Calming the (Cytokine) Storm: Dimethyl Fumarate as a Therapeutic Candidate for COVID-19



1
Institute for Health and Sport, Victoria University, Melbourne, VIC 8001, Australia
2
Australian Institute for Musculoskeletal Science, St Albans, VIC 3021, Australia
*
Author to whom correspondence should be addressed.
Pharmaceuticals 2021, 14(1), 15; https://0-doi-org.brum.beds.ac.uk/10.3390/ph14010015
Submission received: 14 November 2020
/
Revised: 23 December 2020
/
Accepted: 23 December 2020
/
Published: 26 December 2020
(This article belongs to the Special Issue COVID-19 in Pharmaceuticals)
Abstract
:COVID-19 has rapidly spread worldwide and incidences of hospitalisation from respiratory distress are significant. While a vaccine is in the pipeline, there is urgency for therapeutic options to address the immune dysregulation, hyperinflammation and oxidative stress that can lead to death. Given the shared pathogenesis of severe cases of COVID-19 with aspects of multiple sclerosis and psoriasis, we propose dimethyl fumarate as a viable treatment option. Currently approved for multiple sclerosis and psoriasis, dimethyl fumarate is an immunomodulatory, anti-inflammatory and anti-oxidative drug that could be rapidly implemented into the clinic to calm the cytokine storm which drives severe COVID-19.
1. Introduction
The severe acute respiratory syndrome coronavirus type-2 (SARS-CoV-2) is responsible for the COVID-19 pandemic. Transmission, infection and mortality rates are significant (particularly compared to seasonal influenza) indicating the immediate need for a vaccine. Since it is currently unclear when an effective vaccine will be widely available and there are no standard-of-care treatment options for COVID-19, there is high unmet clinical need for therapeutics that can rapidly translate to improve patient care and reduce mortality rates.
Clinical presentation of COVID-19 varies but typical symptomology includes fever, cough and fatigue. In most cases hospitalisation is not required; however, for the elderly and those with co-morbidities (i.e., diabetes, cardiovascular disease, obesity, respiratory disease), the likelihood of hospitalisation (and mortality) increases significantly [1,2]. Severe cases usually present with acute respiratory distress syndrome (ARDS), the result of dysregulated host immune response to the virus [3]. The outcome of this immuno-dysregulation, which may be driven by underlying inflammation associated with age and co-morbidities, is a cytokine storm, i.e., elevated interferon (IFN), interleukins (IL), tumour necrosis factor-α (TNF-α), amongst others. The cytokine storm potentiates hyperinflammation, oxidative stress and haematological changes including lymphopenia, thrombocytopenia and macrophage activation syndrome. Collectively, these pathologies exacerbate the dysregulated host response and cause significant tissue injury to lung (and other) tissues resulting in respiratory (and often multi-organ) failure (Figure 1) [4]. Given the widespread organ/system assault in severe cases of COVID-19, there is clinical need for a therapeutic which addresses the multifactorial pathogenesis to induce systemic cytoprotection and re-establish host responsivity. Dimethyl fumarate (DMF), an approved drug with immunomodulatory, anti-oxidative and anti-inflammatory properties in all tissues, is one potential treatment that could be rapidly implemented into the clinic. Here, we provide perspectives on the potential re-purposement of DMF to treat the cytokine storm caused by severe COVID-19. We have searched the National Library of Medicine Pubmed® database from 20 August 2020 to 14 November 2020 using the key search terms “dimethyl fumarate”, “severe inflammation”, “cytokines”, “immunomodulation” and “COVID-19/SARS-CoV-2” and included in our discussion, those papers which match the mode of action of DMF with the symptomology of the cytokine storm induce by SARS-CoV-2.
2. Main
DMF is a methyl ester of fumaric acid (chemical formula C6H8O4) that is hydrolysed in the small intestine to the active metabolite monomethyl fumarate [5,6,7,8]. DMF is a potent activator of the nuclear factor erythroid 2-related factor 2 (Nrf2) pathway which modulates inflammation and oxidative stress by upregulating cellular defence mechanisms (i.e., cytoprotection through Phase II antioxidant expression, chiefly superoxide dismutase (SOD1), NAD(P)H quinone oxidoreductase-1 (NQO1) and heme oxygenase-1 (HO-1; Figure 2) [6,7,8,9]. In addition, DMF exerts some of its effects through Nrf2-independent mechanisms: (1) indirect inhibition of the inflammatory mediator, nuclear factor kappa B (NF-κB); and (2) hydroxycarboxylic acid receptor 2 (HCAR2) activation, which modulates immune cell (particularly neutrophil) infiltration, adhesion and chemotaxis, reduces pro-inflammatory cytokine production and inhibits NF-κB (Figure 2) [6,8,9,10,11,12]. DMF is clinically indicated for Multiple Sclerosis (MS) and psoriasis, diseases characterised by immune dysregulation, inflammation and oxidative stress [6]. Broadly, DMF treatment of these diseases: (1) Drives anti-inflammatory immune cell composition (Table 1; for a detailed review see [13]); (2) increases the ratio of anti- to pro-inflammatory cytokines; (3) induces the anti-oxidative and cytoprotective response through Nrf2; and (4) inhibits NF-κB to convert T helper (Th) cells from the Th1/17 subset to the anti-inflammatory Th2 subset [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31]. Collectively, these biological activities mitigate the severe pro-inflammatory and -oxidative mechanisms in both MS and psoriasis, which reduces disease progression to impart significant clinical impact.
The SARS-CoV-2 virus has been demonstrated to affect a significant number of immune cell populations including T cells, B cells, natural killer cells, monocytes, eosinophils and basophils [38,39,40,41,42]. These populations typically decrease in number (the severity of the disease dictates the magnitude of reduction), albeit pro-inflammatory phenotypes dominate the remaining immune cell population. Characteristic of the pro-inflammatory immune response is the increase in neutrophil-to-lymphocyte ratio (NLR). Neutrophilia and lymphopenia, which increase the NLR, are associated with severe viral infection and correlate with a poorer prognosis [43,44,45,46,47]. Moreover, in severe cases, the elevated neutrophil count is correlated with the formation of neutrophil extracellular traps (NETs). NETs are an important innate immunity defense mechanism as they trap and kill pathogens; however, their dysregulation induces oxidative stress (through reactive oxygen species (ROS) production), inflammation, damage, thrombosis and fibrosis to the surrounding tissues. It has been documented that neutrophils infiltrate the lungs [48,49,50] and induce elevated NET formation [48,49,50,51,52,53,54] in severe cases of COVID-19. DMF has been shown to modulate neutrophil counts [12,55] and NET formation [56,57]. Importantly, DMF reduces neutrophil adhesion, migration and infiltration [12,55,56,58] and neutrophil-induced ROS production [56] indicating that DMF can moderate the pro-inflammatory (and oxidative) effects of a dysregulated neutrophil response. In addition to DMF’s modulatory effects on various immune cell populations to shift from pro-inflammation to anti-inflammation, DMF also modifies an extensive cytokine profile [16,20,22,23,59], which is consistent with that observed in the cytokine storm characteristic of COVID-19 (i.e., granulocyte-colony stimulating factor (CSF), granulocyte-macrophage-CSF, IFN-γ, interferon-γ-inducible protein-10 (IP-10), IL-1β, IL-6, macrophage inflammatory protein (MIP)-1α, MIP-1β, monocyte chemoattractant protein-1 (MCP-1), TNF-α; see Table 2) [3,40,46,60,61,62]. Given the cytokine storm is strongly correlated with poorer prognosis [40,61,62], calming it is a logical approach. In this regard, the corticosteroid, dexamethasone, which shares some comparable immunosuppressive properties as DMF, has been used successfully in the clinical treatment of COVID-19 [63,64]. However, where corticosteroids elicit immunosuppression mainly through sequestration of CD4+ T-lymphocytes in the mononuclear phagocyte system and by inhibiting cytokine and lymphokine transcription (especially IL-1 and IL-6) [65], DMF modulates a more extensive cytokine profile as well as potent anti-oxidation activity. Of note, there is evidence that DMF induces lymphopenia in some MS patients [66] and, as such, diligent monitoring would be pertinent to ensure DMF does not exacerbate the lymphopenia documented in severe COVID-19 cases [43]. Despite this, DMF is generally well tolerated and is clinically approved indicating scope for rapid clinical translation. Importantly, the overall anti-inflammatory and anti-oxidative phenotype induced by DMF in MS and psoriasis patients would be beneficial for COVID-19 patients given the similar pathological mechanisms which advance disease severity and progression.
