Antifibrotic and Anti-Inflammatory Actions of α-Melanocytic Hormone: New Roles for an Old Player


1
Division of Pulmonary and Critical Care Medicine, Johns Hopkins University, Baltimore, MD 21218, USA
2
Division of Pulmonary and Critical Care, University of Miami, Miami, FL 33146, USA
*
Author to whom correspondence should be addressed.
Pharmaceuticals 2021, 14(1), 45; https://0-doi-org.brum.beds.ac.uk/10.3390/ph14010045
Submission received: 25 December 2020
/
Revised: 4 January 2021
/
Accepted: 6 January 2021
/
Published: 8 January 2021
(This article belongs to the Special Issue Lung Injury and Repair)
Abstract
:The melanocortin system encompasses melanocortin peptides, five receptors, and two endogenous antagonists. Besides pigmentary effects generated by α-Melanocytic Hormone (α-MSH), new physiologic roles in sexual activity, exocrine secretion, energy homeostasis, as well as immunomodulatory actions, exerted by melanocortins, have been described recently. Among the most common and burdensome consequences of chronic inflammation is the development of fibrosis. Depending on the regenerative capacity of the affected tissue and the quality of the inflammatory response, the outcome is not always perfect, with the development of some fibrosis. Despite the heterogeneous etiology and clinical presentations, fibrosis in many pathological states follows the same path of activation or migration of fibroblasts, and the differentiation of fibroblasts to myofibroblasts, which produce collagen and α-SMA in fibrosing tissue. The melanocortin agonists might have favorable effects on the trajectories leading from tissue injury to inflammation, from inflammation to fibrosis, and from fibrosis to organ dysfunction. In this review we briefly summarized the data on structure, receptor signaling, and anti-inflammatory and anti-fibrotic properties of α-MSH and proposed that α-MSH analogues might be promising future therapeutic candidates for inflammatory and fibrotic diseases, regarding their favorable safety profile.
Keywords:
α-MSH; melanocortins; α-MSH analogues; anti-inflammatory; anti-fibrotic; MC1R; lung fibrosis1. Introduction
The melanocortin system encompasses melanocortins, five transmembrane G protein-coupled receptors, plus two endogenous antagonists: the agouti-signaling protein and agouti-related peptide. The melanocortins comprise adrenocorticotropic hormone (ACTH), α-, β-, and γ-melanocyte-stimulating hormones (α-, β-, γ-MSHs), which are generated from proopiomelanocortin (POMC) processing. Besides steroidogenesis and pigmentary effects caused by ACTH and α-MSH, respectively, new roles in energy homeostasis, reproductive system functions, exocrine glands secretion, immunomodulatory, and anti-inflammatory, have been explained [1,2].
Our knowledge of immunomodulatory effects of melanocortins has progressed, since the first clinical experiment of ACTH in arthritis and rheumatic fever patients conducted by Hench et al., in 1949 [3]. Prior to identification and cloning of melanocortin receptor family (MCR), ACTH-induced improvement in subjects with arthritis was presumed to be due to activation of the hypothalamus-pituitary-adrenal (HPA) axis and cortisol production. Today, 53 years later, Getting et al. showed that melanocortin-3 receptor (MC3R) signaling, triggered by ACTH, regardless of steroid synthesis, was likewise responsible for ACTH effectiveness in inflammatory arthritis and proposed MC3R agonists as novel therapeutics for chronic inflammatory diseases [4]. This study opened new perspectives on the role of melanocortins in inflammation, and considering their receptors as potential targets for future anti-inflammatory therapies.
