibitor, in Human Glioma Cells. Pharmaceuticals. 2020; 13(5):104. https://0-doi-org.brum.beds.ac.uk/10.3390/ph13050104
Chicago/Turabian StyleWatanabe, Saiichiro, Nozomi Nishijima, Kaho Hirai, Kaoru Shibata, Akane Hase, Tsuyoshi Yamanaka, and Masato Inazu. 2020. "Anticancer Activity of Amb4269951, a Choline Transporter-Like Protein 1 Inhibitor, in Human Glioma Cells" Pharmaceuticals 13, no. 5: 104. https://0-doi-org.brum.beds.ac.uk/10.3390/ph13050104