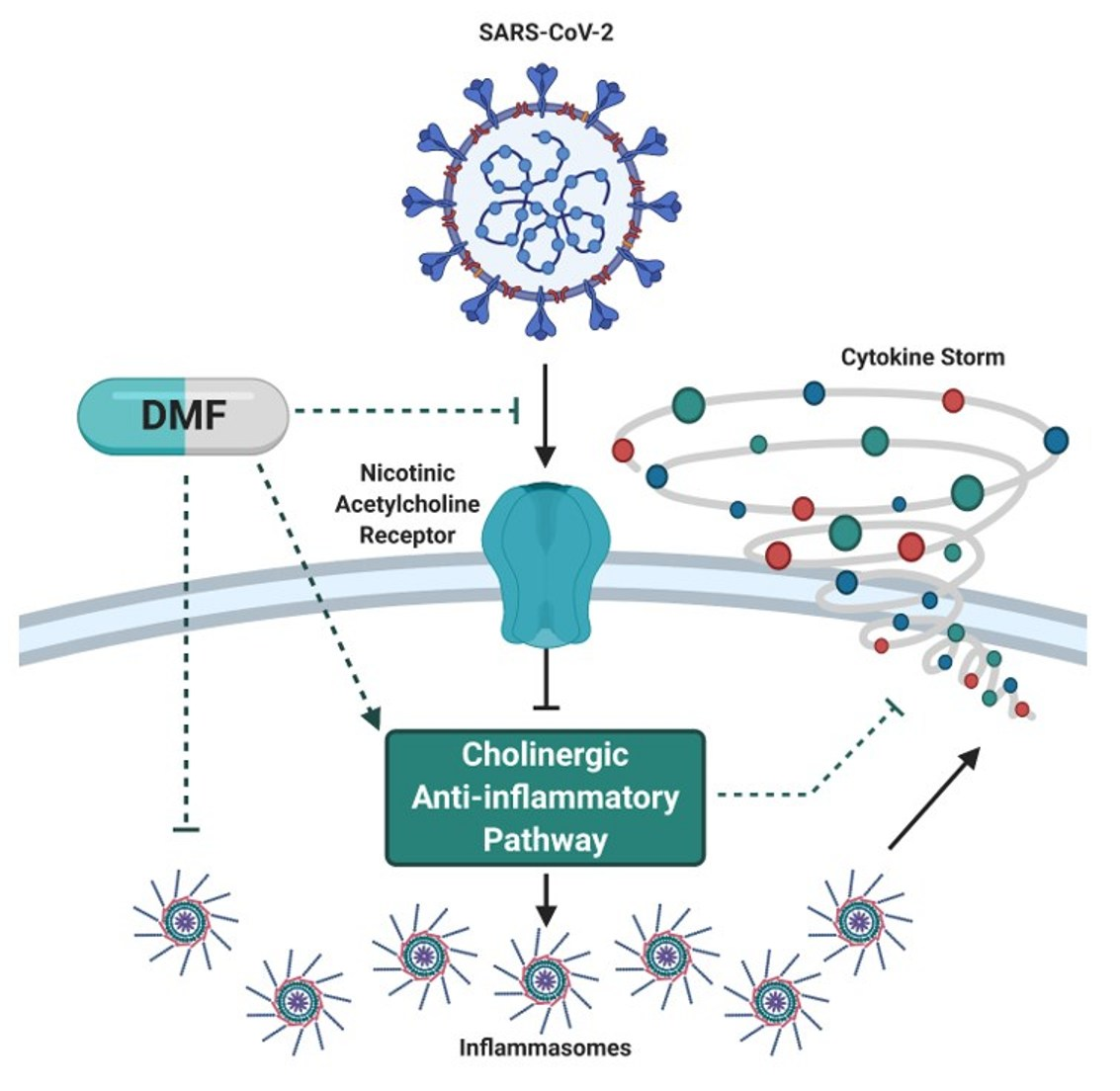
SARS-CoV-2 uses angiotensin converting enzyme II (ACE2) as its cellular entry receptor [67,68]. The binding of SARS-CoV-2 to ACE2 receptors is likely to reduce ACE2 receptor binding ability and attenuate downstream signalling from anti-inflammatory to pro-inflammatory pathways [69,70,71,72,73,74,75]. In the lungs, pro-inflammatory ACE2 signalling mediates immune cell infiltration, inflammation, injury and fibrosis [70,71,72,73,74,75]. While there is little research into the effect of DMF on ACE2 signalling, there is evidence that DMF can promote anti-inflammatory ACE2 signalling since it reduces inflammatory mediators (NF-κB-derived) and cytokine production in a murine model of acute lung injury [76]. In support of Nrf2 playing a role in ACE2 signalling, pharmacological Nrf2 inhibition decreases ACE2 mRNA [77]. Recent modelling also demonstrates that SARS-CoV-2 interacts with nicotinic acetylcholine receptors which may inhibit the cholinergic anti-inflammatory pathway and mediate COVID-19 pathology [78]. Consistent with this, the incidence of hospitalisation in smokers with COVID-19 is lower than predicted (albeit hospitalised smokers have poorer prognosis [79,80]) indicating that binding of nicotine to the receptor may competitively obstruct SARS-CoV-2 virulence [81]. DMF has been shown to facilitate cholinergic stimulation in MS patients [82] suggesting that, in the presence of SARS-CoV-2, DMF may competitively bind nicotinic acetylcholine receptors to reduce COVID-19 pathogenesis in the first instance. This is consistent with only several reported cases of COVID-19 in DMF-treated MS patients and with none of them having significant symptoms that required hospitalization [83].
There is growing evidence that the SARS-CoV-2 virus impedes the homeostatic response to restore redox balance. Nrf2, and its associated downstream antioxidant genes, are significantly reduced in lung biopsies of COVID-19 patients [3,84] suggesting Nrf2 suppression is a mechanism for SARS-CoV-2 replication. However, as with other conditions where Nrf2 induction is suppressed (e.g., aging-related sarcopenia), pharmacological rebalancing of Nrf2 responsiveness to noxious stimuli is possible. In vitro treatment of a SARS-CoV-2 infected Calu3 lung cancer cell line with the potent Nrf2 activator, DMF: (1) exerted an anti-viral effect through inhibition of SARS-CoV-2 replication; (2) reduced the pro-inflammatory cytokine profile; and (3) increased heme oxygenase-1 (HMOX1) gene expression [84]. HMOX1 gene upregulation (and HO-1 production) is associated with anti-viral activity against many viruses including influenza [85,86,87,88,89,90,91,92,93,94] and promotes the anti-inflammatory macrophage phenotype and anti-inflammatory IL-10 cytokine production [95,96]. Indeed, DMF is a well-known inducer of HO-1 [92,97,98,99,100,101,102,103,104,105,106]. Furthermore, previous literature indicates protective effects of DMF on respiratory pathologies including: (1) mitigation of the allergic asthma response [107,108]; (2) inhibition of airway smooth muscle cell proliferation (associated with airway remodeling) [109]; (3) reduction of lung inflammation [99,107,108,110,111] and cytokine production [111,112,113] and; (4) prevention of lung fibrosis in pulmonary arterial hypertension [99]. These findings emphasize the importance of the Nrf2 pathway in SARS-CoV-2 virulence and the potential therapeutic capacity of DMF for COVID-19 treatment, as recently suggested by [114].
3. Conclusions
Marked by immune dysregulation, hyperinflammation and oxidative stress, severe cases of COVID-19 may benefit from the immunomodulatory, anti-inflammatory and anti-oxidative properties of DMF. However, caution must be taken—the immunosuppressive effect of DMF may be counterproductive to mounting the host anti-viral immune response in the early stages of COVID-19 and, therefore, may expediate virulence. Thus, DMF may only be suitable for severe, progressed cases of COVID-19.
Author Contributions
Conceptualization, C.A.T. and E.R.; writing—original draft preparation, C.A.T. and E.R.; writing—review and editing, C.A.T. and E.R.; visualization, C.A.T. and E.R.; supervision, E.R.; project administration, E.R.; funding acquisition, E.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
E.R. is a scientific consultant to Santhera Pharmaceuticals. C.A.T. has no conflict of interest.
References
- Yang, J.; Zheng, Y.; Gou, X.; Pu, K.; Chen, Z.; Guo, Q.; Ji, R.; Wang, H.; Wang, Y.; Zhou, Y. Prevalence of comorbidities and its effects in patients infected with SARS-CoV-2: A systematic review and meta-analysis. Int. J. Infect. Dis. 2020, 94, 91–95. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: Summary of a report of 72 314 cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.-C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Pajares, M.; Benito, C.; Jiménez-Villegas, J.; Escoll, M.; Fernández-Ginés, R.; Yagüe, A.J.G.; Lastra, D.; Manda, G.; Rojo, A.I. Can activation of NRF2 be a strategy against COVID-19? Trends Pharmacol. Sci. 2020, 41, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Landeck, L.; Asadullah, K.; Amasuno, A.; Pau-Charles, I.; Mrowietz, U. Dimethyl fumarate (DMF) vs. monoethyl fumarate (MEF) salts for the treatment of plaque psoriasis: A review of clinical data. Arch. Dermatol. Res. 2018, 310, 475–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourakis, S.; Timpani, C.A.; de Haan, J.B.; Gueven, N.; Fischer, D.; Rybalka, E. Dimethyl Fumarate and Its Esters: A Drug with Broad Clinical Utility? Pharmaceuticals 2020, 13, 306. [Google Scholar] [CrossRef] [PubMed]
- Gill, A.J.; Kolson, D.L. Dimethyl fumarate modulation of immune and antioxidant responses: Application to HIV therapy. Crit. Rev. Immunol. 2013, 33, 307–359. [Google Scholar] [CrossRef] [Green Version]
- Brück, J.; Glocova, I.; Geisel, J.; Kellerer, C.; Röcken, M.; Ghoreschi, K. Dimethyl fumarate-induced IL-17lowIFN-γlowIL-4+ Th cells protect mice from severe encephalomyelitis. Eur. J. Immunol. 2018, 48, 1588–1591. [Google Scholar] [CrossRef] [Green Version]
- Kourakis, S.; Timpani, C.A.; de Haan, J.B.; Gueven, N.; Fischer, D.; Rybalka, E. Targeting Nrf2 for the treatment of Duchenne Muscular Dystrophy. Redox Biol. 2020, 38, 101803. [Google Scholar] [CrossRef]
- Al-Jaderi, Z.; Maghazachi, A.A. Utilization of Dimethyl Fumarate and Related Molecules for Treatment of Multiple Sclerosis, Cancer, and Other Diseases. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Von Glehn, F.; Dias-Carneiro, R.P.C.; Moraes, A.S.; Farias, A.S.; Silva, V.A.P.G.; Oliveira, F.T.M.; Silva, C.E.B.G.; de Carvalho, F.; Rahal, E.; Baecher-Allan, C.; et al. Dimethyl fumarate downregulates the immune response through the HCA2/GPR109A pathway: Implications for the treatment of multiple sclerosis. Mult. Scler. Relat. Disord. 2018, 23, 46–50. [Google Scholar] [CrossRef]
- Chen, H.; Assmann, J.C.; Krenz, A.; Rahman, M.; Grimm, M.; Karsten, C.M.; Köhl, J.; Offermanns, S.; Wettschureck, N.; Schwaninger, M. Hydroxycarboxylic acid receptor 2 mediates dimethyl fumarate’s protective effect in EAE. J. Clin. Investig. 2014, 124, 2188–2192. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.A.; Ogrodnik, M.A.; Plave, A.; Mao-Draayer, Y. Emerging Understanding of the Mechanism of Action for Dimethyl Fumarate in the Treatment of Multiple Sclerosis. Front. Neurol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Soin, D.; Ito, K.; Dhib-Jalbut, S. Insight into the mechanism of action of dimethyl fumarate in multiple sclerosis. J. Mol. Med. 2019, 97, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Gesser, B.; Rasmussen, M.K.; Iversen, L. Dimethyl Fumarate Targets MSK1, RSK1, 2 and IKKα/β Kinases and Regulates NF-κB/p65 Activation in Psoriasis: A Demonstration of the Effect on Peripheral Blood Mononuclear Cells, Drawn from Two Patients with Severe Psoriasis Before and After Treatment with Dimethyl Fumarate. Psoriasis Targets Ther. 2020, 10, 1. [Google Scholar]
- Gross, C.C.; Schulte-Mecklenbeck, A.; Klinsing, S.; Posevitz-Fejfár, A.; Wiendl, H.; Klotz, L. Dimethyl fumarate treatment alters circulating T helper cell subsets in multiple sclerosis. Neurol. Neuroimmunol. Neuroinflamm. 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Rezk, A.; Ghadiri, M.; Luessi, F.; Zipp, F.; Li, H.; Giacomini, P.S.; Antel, J.; Bar-Or, A. Dimethyl Fumarate Treatment Mediates an Anti-Inflammatory Shift in B Cell Subsets of Patients with Multiple Sclerosis. J. Immunol. 2017, 198, 691–698. [Google Scholar] [CrossRef]