One of the most common and burdensome consequences of chronic inflammation is the development of fibrosis. Depending on the regenerative capacity of the involved tissue and the quality of the inflammatory response, the outcome is not always perfect, with development of some fibrosis. Disease states in which fibrosis is the leading cause of mortality and morbidity encompass a broad range of illnesses. These include pulmonary fibrosis, liver fibrosis and cirrhosis, chronic kidney disease, myocardial infarction, systemic autoimmune diseases such as systemic sclerosis, etc. Despite the heterogeneous etiology and clinical presentations, fibrosis in many pathological states follow the same path of activation or migration of fibroblasts, and the differentiation of fibroblasts to myofibroblasts, which produce collagen and α-smooth muscle actin (α-SMA) in fibrosing tissue [5,6,7]. In this review we reported anti-inflammatory, anti-fibrotic and regenerative properties of melanocortins.
2. Melanocortins
2.1. Ligands
The melanocortins are derived from processing of POMC [8]. Interestingly, cloning and sequencing has revealed POMC in “lamprey”, the most ancient vertebrate, that shares similarities to those of higher vertebrates, implying that POMC has an ancient linage, likely dating back millions of years (Reviewed in [9]). Besides pituitary, melanocytes, keratinocytes, and central nervous system (CNS) where POMC was discovered, POMC messenger RNA (mRNA) has been recognized in immune cells such as monocytes and lymphocytes which suggests immunomodulatory roles for POMC-derived peptides [4,10,11]. Proprotein convertase-1 (PC1) catalyzes cleavage of POMC to generate ACTH, and PC2 mediates MSH synthesis in pars intermedia of pituitary, CNS, skin and hair follicles [12]. Each melanocortin peptide is rectified from a different region of POMC. γ-MSH is obtained from the amino-terminus, while α-MSH and ACTH are cleaved from the middle part. β-MSH, β-LPH, γ-LPH, and β-Endorphin are processed from the carboxy-terminal region of POMC [13]. Each of the melanocortin ligands shares the conserved sequence of His-Phe-Arg-Trp (HFRW), which functions as a pharmacophore for MCR signaling [14,15].
ACTH is a 39 amino acid polypeptide, best recognized for its role in physiological stress response [16]. ACTH stimulates the glucocorticoid production by triggering cholesterol conversion to pregnenolone in the cortex of adrenal gland [17]. This sequence is mediated by binding to melanocortin receptor 2 (MC2R), stimulation of the membrane-bound adenylate cyclase and calcium influx [18,19]. With the emerging evidence showing the expression of MC2R in tissues other than adrenal cortex, recent literature have shed light on new roles of ACTH, including lipolytic activity in adipocytes [20], reducing lipid content of cells by knockdown of MC2R and inhibition of peroxisome proliferator-activated receptor gamma 2 (PPARγ2) [21], suppressing leptin expression [22], playing a role in the differentiation of mesenchymal cells [23], regulation of bone mass [24], a role in the maintenance and repair of the vascular extracellular matrix [25], controlling thymocyte homeostasis [26], and amelioration of tumor necrosis factor (TNF)-induced acute kidney injury [27].
α-MSH is composed of 13 amino acids and is best known for its pigmentary effects in skin [28], but also has been indicated to exert anti-inflammatory and microbicidal effects discussed below [29,30,31,32,33,34].
β-MSH and γ-MSH are much less understood, compared with α-MSH and ACTH. It is shown that intraventricular infusion of γ2-MSH suppresses LPS-induced inflammatory responses [35]. Getting et al. demonstrated that natural and synthetic ligands for MC3R (γ2-MSH and synthetic agonist MTII, respectively) in a murine model of experimental gout, inhibit aggregation of chemokine C-X-C motif ligand 1 (CXCL1), polymorphonuclear cells (PMNs), and suppress production of interleukin-1 beta (IL-1β), evoked by monosodium urate crystals in the peritoneal cavity [36]. Similar anti-inflammatory actions of γ-MSH and β-MSH were detected in other investigations [37,38,39].