- Montes Diaz, G.; Fraussen, J.; Van Wijmeersch, B.; Hupperts, R.; Somers, V. Dimethyl fumarate induces a persistent change in the composition of the innate and adaptive immune system in multiple sclerosis patients. Sci. Rep. 2018, 8, 8194. [Google Scholar] [CrossRef]
- Najjar, E.; Staun-Ram, E.; Volkowich, A.; Miller, A. Dimethyl fumarate promotes B cell-mediated anti-inflammatory cytokine profile in B and T cells, and inhibits immune cell migration in patients with MS. J. Neuroimmunol. 2020, 343, 577230. [Google Scholar] [CrossRef]
- Ockenfels, H.; Schultewolter, T.; Ockenfels, G.; Funk, R.; Goos, M. The antipsoriatic agent dimethylfumarate immunomodulates T-cell cytokine secretion and inhibits cytokines of the psoriatic cytokine network. Brit. J. Dermatol. 1998, 139, 390–395. [Google Scholar] [CrossRef]
- Schlöder, J.; Berges, C.; Luessi, F.; Jonuleit, H. Dimethyl Fumarate Therapy Significantly Improves the Responsiveness of T Cells in Multiple Sclerosis Patients for Immunoregulation by Regulatory T Cells. Int. J. Mol. Sci. 2017, 18, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.D.; Martin, K.A.; Calabresi, P.A.; Bhargava, P. Dimethyl fumarate alters B-cell memory and cytokine production in MS patients. Ann. Clin. Transl. Neurol. 2017, 4, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Stoof, T.J.; Flier, J.; Sampat, S.; Nieboer, C.; Tensen, C.P.; Boorsma, D.M. The antipsoriatic drug dimethylfumarate strongly suppresses chemokine production in human keratinocytes and peripheral blood mononuclear cells. Brit. J. Dermatol. 2001, 144, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Tahvili, S.; Zandieh, B.; Amirghofran, Z. The effect of dimethyl fumarate on gene expression and the level of cytokines related to different T helper cell subsets in peripheral blood mononuclear cells of patients with psoriasis. Int. J. Dermatol. 2015, 54, e254–e260. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, Q.; Mao, G.; Dowling, C.A.; Lundy, S.K.; Mao-Draayer, Y. Dimethyl Fumarate Selectively Reduces Memory T Cells and Shifts the Balance between Th1/Th17 and Th2 in Multiple Sclerosis Patients. J. Immunol. 2017, 198, 3069–3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longbrake, E.E.; Ramsbottom, M.J.; Cantoni, C.; Ghezzi, L.; Cross, A.H.; Piccio, L. Dimethyl fumarate selectively reduces memory T cells in multiple sclerosis patients. Mult. Scler. J. 2016, 22, 1061–1070. [Google Scholar] [CrossRef] [Green Version]
- Longbrake, E.E.; Cantoni, C.; Chahin, S.; Cignarella, F.; Cross, A.H.; Piccio, L. Dimethyl fumarate induces changes in B-and T-lymphocyte function independent of the effects on absolute lymphocyte count. Mult. Scler. J. 2018, 24, 728–738. [Google Scholar] [CrossRef]
- Michell-Robinson, M.A.; Moore, C.S.; Healy, L.M.; Osso, L.A.; Zorko, N.; Grouza, V.; Touil, H.; Poliquin-Lasnier, L.; Trudelle, A.M.; Giacomini, P.S. Effects of fumarates on circulating and CNS myeloid cells in multiple sclerosis. Ann. Clin. Transl. Neurol. 2016, 3, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Fleischer, V.; Friedrich, M.; Rezk, A.; Bühler, U.; Witsch, E.; Uphaus, T.; Bittner, S.; Groppa, S.; Tackenberg, B.; Bar-Or, A. Treatment response to dimethyl fumarate is characterized by disproportionate CD8+ T cell reduction in MS. Mult. Scler. J. 2018, 24, 632–641. [Google Scholar] [CrossRef]
- Lundy, S.K.; Wu, Q.; Wang, Q.; Dowling, C.A.; Taitano, S.H.; Mao, G.; Mao-Draayer, Y. Dimethyl fumarate treatment of relapsing-remitting multiple sclerosis influences B-cell subsets. Neurol. Neuroimmunol. Neuroinflamm. 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Topphoff, U.; Varrin-Doyer, M.; Pekarek, K.; Spencer, C.M.; Shetty, A.; Sagan, S.A.; Cree, B.A.; Sobel, R.A.; Wipke, B.T.; Steinman, L. Dimethyl fumarate treatment induces adaptive and innate immune modulation independent of Nrf2. Proc. Natl. Acad. Sci. USA 2016, 113, 4777–4782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghadiri, M.; Rezk, A.; Li, R.; Evans, A.; Luessi, F.; Zipp, F.; Giacomini, P.S.; Antel, J.; Bar-Or, A. Dimethyl fumarate–induced lymphopenia in MS due to differential T-cell subset apoptosis. Neurol. Neuroimmunol. Neuroinflamm. 2017, 4, e340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diebold, M.; Sievers, C.; Bantug, G.; Sanderson, N.; Kappos, L.; Kuhle, J.; Lindberg, R.L.P.; Derfuss, T. Dimethyl fumarate influences innate and adaptive immunity in multiple sclerosis. J. Autoimmun. 2018, 86, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Traub, J.; Traffehn, S.; Ochs, J.; Häusser-Kinzel, S.; Stephan, S.; Scannevin, R.; Brück, W.; Metz, I.; Weber, M.S. Dimethyl fumarate impairs differentiated B cells and fosters central nervous system integrity in treatment of multiple sclerosis. Brain Pathol. 2019, 29, 640–657. [Google Scholar] [CrossRef] [Green Version]
- Spencer, C.M.; Crabtree-Hartman, E.C.; Lehmann-Horn, K.; Cree, B.A.C.; Zamvil, S.S. Reduction of CD8+ T lymphocytes in multiple sclerosis patients treated with dimethyl fumarate. Neurol. Neuroimmunol. Neuroinflamm. 2015, 2, e76. [Google Scholar] [CrossRef] [Green Version]
- Staun-Ram, E.; Najjar, E.; Volkowich, A.; Miller, A. Dimethyl fumarate as a first- vs second-line therapy in MS. Focus B Cells 2018, 5, e508. [Google Scholar] [CrossRef] [Green Version]
- Holm Hansen, R.; Højsgaard Chow, H.; Sellebjerg, F.; Rode von Essen, M. Dimethyl fumarate therapy suppresses B cell responses and follicular helper T cells in relapsing-remitting multiple sclerosis. Mult. Scler. J. 2019, 25, 1289–1297. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Tan, M.; Liu, Y.; Zhou, R.; Deng, X.; Li, F.; Liang, K.; Shi, Y. Immunopathological characteristics of coronavirus disease 2019 cases in Guangzhou, China. Immunology 2020, 160, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, Y.; Xiang, P.; Pu, L.; Xiong, H.; Li, C.; Zhang, M.; Tan, J.; Xu, Y.; Song, R.; et al. Neutrophil-to-lymphocyte ratio predicts critical illness patients with 2019 coronavirus disease in the early stage. J. Transl. Med. 2020, 18, 206. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus–Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- Zhang, B.; Zhou, X.; Zhu, C.; Song, Y.; Feng, F.; Qiu, Y.; Feng, J.; Jia, Q.; Song, Q.; Zhu, B.; et al. Immune Phenotyping Based on the Neutrophil-to-Lymphocyte Ratio and IgG Level Predicts Disease Severity and Outcome for Patients With COVID-19. Front. Mol. Biosci. 2020, 7. [Google Scholar] [CrossRef]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef]
- Fu, J.; Kong, J.; Wang, W.; Wu, M.; Yao, L.; Wang, Z.; Jin, J.; Wu, D.; Yu, X. The clinical implication of dynamic neutrophil to lymphocyte ratio and D-dimer in COVID-19: A retrospective study in Suzhou China. Thromb. Res. 2020, 192, 3–8. [Google Scholar] [CrossRef]
- Barnes, B.J.; Adrover, J.M.; Baxter-Stoltzfus, A.; Borczuk, A.; Cools-Lartigue, J.; Crawford, J.M.; Daßler-Plenker, J.; Guerci, P.; Huynh, C.; Knight, J.S.; et al. Targeting potential drivers of COVID-19: Neutrophil extracellular traps. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Fox, S.E.; Akmatbekov, A.; Harbert, J.L.; Li, G.; Quincy Brown, J.; Vander Heide, R.S. Pulmonary and cardiac pathology in African American patients with COVID-19: An autopsy series from New Orleans. Lancet Respir. Med. 2020, 8, 681–686. [Google Scholar] [CrossRef]
- Wang, J.; Li, Q.; Yin, Y.; Zhang, Y.; Cao, Y.; Lin, X.; Huang, L.; Hoffmann, D.; Lu, M.; Qiu, Y. Excessive Neutrophils and Neutrophil Extracellular Traps in COVID-19. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Radermecker, C.; Detrembleur, N.; Guiot, J.; Cavalier, E.; Henket, M.; d’Emal, C.; Vanwinge, C.; Cataldo, D.; Oury, C.; Delvenne, P.; et al. Neutrophil extracellular traps infiltrate the lung airway, interstitial, and vascular compartments in severe COVID-19. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil extracellular traps in COVID-19. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, E.A.; He, X.-Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Skendros, P.; Mitsios, A.; Chrysanthopoulou, A.; Mastellos, D.C.; Metallidis, S.; Rafailidis, P.; Ntinopoulou, M.; Sertaridou, E.; Tsironidou, V.; Tsigalou, C.; et al. Complement and tissue factor–enriched neutrophil extracellular traps are key drivers in COVID-19 immunothrombosis. J. Clin. Investig. 2020, 130, 6151–6157. [Google Scholar] [CrossRef]
- Wannick, M.; Assmann, J.C.; Vielhauer, J.F.; Offermanns, S.; Zillikens, D.; Sadik, C.D.; Schwaninger, M. The Immunometabolomic Interface Receptor Hydroxycarboxylic Acid Receptor 2 Mediates the Therapeutic Effects of Dimethyl Fumarate in Autoantibody-Induced Skin Inflammation. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Müller, S.; Behnen, M.; Bieber, K.; Möller, S.; Hellberg, L.; Witte, M.; Hänsel, M.; Zillikens, D.; Solbach, W.; Laskay, T.; et al. Dimethylfumarate Impairs Neutrophil Functions. J. Investig. Dermatol. 2016, 136, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, J.H.O.; Schaekel, K.; Hartl, D.; Enk, A.H.; Hadaschik, E.N. Dimethyl fumarate modulates neutrophil extracellular trap formation in a glutathione- and superoxide-dependent manner. Br. J. Dermatol. 2018, 178, 207–214. [Google Scholar] [CrossRef]
- Giustina, A.D.; Bonfante, S.; Zarbato, G.F.; Danielski, L.G.; Mathias, K.; de Oliveira, A.N.; Garbossa, L.; Cardoso, T.; Fileti, M.E.; De Carli, R.J.; et al. Dimethyl Fumarate Modulates Oxidative Stress and Inflammation in Organs After Sepsis in Rats. Inflammation 2018, 41, 315–327. [Google Scholar] [CrossRef]
- Gillard, G.O.; Collette, B.; Anderson, J.; Chao, J.; Scannevin, R.H.; Huss, D.J.; Fontenot, J.D. DMF, but not other fumarates, inhibits NF-κB activity in vitro in an Nrf2-independent manner. J. Neuroimmunol. 2015, 283, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 cytokine storm; what we know so far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, C.; Huang, F.; Yang, Y.; Wang, F.; Yuan, J.; Zhang, Z.; Qin, Y.; Li, X.; Zhao, D.; et al. Elevated plasma levels of selective cytokines in COVID-19 patients reflect viral load and lung injury. Nat. Sci. Rev. 2020, 7, 1003–1011. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Shen, C.; Li, J.; Yuan, J.; Wei, J.; Huang, F.; Wang, F.; Li, G.; Li, Y.; Xing, L.; et al. Plasma IP-10 and MCP-3 levels are highly associated with disease severity and predict the progression of COVID-19. J. Allergy Clin. Immunol. 2020, 146, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Tomazini, B.M.; Maia, I.S.; Cavalcanti, A.B.; Berwanger, O.; Rosa, R.G.; Veiga, V.C.; Avezum, A.; Lopes, R.D.; Bueno, F.R.; Silva, M.; et al. Effect of Dexamethasone on Days Alive and Ventilator-Free in Patients With Moderate or Severe Acute Respiratory Distress Syndrome and COVID-19: The CoDEX Randomized Clinical Trial. JAMA 2020, 324, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; Elmahi, E.; et al. Dexamethasone in Hospitalized Patients with Covid-19—Preliminary Report. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Barshes, N.R.; Goodpastor, S.E.; Goss, J.A. Pharmacologic immunosuppression. Front. Biosci. 2004, 9, 411–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longbrake, E.E.; Naismith, R.T.; Parks, B.J.; Wu, G.F.; Cross, A.H. Dimethyl fumarate-associated lymphopenia: Risk factors and clinical significance. Mult. Scler. J. Exp. Transl. Clin. 2015, 1, 2055217315596994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, P.; Wang, J.; Feng, J.; Zhou, H.; Li, X.; Zhong, W.; Hao, P. Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission. Sci. China Life Sci. 2020, 63, 457–460. [Google Scholar] [CrossRef] [Green Version]
- Verdecchia, P.; Cavallini, C.; Spanevello, A.; Angeli, F. The pivotal link between ACE2 deficiency and SARS-CoV-2 infection. Eur. J. Intern. Med. 2020, 76, 14–20. [Google Scholar] [CrossRef]
- Li, Y.; Cao, Y.; Zeng, Z.; Liang, M.; Xue, Y.; Xi, C.; Zhou, M.; Jiang, W. Angiotensin-converting enzyme 2/angiotensin-(1–7)/Mas axis prevents lipopolysaccharide–induced apoptosis of pulmonary microvascular endothelial cells by inhibiting JNK/NF–κB pathways. Sci. Rep. 2015, 5, 8209. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Yang, Y.; Huang, Y.; Pan, C.; Liu, L.; Qiu, H. Angiotensin-(1-7) attenuates lung fibrosis by way of Mas receptor in acute lung injury. J. Surg. Res. 2013, 185, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Yu, C.-H.; Li, W.; Li, T.; Luo, W.; Huang, S.; Wu, P.-S.; Cai, S.-X.; Li, X. Angiotensin-converting enzyme 2/angiotensin-(1-7)/Mas axis protects against lung fibrosis by inhibiting the MAPK/NF-κB pathway. Am. J. Respir. Cell Mol. Biol. 2014, 50, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, C.P.; Wohlford-Lenane, C.; Yamaguchi, Y.; Prindle, T.; Fulton, W.B.; Wang, S.; McCray, P.B., Jr.; Chappell, M.; Hackam, D.J.; Jia, H. Attenuation of pulmonary ACE2 activity impairs inactivation of des-Arg9 bradykinin/BKB1R axis and facilitates LPS-induced neutrophil infiltration. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 314, L17–L31. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-N.; Yang, X.-H.; Nissen, D.H.; Chen, Y.-Y.; Wang, L.-J.; Wang, J.-H.; Gao, J.-L.; Zhang, L.-Y. Dysregulated renin-angiotensin system contributes to acute lung injury caused by hind-limb ischemia-reperfusion in mice. Shock 2013, 40, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Qiu, H.-B.; Yang, Y.; Wang, L.; Ding, H.-M.; Li, H.-P. Losartan, an antagonist of AT1 receptor for angiotensin II, attenuates lipopolysaccharide-induced acute lung injury in rat. Arch. Biochem. Biophys. 2009, 481, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Liu, Z. ACE2 exhibits protective effects against LPS-induced acute lung injury in mice by inhibiting the LPS-TLR4 pathway. Exp. Mol. Pathol. 2020, 113, 104350. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Ghosh, A.; Lo, C.-S.; Chenier, I.; Scholey, J.W.; Filep, J.G.; Ingelfinger, J.R.; Zhang, S.-L.; Chan, J.S.D. Nrf2 Deficiency Upregulates Intrarenal Angiotensin-Converting Enzyme-2 and Angiotensin 1-7 Receptor Expression and Attenuates Hypertension and Nephropathy in Diabetic Mice. Endocrinology 2017, 159, 836–852. [Google Scholar] [CrossRef]
- Oliveira, A.S.F.; Ibarra, A.A.; Bermudez, I.; Casalino, L.; Gaieb, Z.; Shoemark, D.K.; Gallagher, T.; Sessions, R.B.; Amaro, R.E.; Mulholland, A.J. Simulations support the interaction of the SARS-CoV-2 spike protein with nicotinic acetylcholine receptors and suggest subtype specificity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Reddy, R.K.; Charles, W.N.; Sklavounos, A.; Dutt, A.; Seed, P.T.; Khajuria, A. The effect of smoking on COVID-19 severity: A systematic review and meta-analysis. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Farsalinos, K.; Barbouni, A.; Niaura, R. Systematic review of the prevalence of current smoking among hospitalized COVID-19 patients in China: Could nicotine be a therapeutic option? Int. Emerg. Med. 2020, 15, 845–852. [Google Scholar] [CrossRef]
- Farsalinos, K.; Niaura, R.; Le Houezec, J.; Barbouni, A.; Tsatsakis, A.; Kouretas, D.; Vantarakis, A.; Poulas, K. Nicotine and SARS-CoV-2: COVID-19 may be a disease of the nicotinic cholinergic system. Toxicol. Rep. 2020, 7, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, C.G.; Landi, D.; Monteleone, F.; Mataluni, G.; Albanese, M.; Lauretti, B.; Rocchi, C.; Simonelli, I.; Boffa, L.; Buttari, F.; et al. Treatment with Dimethyl Fumarate Enhances Cholinergic Transmission in Multiple Sclerosis. CNS Drugs 2019, 33, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Mantero, V.; Abate, L.; Basilico, P.; Balgera, R.; Salmaggi, A.; Nourbakhsh, B.; Cordano, C. COVID-19 in dimethyl fumarate-treated patients with multiple sclerosis. J. Neurol. 2020, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Olagnier, D.; Farahani, E.; Thyrsted, J.; Blay-Cadanet, J.; Herengt, A.; Idorn, M.; Hait, A.; Hernaez, B.; Knudsen, A.; Iversen, M.B.; et al. SARS-CoV2-mediated suppression of NRF2-signaling reveals potent antiviral and anti-inflammatory activity of 4-octyl-itaconate and dimethyl fumarate. Nat. Commun. 2020, 11, 4938. [Google Scholar] [CrossRef]
- Espinoza, J.A.; González, P.A.; Kalergis, A.M. Modulation of Antiviral Immunity by Heme Oxygenase-1. Am. J. Pathol. 2017, 187, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Hashiba, T.; Suzuki, M.; Nagashima, Y.; Suzuki, S.; Inoue, S.; Tsuburai, T.; Matsuse, T.; Ishigatubo, Y. Adenovirus-mediated transfer of heme oxygenase-1 cDNA attenuates severe lung injury induced by the influenza virus in mice. Gene Ther. 2001, 8, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Cummins, N.W.; Weaver, E.A.; May, S.M.; Croatt, A.J.; Foreman, O.; Kennedy, R.B.; Poland, G.A.; Barry, M.A.; Nath, K.A.; Badley, A.D. Heme oxygenase-1 regulates the immune response to influenza virus infection and vaccination in aged mice. FASEB J. 2012, 26, 2911–2918. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Yasuda, H.; Sasaki, T.; Yamaya, M. Low-Dose Carbon Monoxide Inhibits Rhinovirus Replication in Human Alveolar and Airway Epithelial Cells. Tohoku J. Exp. Med. 2019, 247, 215–222. [Google Scholar] [CrossRef]
- Zhang, A.; Wan, B.; Jiang, D.; Wu, Y.; Ji, P.; Du, Y.; Zhang, G. The Cytoprotective Enzyme Heme Oxygenase-1 Suppresses Pseudorabies Virus Replication in vitro. Front. Microbiol. 2020, 11, 412. [Google Scholar] [CrossRef]