2.2. Receptors
Effects of melanocortin system ligands are mediated by five transmembrane G protein-coupled receptors, which are named based on the sequence of their cloning. Sequence comparison of MCRs reveals 38 to 60% identity between these receptors [40]. Table 1 demonstrates the diverse tissue distribution of agonists and antagonists of melanocortin receptors. These G protein-coupled receptors (GPCRs) when activated, lead to activation of adenylyl cyclase, which catalyzes the conversion of ATP to cyclic adenosine monophosphate (cAMP) in cytoplasm. cAMP activates protein kinases (mainly protein kinase A, which ends in the phosphorylation of cAMP response element-binding protein (CREBP) [40].
3. Anti-Inflammatory Effects of α-MSH
During the last four decades, many investigations have established that α-MSH has strong anti-inflammatory effects.
3.1. Fever and Multiple Organ Dysfunction Syndrome (MODS)
In 1981, Glyn and Lipton showed that five micrograms of intravenous (IV) or intracerebroventricular α-MSH reduces fever produced by leukocytic pyrogen in rabbits [84]. Thereafter, other studies further confirmed the antipyretic effects of α-MSH in guinea pigs and squirrel monkeys [85,86].
In another study, Bitto et al. showed that intraperitoneal injection of NDP-α-MSH (340 µk/kg) in LPS-induced MODS model significantly decreased expression of tumor necrosis factor-α (TNF-α), increased expression of IL-10, and reduced serum levels of TNF-α and improved survival [87]. Other investigations have further confirmed the therapeutic efficacy of α-MSH in septic shock, systemic inflammatory response syndrome, and cardiac arrest [64,88,89,90,91,92].
3.2. α-MSH and the Respiratory System
Recent analyses have recognized expression of the MC1R and MC3R in alveolar macrophages in mice [61,93]. α-MSH inhibited leukocyte migration to the lungs in a lypopolysaccharide-induced acute lung injury in rats [94]. Deng et al. investigated the protective properties of 25 µg of IV α-MSH at zero, 8, and 16 h after clamping renal arteries for 40 min and reperfusion in a mice acute lung injury model. This study reported that α-MSH administration improved tissue injury, inhibited production of intracellular adhesion molecule-1 (ICAM-1) and TNF-α in lungs, and turned off inflammatory transcription factors and stress-induced genes [95]. In agreement with previous literature, Miao and co-workers found that 17 mg/kg IV α-MSH has anti-apoptotic effects on vascular endothelial cells in rat model of acute respiratory distress syndrome [96].
In another study, Colombo et al. examined the effects of 100 µg intraperitoneal (IP) α-MSH analogue (NDP-α-MSH) on a bleomycin-induced ALI model. After instillation of 1 mg bleomycin into the trachea, ten of the bleomycin recipients received α-MSH analogue. NDP-α-MSH offset bleomycin-induced edema in lung tissue (lung weight in control vs. α-MSH group, 5.8 ± 0.5 vs. 3.9 ± 0.1, respectively, p < 0.05) and mediated transcriptional modifications in genes involved in fluid handling, including activation of Na+/K+ ATPase and ENaC (epithelial sodium channels). Moreover, decreased expression of TNF-α, IL-6, transforming growth factor-β, and iNOS was noticed [97].
Raap et al. sensitized mice by three IP injections of 10 µg ovalbumin (OVA) on days 1, 14, and 21. Injections of α-MSH (1 mg/kg body weight) were performed 30 min before sensitization or allergen aerosolization. Mice received 2 allergen challenges implemented by an exposure to diluted 1% OVA on days 27 and 28. Significant decreases in the serum levels of allergen-specific immunoglobulin E (p < 0.05), IgG1 (p < 0.05), and IgG2a (p < 0.001) were identified in intervention group compared to controls [98]. In a similar study, Webering et al. measured α-MSH levels in bronchoalveolar lavage (BAL) fluid of asthmatic versus non-asthmatic participants in addition of healthy and asthmatic mice (OVA-induced eosinophilic airway inflammation model). α-MSH was delivered intratracheally and α-MSH antibody was injected immediately before each sensitization, for neutralization of endogenous α-MSH. Results revealed that α-MSH levels were significantly higher in participants with eosinophilic asthma than in healthy subjects. The OVA-induced asthmatic mice also showed high α-MSH levels in BAL fluid. Additional administration of α-MSH to OVA-sensitized mice significantly reduced eosinophils and lymphocytosis in BAL as well as inflammation in airways. This study also revealed MC5R expression in airway epithelium [99].