- Gutiérrez-Grobe, Y.; Vitek, L.; Tiribelli, C.; Kobashi-Margáin, R.A.; Uribe, M.; Méndez-Sánchez, N. Biliverdin and heme oxygenase antiviral activity against hepatitis C virus. Ann. Hepatol. 2016, 10, 105–107. [Google Scholar]
- Tseng, C.-K.; Lin, C.-K.; Wu, Y.-H.; Chen, Y.-H.; Chen, W.-C.; Young, K.-C.; Lee, J.-C. Human heme oxygenase 1 is a potential host cell factor against dengue virus replication. Sci. Rep. 2016, 6, 32176. [Google Scholar] [CrossRef] [PubMed]
- Cross, S.A.; Cook, D.R.; Chi, A.W.S.; Vance, P.J.; Kolson, L.L.; Wong, B.J.; Jordan-Sciutto, K.L.; Kolson, D.L. Dimethyl Fumarate, an Immune Modulator and Inducer of the Antioxidant Response, Suppresses HIV Replication and Macrophage-Mediated Neurotoxicity: A Novel Candidate for HIV Neuroprotection. J. Immunol. 2011, 187, 5015–5025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.; Cai, J.; Kostuk, E.W.; Rosenwasser, R.; Iacovitti, L. Fumarate modulates the immune/inflammatory response and rescues nerve cells and neurological function after stroke in rats. J. Neuroinflamm. 2016, 13, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, R.; Xiao, J.; Zhai, H.; Hao, J. Dimethyl fumarate attenuates experimental autoimmune neuritis through the nuclear factor erythroid-derived 2-related factor 2/hemoxygenase-1 pathway by altering the balance of M1/M2 macrophages. J. Neuroinflamm. 2016, 13, 97. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Nakamura, K.; Kageyama, S.; Lawal, A.O.; Gong, K.W.; Bhetraratana, M.; Fujii, T.; Sulaiman, D.; Hirao, H.; Bolisetty, S.; et al. Myeloid HO-1 modulates macrophage polarization and protects against ischemia-reperfusion injury. JCI Insight 2018, 3, e120596. [Google Scholar] [CrossRef]
- Drechsler, Y.; Dolganiuc, A.; Norkina, O.; Romics, L.; Li, W.; Kodys, K.; Bach, F.H.; Mandrekar, P.; Szabo, G. Heme Oxygenase-1 Mediates the Anti-Inflammatory Effects of Acute Alcohol on IL-10 Induction Involving p38 MAPK Activation in Monocytes. J. Immunol. 2006, 177, 2592–2600. [Google Scholar] [CrossRef]
- Pitarokoili, K.; Bachir, H.; Sgodzai, M.; Grüter, T.; Haupeltshofer, S.; Duscha, A.; Pedreiturria, X.; Motte, J.; Gold, R. Induction of Regulatory Properties in the Intestinal Immune System by Dimethyl Fumarate in Lewis Rat Experimental Autoimmune Neuritis. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Campolo, M.; Casili, G.; Lanza, M.; Filippone, A.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Multiple mechanisms of dimethyl fumarate in amyloid β-induced neurotoxicity in human neuronal cells. J. Cell. Mol. Med. 2018, 22, 1081–1094. [Google Scholar] [CrossRef]
- Grzegorzewska, A.P.; Seta, F.; Han, R.; Czajka, C.A.; Makino, K.; Stawski, L.; Isenberg, J.S.; Browning, J.L.; Trojanowska, M. Dimethyl Fumarate ameliorates pulmonary arterial hypertension and lung fibrosis by targeting multiple pathways. Sci. Rep. 2017, 7, 41605. [Google Scholar] [CrossRef] [Green Version]
- Robles, L.; Vaziri, N.D.; Li, S.; Masuda, Y.; Takasu, C.; Takasu, M.; Vo, K.; Farzaneh, S.H.; Stamos, M.J.; Ichii, H. Dimethyl Fumarate Protects Pancreatic Islet Cells and Non-Endocrine Tissue in L-Arginine-Induced Chronic Pancreatitis. PLoS ONE 2014, 9, e107111. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, R.S.; Abdel-Rahman, N. Dimethyl fumarate ameliorates acetaminophen-induced hepatic injury in mice dependent of Nrf-2/HO-1 pathway. Life Sci. 2019, 217, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Gopal, S.; Mikulskis, A.; Gold, R.; Fox, R.J.; Dawson, K.T.; Amaravadi, L. Evidence of activation of the Nrf2 pathway in multiple sclerosis patients treated with delayed-release dimethyl fumarate in the Phase 3 DEFINE and CONFIRM studies. Mult. Scler. J. 2017, 23, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.J.; Park, S.; Kim, J.-Y.; Kim, H.-J.; Jeoung, N.H.; Choi, Y.-K.; Go, Y.; Park, K.-G.; Lee, I.-K. Dimethylfumarate attenuates restenosis after acute vascular injury by cell-specific and Nrf2-dependent mechanisms. Redox Biol. 2014, 2, 855–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoreschi, K.; Brück, J.; Kellerer, C.; Deng, C.; Peng, H.; Rothfuss, O.; Hussain, R.Z.; Gocke, A.R.; Respa, A.; Glocova, I.; et al. Fumarates improve psoriasis and multiple sclerosis by inducing type II dendritic cells. J. Exp. Med. 2011, 208, 2291–2303. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.X.; Lisi, L.; Russo, C.D.; Polak, P.E.; Sharp, A.; Weinberg, G.; Kalinin, S.; Feinstein, D.L. The Anti-Inflammatory Effects of Dimethyl Fumarate in Astrocytes Involve Glutathione and Haem Oxygenase-1. ASN Neuro 2011, 3, AN20100033. [Google Scholar] [CrossRef]
- Zhao, G.; Liu, Y.; Fang, J.; Chen, Y.; Li, H.; Gao, K. Dimethyl fumarate inhibits the expression and function of hypoxia-inducible factor-1α (HIF-1α). Biochem. Biophys. Res. Commun. 2014, 448, 303–307. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Sandey, M.; Suryawanshi, A.; Cattley, R.C.; Mishra, A. Dimethyl fumarate abrogates dust mite-induced allergic asthma by altering dendritic cell function. Immun. Inflamm. Dis. 2019, 7, 201–213. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Sandey, M.; Suryawanshi, A.; Cattley, R.C.; Mishra, A. Dimethyl fumarate attenuates T helper type 2 (Th2)-mediated allergic airway inflammation by modulating dendritic cell function. J. Immunol. 2019, 202, 55.16. [Google Scholar]
- Seidel, P.; Goulet, S.; Hostettler, K.; Tamm, M.; Roth, M. DMF inhibits PDGF-BB induced airway smooth muscle cell proliferation through induction of heme-oxygenase-1. Respir. Res. 2010, 11, 145. [Google Scholar] [CrossRef] [Green Version]
- Seidel, P.; Hostettler, K.E.; Hughes, J.M.; Tamm, M.; Roth, M. Dimethylfumarate inhibits CXCL10 via haem oxygenase-1 in airway smooth muscle. Eur. Respir. J. 2013, 41, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Seidel, P.; Merfort, I.; Hughes, J.M.; Oliver, B.G.G.; Tamm, M.; Roth, M. Dimethylfumarate inhibits NF-κB function at multiple levels to limit airway smooth muscle cell cytokine secretion. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L326–L339. [Google Scholar] [CrossRef] [PubMed]
- Alrashdan, Y.A.; Alkhouri, H.; Chen, E.; Lalor, D.J.; Poniris, M.; Henness, S.; Brightling, C.E.; Burgess, J.K.; Armour, C.L.; Ammit, A.J.; et al. Asthmatic airway smooth muscle CXCL10 production: Mitogen-activated protein kinase JNK involvement. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L1118–L1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidel, P.; Roth, M.; Ge, Q.; Merfort, I.; S’ng, C.T.; Ammit, A.J. IκBα glutathionylation and reduced histone H3 phosphorylation inhibit eotaxin and RANTES. Eur. Respir. J. 2011, 38, 1444–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, S.M.; Jawad, M.J.; Ahjel, S.W.; Singh, R.B.; Singh, J.; Awad, S.M.; Hadi, N.R. The Nrf2 Activator (DMF) and Covid-19: Is there a Possible Role? Med. Arch. 2020, 74, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Robles, L.; Vaziri, N.D.; Li, S.; Takasu, C.; Masuda, Y.; Vo, K.; Farzaneh, S.H.; Stamos, M.J.; Ichii, H. Dimethyl fumarate ameliorates acute pancreatitis in rodent. Pancreas 2015, 44, 441–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerhardt, S.; König, V.; Doll, M.; Hailemariam-Jahn, T.; Hrgovic, I.; Zöller, N.; Kaufmann, R.; Kippenberger, S.; Meissner, M. Dimethylfumarate protects against TNF-α-induced secretion of inflammatory cytokines in human endothelial cells. J. Inflamm. 2015, 12, 49. [Google Scholar] [CrossRef] [Green Version]
- McGuire, V.A.; Diez, T.R.-Z.; Emmerich, C.H.; Strickson, S.; Ritorto, M.S.; Sutavani, R.V.; Weiβ, A.; Houslay, K.F.; Knebel, A.; Meakin, P.J. Dimethyl fumarate blocks pro-inflammatory cytokine production via inhibition of TLR induced M1 and K63 ubiquitin chain formation. Sci. Rep. 2016, 6, 31159. [Google Scholar] [CrossRef] [Green Version]
- Safavi, F.; Thome, R.; Li, Z.; Zhang, G.-X.; Rostami, A. Dimethyl fumarate suppresses granulocyte macrophage colony-stimulating factor–producing Th1 cells in CNS neuroinflammation. Neurol. Neuroimmunol. Neuroinflamm. 2020, 7, e729. [Google Scholar] [CrossRef]
- Sangineto, M.; Grabherr, F.; Adolph, T.E.; Grander, C.; Reider, S.; Jaschke, N.; Mayr, L.; Schwärzler, J.; Dallio, M.; Moschen, A.R.; et al. Dimethyl fumarate ameliorates hepatic inflammation in alcohol related liver disease. Liver Int. 2020, 40, 1610–1619. [Google Scholar] [CrossRef] [Green Version]
- Wallbrecht, K.; Drick, N.; Hund, A.-C.; Schön, M.P. Downregulation of endothelial adhesion molecules by dimethylfumarate, but not monomethylfumarate, and impairment of dynamic lymphocyte-endothelial cell interactions. Exp. Dermatol. 2011, 20, 980–985. [Google Scholar] [CrossRef]
- Albrecht, P.; Bouchachia, I.; Goebels, N.; Henke, N.; Hofstetter, H.H.; Issberner, A.; Kovacs, Z.; Lewerenz, J.; Lisak, D.; Maher, P.; et al. Effects of dimethyl fumarate on neuroprotection and immunomodulation. J. Neuroinflamm. 2012, 9, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Ma, S.; Gong, H.; Liu, S.; Lei, L.; Hu, B.; Xu, Y.; Liu, H.; Wu, D. Inhibition of Acute Graft-versus-Host Disease with Retention of Graft-versus-Tumor Effects by Dimethyl Fumarate. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, J.C.U.; Listopad, J.J.; Rentzsch, C.U.; Igney, F.H.; von Bonin, A.; Hennekes, H.H.; Asadullah, K.; Docke, W.-D.F. Dimethylfumarate Induces Immunosuppression via Glutathione Depletion and Subsequent Induction of Heme Oxygenase 1. J. Investig. Dermatol. 2007, 127, 835–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, H.; Guerau-de-Arellano, M.; Mehta, V.B.; Yang, Y.; Huss, D.J.; Papenfuss, T.L.; Lovett-Racke, A.E.; Racke, M.K. Dimethyl Fumarate Inhibits Dendritic Cell Maturation via Nuclear Factor κB (NF-κB) and Extracellular Signal-regulated Kinase 1 and 2 (ERK1/2) and Mitogen Stress-activated Kinase 1 (MSK1) Signaling. J. Biol. Chem. 2012, 287, 28017–28026. [Google Scholar] [CrossRef] [Green Version]
- Müller, S.; Smatlik, N.; Burian, M.; Ghoreschi, K.; Röcken, M.; Yazdi, A.S. Differential induction of ATF3 and HO-1 in myeloid cells and keratinocytes via Dimethylfumarate or Cyclosporine A. J. Dermatol. Sci. 2017, 87, 246–251. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhou, W.; Zhang, X.; Lu, P.; Du, Q.; Tao, L.; Ding, Y.; Wang, Y.; Hu, R. Dimethyl fumarate ameliorates dextran sulfate sodium-induced murine experimental colitis by activating Nrf2 and suppressing NLRP3 inflammasome activation. Biochem. Pharmacol. 2016, 112, 37–49. [Google Scholar] [CrossRef]
- Singh, N.; Saha, L.; Kumari, P.; Singh, J.; Bhatia, A.; Banerjee, D.; Chakrabarti, A. Effect of dimethyl fumarate on neuroinflammation and apoptosis in pentylenetetrazol kindling model in rats. Brain Res. Bull. 2019, 144, 233–245. [Google Scholar] [CrossRef]
- Wierinckx, A.; Brevé, J.; Mercier, D.; Schultzberg, M.; Drukarch, B.; Van Dam, A.-M. Detoxication enzyme inducers modify cytokine production in rat mixed glial cells. J. Neuroimmunol. 2005, 166, 132–143. [Google Scholar] [CrossRef]
- Wilms, H.; Sievers, J.; Rickert, U.; Rostami-Yazdi, M.; Mrowietz, U.; Lucius, R. Dimethylfumarate inhibits microglial and astrocytic inflammation by suppressing the synthesis of nitric oxide, IL-1β, TNF-α and IL-6 in an in-vitro model of brain inflammation. J. Neuroinflamm. 2010, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Belcher, J.D.; Chen, C.; Nguyen, K.; Zhang, P.; Abdulla, F.; Nguyen, P.; Killeen, T.; Xu, P.; O’Sullivan, G.; Nath, K.A.; et al. Control of Oxidative Stress and Inflammation in Sickle Cell Disease with the Nrf2 Activator Dimethyl Fumarate. Antioxid. Redox Signal. 2017, 26, 748–762. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, G.; Zhang, J.; Ting, S.-M.; Gonzales, N.; Aronowski, J. Dimethyl Fumarate Protects Brain from Damage Produced by Intracerebral Hemorrhage by Mechanism Involving Nrf2. Stroke 2015, 46, 1923–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blewett, M.M.; Xie, J.; Zaro, B.W.; Backus, K.M.; Altman, A.; Teijaro, J.R.; Cravatt, B.F. Chemical proteomic map of dimethyl fumarate–sensitive cysteines in primary human T cells. Sci. Signal. 2016, 9, rs10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, N.Z.M.; Chiarotto, G.B.; Bernardes, D.; Kempe, P.R.G.; Oliveira, A.L.R. Neuroprotection by dimethyl fumarate following ventral root crush in C57BL/6J mice. Brain Res. Bull. 2020, 164, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Galloway, D.A.; Williams, J.B.; Moore, C.S. Effects of fumarates on inflammatory human astrocyte responses and oligodendrocyte differentiation. Ann. Clin. Transl. Neurol. 2017, 4, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, S.; Pace, B.; Gupta, D.; Sturtevant, S.; Li, B.; Makala, L.; Brittain, J.; Moore, N.; Vieira, B.F.; Thullen, T.; et al. Dimethyl fumarate increases fetal hemoglobin, provides heme detoxification, and corrects anemia in sickle cell disease. JCI Insight 2017, 2, e96409. [Google Scholar] [CrossRef] [Green Version]
- Ragab, D.; Abdallah, D.M.; El-Abhar, H.S. The dual reno-and neuro-protective effects of dimethyl fumarate against uremic encephalopathy in a renal ischemia/reperfusion model. Pharmacol. Rep. 2020, 72, 969–983. [Google Scholar] [CrossRef]
- Takasu, C.; Vaziri, N.D.; Li, S.; Robles, L.; Vo, K.; Takasu, M.; Pham, C.; Farzaneh, S.H.; Shimada, M.; Stamos, M.J.; et al. Treatment with dimethyl fumarate ameliorates liver ischemia/reperfusion injury. World J. Gastroenterol. 2017, 23, 4508–4516. [Google Scholar] [CrossRef]
- Seidel, P.; Merfort, I. Inhibition of NF-κB and AP-1 by dimethylfumarate correlates with down-regulated IL-6 secretion and proliferation in human lung fibroblasts. Swiss Med. Wkly. 2010, 140. [Google Scholar] [CrossRef]
- Zarbato, G.F.; de Souza Goldim, M.P.; Giustina, A.D.; Danielski, L.G.; Mathias, K.; Florentino, D.; de Oliveira Junior, A.N.; da Rosa, N.; Laurentino, A.O.; Trombetta, T.; et al. Dimethyl Fumarate Limits Neuroinflammation and Oxidative Stress and Improves Cognitive Impairment After Polymicrobial Sepsis. Neurotox. Res. 2018, 34, 418–430. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, G.; Wang, Y.; Lin, X.; Zhou, J.; Wang, Z.; Suo, N. Dimethyl fumarate protects nucleus pulposus cells from inflammation and oxidative stress and delays the intervertebral disc degeneration. Exp. Ther. Med. 2020, 20, 269. [Google Scholar] [CrossRef]
- Förster, A.; Preussner, L.M.; Seeger, J.M.; Rabenhorst, A.; Kashkar, H.; Mrowietz, U.; Hartmann, K. Dimethylfumarate induces apoptosis in human mast cells. Exp. Dermatol. 2013, 22, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Li, H.; Sheehy, A.; Cullen, P.; Allaire, N.; Scannevin, R.H. Dimethyl fumarate alters microglia phenotype and protects neurons against proinflammatory toxic microenvironments. J. Neuroimmunol. 2016, 299, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Litjens, N.H.R.; Rademaker, M.; Ravensbergen, B.; Rea, D.; van der Plas, M.J.A.; Thio, B.; Walding, A.; van Dissel, J.T.; Nibbering, P.H. Monomethylfumarate affects polarization of monocyte-derived dendritic cells resulting in down-regulated Th1 lymphocyte responses. Eur. J. Immunol. 2004, 34, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Lückel, C.; Picard, F.; Raifer, H.; Campos Carrascosa, L.; Guralnik, A.; Zhang, Y.; Klein, M.; Bittner, S.; Steffen, F.; Moos, S.; et al. IL-17+ CD8+ T cell suppression by dimethyl fumarate associates with clinical response in multiple sclerosis. Nat. Commun. 2019, 10, 5722. [Google Scholar] [CrossRef] [PubMed]
- Selman, M.; Ou, P.; Rousso, C.; Bergeron, A.; Krishnan, R.; Pikor, L.; Chen, A.; Keller, B.A.; Ilkow, C.; Bell, J.C.; et al. Dimethyl fumarate potentiates oncolytic virotherapy through NF-κB inhibition. Sci. Transl. Med. 2018, 10, eaao1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
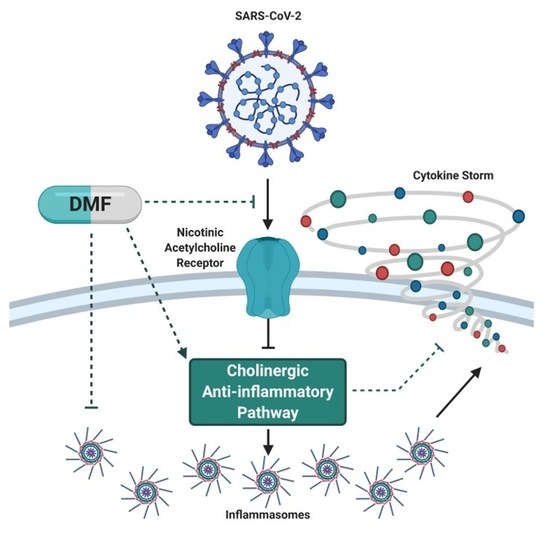
Figure 1.
Simplified pathological pathway to lung (and systemic) injury in severe cases of COVID-19. Infection with the SARS-CoV-2 virus can lead to a dysregulated immune response in which pro-inflammatory cells dominate the immune cell population. These pro-inflammatory cells intensify cytokine production and release resulting in hyperinflammation. This hyperinflammatory state promotes lung (and systemic) pathology, which correlates with poorer prognosis. It is well documented that dimethyl fumarate (DMF) can modulate immune cell populations to shift the ratio of anti-inflammatory to pro-inflammatory cytokine production and release, which in turn reduces hyperinflammation and subsequent tissue injury.
Figure 1.
Simplified pathological pathway to lung (and systemic) injury in severe cases of COVID-19. Infection with the SARS-CoV-2 virus can lead to a dysregulated immune response in which pro-inflammatory cells dominate the immune cell population. These pro-inflammatory cells intensify cytokine production and release resulting in hyperinflammation. This hyperinflammatory state promotes lung (and systemic) pathology, which correlates with poorer prognosis. It is well documented that dimethyl fumarate (DMF) can modulate immune cell populations to shift the ratio of anti-inflammatory to pro-inflammatory cytokine production and release, which in turn reduces hyperinflammation and subsequent tissue injury.

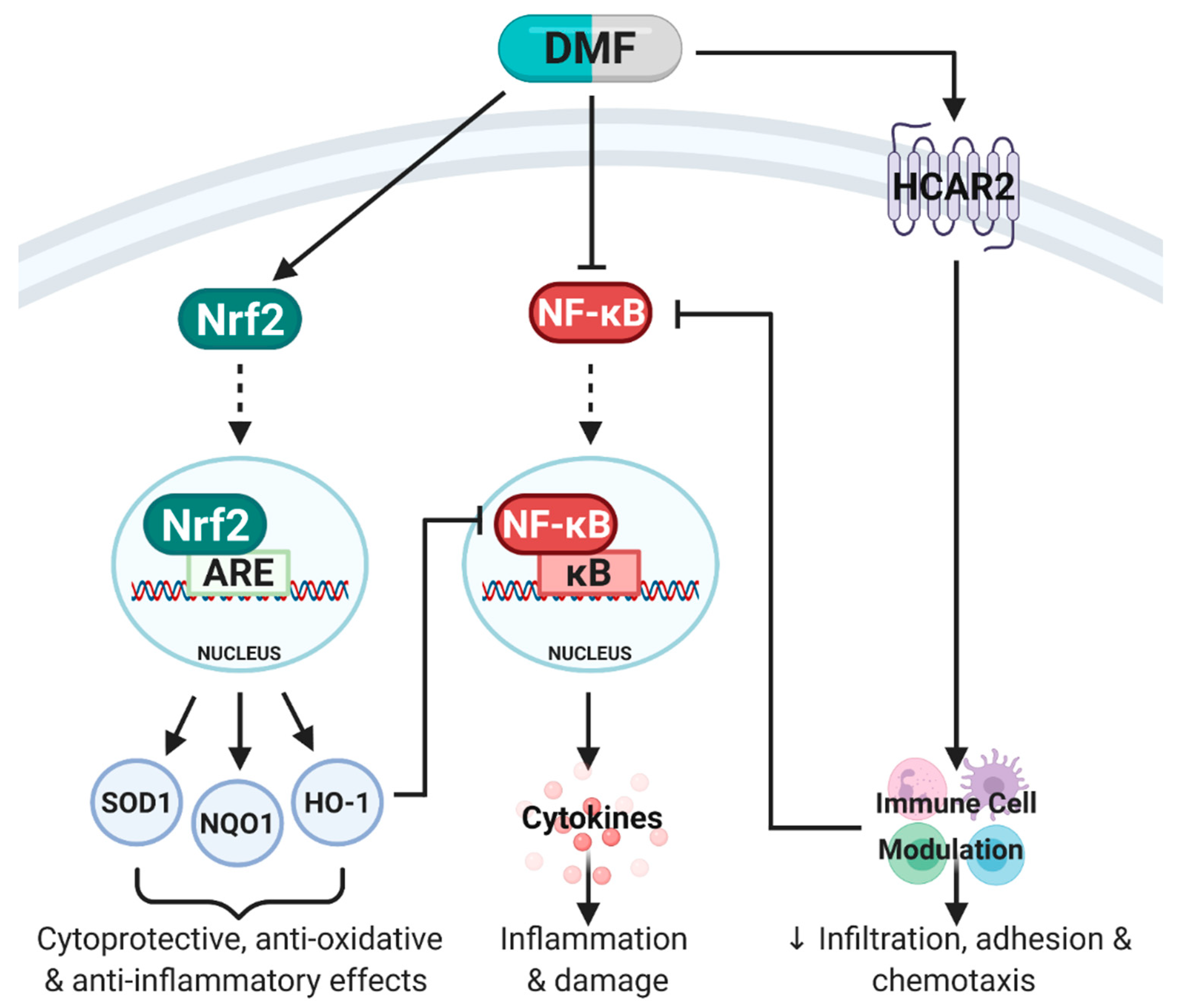
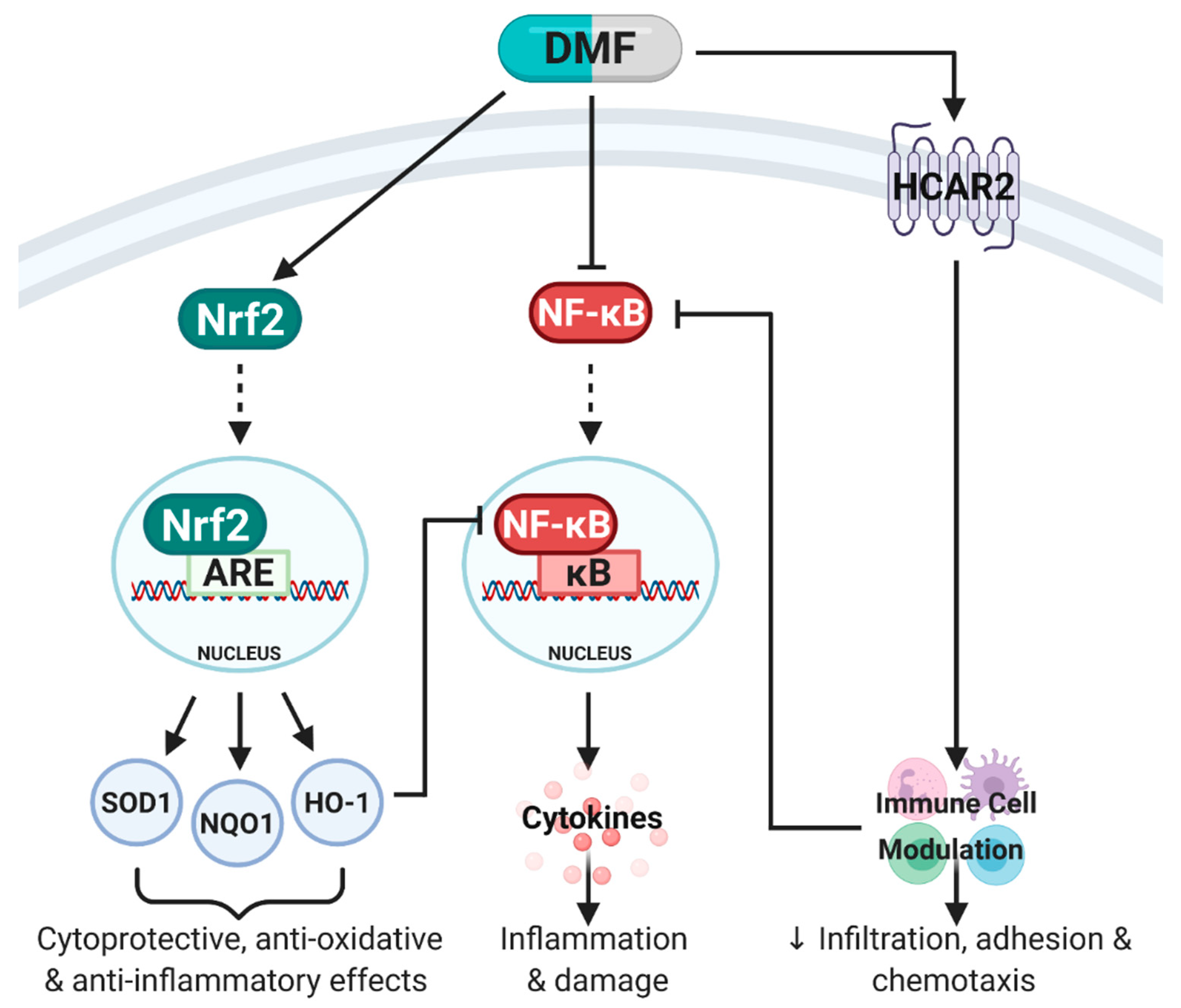
Figure 2.
Simplified schematic of pathways activated by dimethyl fumarate (DMF). DMF is known to mediate its anti-inflammatory, anti-oxidative and immunomodulatory effects primarily through three molecular pathways: (1) Activation of nuclear factor erythroid 2-related factor 2 (Nrf2) which binds to the antioxidant response element (ARE) in the nucleus to stimulate transcription of Phase II enzymes including superoxide dismutase (SOD1), NAD(P)H quinone oxidoreductase-1 (NQO1) and heme oxygenase-1 (HO-1). Together, SOD1, NQO1 and HO-1 mediate cytoprotective, anti-oxidative and anti-inflammatory effects. (2) DMF can directly inhibit nuclear factor kappa B (NF-κB), which prevents the translocation of NF-κB into the nucleus, binding to the κB site, release of pro-inflammatory cytokines and subsequent inflammation and damage. DMF can also indirectly inhibit NF-κB through HO-1 expression and immune cell modulation through hydroxycarboxylic acid receptor 2 (HCAR2) activation. (3) DMF’s immunomodulatory effects are predominantly mediated through HCAR2 activation which modulates immune cell populations (pro-inflammatory to anti-inflammatory shift) and inhibits NF-κB. Adapted from [7]. Created with BioRender.com.
Figure 2.
Simplified schematic of pathways activated by dimethyl fumarate (DMF). DMF is known to mediate its anti-inflammatory, anti-oxidative and immunomodulatory effects primarily through three molecular pathways: (1) Activation of nuclear factor erythroid 2-related factor 2 (Nrf2) which binds to the antioxidant response element (ARE) in the nucleus to stimulate transcription of Phase II enzymes including superoxide dismutase (SOD1), NAD(P)H quinone oxidoreductase-1 (NQO1) and heme oxygenase-1 (HO-1). Together, SOD1, NQO1 and HO-1 mediate cytoprotective, anti-oxidative and anti-inflammatory effects. (2) DMF can directly inhibit nuclear factor kappa B (NF-κB), which prevents the translocation of NF-κB into the nucleus, binding to the κB site, release of pro-inflammatory cytokines and subsequent inflammation and damage. DMF can also indirectly inhibit NF-κB through HO-1 expression and immune cell modulation through hydroxycarboxylic acid receptor 2 (HCAR2) activation. (3) DMF’s immunomodulatory effects are predominantly mediated through HCAR2 activation which modulates immune cell populations (pro-inflammatory to anti-inflammatory shift) and inhibits NF-κB. Adapted from [7]. Created with BioRender.com.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Overview of the effect of dimethyl fumarate on T and B cells in relapsing–remitting multiple sclerosis patients.
Table 1.
Overview of the effect of dimethyl fumarate on T and B cells in relapsing–remitting multiple sclerosis patients.
| Patient Number | Age Range (years) | Length of DMF Treatment | Additional Medication during DMF Treatment | Effect on T & B Cells | Ref |
|---|---|---|---|---|---|
| 15 (7F/8M) | 24–54 (median 40.7) | 6 m | - | T cells: ↓ Th1 & Th17 cells, ↑ CD4 and CD8 naïve cells, ↓ CD4 and CD8 memory cells, ↓ CD8 cells | [16] |
| 13 (11F/2M) | 20–60 (median 41) | Not stated | - | B cells: ↓ B cell number, ↓ memory B cells, ↑ naïve B cells, ↓ pro-inflammatory B cells (GM-CSF+, IL-6+, TNF-α+), ↓ pro-inflammatory co-stimulatory molecules (CD80+) | [17] |
| 20 (16F/4M) | 43 ± 8 | 4–6 m | - | T cells: ↑ Th2/Th1Th17 ratio, ↓ memory T cells, ↑ naïve T cells, ↓ CD4 and CD8 cells, ↓ pro-inflammatory T cells (IFN-γ+), ↑ anti-inflammatory T cells (IL-4+) B cells: ↓ B cell number | [25] |
| 18 (14F/4M) | 43 ± 9 | 18–26 m | - | T cells: ↓ Th1 & Th17 cells, ↑ Th2 cells, ↑Th2/Th1Th17 ratio, ↓ CD4 and CD8 cells, ↓ memory T cells, ↑ naïve T cells, ↓ pro-inflammatory T cells (IFN-γ+, IL-17+), ↑ anti-inflammatory T cells (IL-4+) B cells: ↓ B cell number | |
| 18 (13F/5M) | 43.9 ± 10.8 | 6 m | - | B cells: ↓ memory B cells, ↑ naïve B cells, ↓ pro-inflammatory B cells (GM-CSF+, IL-6+, TNF-α+) | [22] |
| 24 (21F/3M) | 24–63 (median 44.6) | ≥6 m | - | T cells: ↓ Th1 cells, ↓ CD4 and CD8 memory cells, ↑ CD4 and CD8 naïve cells | [26] |
| 43 (31F/12M) | 46±11 | 15 ± 9 m | - | T cells: ↓ CD8 memory cells, ↑ CD8 naïve cells, ↓ pro-inflammatory T cells (GM-CSF+, IFN-γ+, TNF-α+), B cells: ↓ pro-inflammatory co-stimulatory molecules (CD80+) | [27] |
| 13 (8F/5M) | Female: 31–58 (median 46.5) Male:33–57 (median 35) | 4–6 m | One patient tapered off steroids for first 6 weeks of DMF treatment | B cells: ↓ B cell number, ↓ memory B cells | [30] |
| 13 (11F/2M) | 20–60 (median 41) | 0–12 m | - | T cells: ↓ CD4 and CD8 cells, ↓ memory T cells, ↑ naïve T cells, ↓ pro-inflammatory T cells (IFN-γ+) | [32] |
| 20 (11F/9M) | 26–60 (median 41) | 0–12 m | - | T cells: ↓ T cell number, ↓ CD4 and CD8 cells, ↑ anti-inflammatory Treg cells, ↓ memory T cells, ↑ naïve T cells | [33] |
| 25 (48% F/52% M) | 35.4 ± 11.1 | At least 3 m | - | T cells: ↓ CD8 cells B cells: ↓ memory B cells, ↓ pro-inflammatory B cells (IL-6+, TNF-α+), ↓ pro-inflammatory co-stimulatory molecules (CD 40+, CD69+, CD80+, CD86+) | [34] |
| 35 (71.4% F/28.6% M) | 21–67 (mean 46.1) | 0–12 m | - | T cells: ↓ T cell number, ↓ CD4 and CD8 cells B cells: ↓ B cell number | [35] |
| 51 (35F/16M) | 34.8 ± 10.8 | 6 m | Methylprednisone-treated patient samples collected 4 weeks after last administration | T cells: ↓ T cell number, ↓ CD4 and CD8 cells B cells: ↓ B cell number | [29] |
| 43 (28F/15M) | 38 ± 2 | 15 w | - | T cells: ↑ transitional T cells B cells: ↓ memory B cells, ↑ naïve B cells, ↑ anti-inflammatory B cells (IL-4+, IL-10+, TGF-β+), ↓ pro-inflammatory co-stimulatory molecules (CD69+, CD80+, CD86+) | [36] |
| 21 | 25–50 (median 37) | 12 m | - | T cells: ↓ T cell number, ↑ transitional T cells B cells: ↓ B cell number, ↓ memory B cells, ↑ naïve B cells, | [37] |
Abbreviations: DMF: Dimethyl fumarate; GM-CSF: Granulocyte-macrophage-colony stimulating factor; IFN-γ: Interferon-γ; IL: Interleukin; Th: T helper cells; Treg: T regulatory cells; TGF-β: Transforming growth factor-β; TNF-α: Tumour necrosis factor-α.
Table 2.
Overview of the effect of dimethyl fumarate on cytokine production.
| Elevated Cytokines in COVID-19 Patients | Effect of DMF | Model/Disease |
|---|---|---|
| G-CSF [40,61,62] | ↓ | Murine splenocytes [59], Human primary ASMCs [110] |
| GM-CSF [40,61,62] | ↓ | Human RRMS PBMCs [16,17,18,22], Human Psoriatic PBMCs [24], Murine splenocytes [59,115], Human UVECs [116], Murine BMDMs [117], Murine EAE [118] |
| Gro-1α [62] | ↓ | Human keratinocytes & PBMCs [23], Murine hepatic injury & Kupffer cells [119], Human UVECs [120] |
| IFN-γ [40,61,62] | ↓ | Murine EAE [8,104,118], Human RRMS PBMCs [16,18,19,21,25], Human psoriatic keratinocytes [20], Human psoriatic PBMCs [24], Murine ischaemic stroke model [93], Murine EAN & macrophage cell line [94], Human psoriatic T cells [104], Murine splenocytes [115,121], Murine BMDCs & allogeneic splenic T cell co-culture [122], Human PBMCs [123], Murine BMDCs [124] |
| IL-1α [61,62] | ↓ | Murine splenocytes [115], Primary human keratinocytes & PBMCs [125] |
| IL-1β [40,61,62] | ↓ | Murine splenocytes [59], Murine ischaemic stroke model [93], Murine hepatic injury & Kupffer cells [119], Primary human keratinocytes & PBMCs [125], Murine colitis model [126], Murine epilepsy model [127], Primary murine microglial & astroglial co-cultures [128,129], Murine SCD model [130], Murine intracerebral hemorrhage models [131] |
| IL-2 [40,61,62] | ↓ | Murine EAE [8], Murine splenocytes [121], Murine BMDCs & allogeneic splenic T cell co-culture [122], Human PBMCs [123], Primary human & murine T cells [132] |
| IL-4 [40,61] | ↑ | Murine EAE [8,104], Human PBMCs [24], Human RRMS PBMCs [25], Murine EAN & macrophage cell line [94], Human psoriatic T cells [104], Murine spinal cord damage model [133] |
| IL-6 [3,40,61,62] | ↓ | Human RRMS PBMCs [17,21,22], Human psoriatic keratinocytes [20], Murine splenocytes [59,115,121], Murine EAN & macrophage cell line [94], Primary human asthmatic ASMCs [111], Human UVECs [116,120], Murine BMDMs [117], Murine BMDCs & allogeneic splenic T cell co-culture [122], Human PBMCs [123], Murine BMDCs [124], Primary human keratinocytes & PBMCs [125], Murine colitis model [126], Murine epilepsy model [127], Primary murine microglial & astroglial co-cultures [128,129], Primary human & murine astrocyte cultures [134], Murine & primate SCD models [130,135], Murine renal/liver I/R injury model [136,137], Primary human lung fibroblasts [138], Murine experimental sepsis [139], Murine IDD model [140] |
| IL-8 [3,40,61] | ↓ | Human keratinocytes & PBMCs [23], Murine osteoblastic cells [106], Human UVECs [120], Murine IDD model [140], Human mast cell line & primary CBDMCs [141] |
| IL-10 [40,61,62] | ↑ | Human RRMS PBMCs [30], Murine EAN & macrophage cell line [94], Human psoriatic T cells & murine EAE [104], Murine intracerebral hemorrhage models [131] |
| IL-12p40 [61,62] | ↓ | Human psoriatic T cells & murine EAE [104], Human PBMCs [123], Murine BMDCs [124], Murine primary microglia [142] |
| IL-12p70 [62] | ↓ | Murine ischaemic stroke model [93], Human PBMCs [123,143] |
| IL-13 [40,61,62] | ↓ | Murine BMDMs [117], Murine splenocytes [115] |
| IL-17 [40] | ↓ | Murine EAE [8,104], Human PBMCs [24], Human RRMS PBMCs [18,25,144], Murine ischaemic stroke model [93], Murine EAN & macrophage cell line [94], Human psoriatic T cells [104], Murine splenocytes [121], Murine BMDCs [124] |
| IL-18 [61,62] | ↓ | Murine SCD model [130], Murine renal I/R injury model [136] |
| IP-10 [40,62] | ↓ | Human keratinocytes & PBMCs [23], Murine splenocytes [59], Murine ischaemic stroke model [93], Human primary ASMCs [110,112], Primary human & murine astrocyte cultures [134], Human tumour biopsies, cancer cell lines & oncolytic viruses [145] |
| MCP-1 [3,40,61] | ↓ | Murine splenocytes [59], Human HIV-infected monocyte-derived macrophages [92], Human UVECs [116,120], Primary human & murine astrocyte cultures [134], Primary murine microglia [142] |
| MCP-3 [62] | ↓ | Murine liver I/R injury model [137] |
| MIG [3,61,62] | ↓ | Human keratinocytes & PBMCs [23] |
| MIP-1α [40,61,62] | ↓ | Murine splenocytes [59] |
| MIP-1β [62] | ↓ | Murine splenocytes [59] |
| PDGF-BB [40,61,62] | ↓ | Human UVECs [115] |
| RANTES [61] | ↓ | Murine splenocytes [59], Murine ischaemic stroke model [93], Primary human asthmatic ASMCs [111], Primary human ASMCs [113], Human UVECs [116] |
| TNF-α [40,61] | ↓ | Human RRMS PBMCs [16,17,19,21,22], Murine splenocytes [59,115], Human HIV-infected monocyte-derived macrophages [92], Murine EAN & macrophage cell line [94], Murine hepatotoxicity model [101], Murine BMDMs [117], Murine hepatic injury & Kupffer cells [119], Murine BMDCs & allogeneic splenic T cell co-culture [122], Healthy human PBMCs [123], Murine colitis model [126], Murine epilepsy model [127], Primary murine microglial & astroglial co-cultures [128,129], Murine & primate SCD models [135], Murine renal/liver I/R injury model [136,137], Murine experimental sepsis [139], Primary murine microglia [142] |
| TNF-β [61,62] | ↓ | Murine BMDMs [117] |
| VEGF [40] | ↓ | Murine ischaemic stroke model [93], Murine osteoblastic cells [106], Murine splenocytes [115], Murine liver I/R injury model [137] |
Abbreviations: ASMC: Airway smooth muscle cell; BMDC: Bone marrow-derived cells; BMDM: Bone marrow-derived macrophages; CBDMC: Cord blood-derived mast cells; DMF: Dimethyl fumarate; EAE: Experimental autoimmune encephalomyelitis; EAN: Experimental autoimmune neuritis; G-CSF: Granulocyte-colony stimulating factor; GM-CSF: Granulocyte-macrophage-colony stimulating factor; IDD: Intervertebral disc degeneration murine; IFN: Interferon; IL: Interleukin; I/R: Ischemia/reperfusion; MCP: Monocyte chemoattractant protein; MIG: Monokine induced by gamma interferon; MIP: Macrophage inflammatory protein; PBMC: Peripheral blood mononuclear cells; PDGF-BB: Platelet derived growth factor-BB; RANTES: Regulated on Activation, Normal T Cell Expressed and Secreted; RRMS: Relapsing-Remitting Multiple Sclerosis; SCD: Sickle cell disease; TNF: Tumour necrosis factor; UVEC: Umbilical vascular endothelial cells; VEGF: Vascular endothelial growth factor. Note: DMF increases IL-4 and IL-10 which are anti-inflammatory cytokines- these cytokines are already elevated in COVID-19.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Timpani, C.A.; Rybalka, E. Calming the (Cytokine) Storm: Dimethyl Fumarate as a Therapeutic Candidate for COVID-19. Pharmaceuticals 2021, 14, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14010015
AMA Style
Timpani CA, Rybalka E. Calming the (Cytokine) Storm: Dimethyl Fumarate as a Therapeutic Candidate for COVID-19. Pharmaceuticals. 2021; 14(1):15. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14010015
Chicago/Turabian StyleTimpani, Cara A., and Emma Rybalka. 2021. "Calming the (Cytokine) Storm: Dimethyl Fumarate as a Therapeutic Candidate for COVID-19" Pharmaceuticals 14, no. 1: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14010015
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.