Literature have confirmed that C-terminal tripeptide derivatives of α-MSH (e.g., KPV (Lys-Pro-Val, the C-terminal sequence of alpha-MSH) and KdPT (Lys-D-Pro-Thr), possess anti-inflammatory properties with no pigmentary effects [100]. In an immortalized human bronchial epithelial cell model challenged with rhino-syncytial virus, 1 µg of IV KPV suppressed intracellular and systemic proinflammatory signaling (TNF-α–dependent NF-κB–driven reporter activity and IL-8, respectively) and reduced the activity of matrix metalloprotease-9 (MMP-9) responsible for lung remodeling via MC3R, in a dose-dependent way [101].
Zhang et al. tested the effects of α-MSH in a novel in experimental sarcoid model. The granuloma model was developed by mycobacterium-challenged peripheral blood mononuclear cells (PBMCs) obtained from sarcoidosis patients. Both challenged and non-challenged PBMCs were treated with 10 μM α-MSH daily or saline. RNA-Seq analysis on 3rd day after exposure to α-MSH revealed significant decrease in IL-1b, IL-1R, IL-8, IL-12, chemokine C-C ligand 3 (CCL3), CCL4, CCL5, GM-CSF, IFN-γ, and TNF-α levels in intervention group. They reported a significantly increase expression of p-CREB in α-MSH-treated experimental sarcoidosis model. Furthermore, addition of a highly selective CREB inhibitor (666-15), significantly counterbalanced the effects of α-MSH, suggesting that CREB phosphorylation is essential for anti-inflammatory effects of α-MSH [31].
3.3. α-MSH and the Eye
In the immune-privileged eye, melanocortins are involved in regulation of inflammatory pathways (cytoprotection) and promoting the immune tolerance [102]. An experiment conducted by Lee et al. showed that α-MSH treatment of mice with experimental autoimmune uveoretinitis inhibited inflammation and may re-define some aspects of immune privilege [103]. Other studies also supported this finding [34,104,105].
It has been shown that intravitreal injection of 3 µL of α-MSH at week 1 and 3 after streptozotocin-induced hyperglycemia inhibited breakdown of blood-retina barrier and vascular permeability through MC4R signaling in diabetic retinas [106]. Additionally, 10 µg/3µL of intravitreal α-MSH normalized levels of H2O2, reactive oxygen species (ROS), and the total antioxidant capacity and corrected the aberrant changes in endothelial nitric oxide synthase (eNOS), inducible nitric oxide synthase (iNOS), intracellular adhesion molecule 1 (ICAM-1), and TNF-α expression levels in diabetic retinas [107]. Zhang and co-workers indicated that anti-inflammatory actions of melanocortins in the eye are caused by under-expression of inflammatory cytokines (i.e., TNF-α and IL-6) and suppression of the NFκB-dependent signaling pathway [108].
3.4. α-MSH and Gastrointestinal System
KPV can weaken the inflammatory responses in colonic epithelium and intestinal immune cells and lead to reduction in the incidence of inflammatory bowel diseases (IBD) in vivo [109,110,111,112]. Dalmasso et al. investigated effects of 100 µM KPV on mice with experimental colitis induced by dextran sulfate sodium (DSS) and trinitrobenzene sulfonic acid (TNBS). As an index of neutrophilic infiltration intestinal myeloperoxidase (MPO) activity was assessed. They proved that treatment with oral KPV decreased MPO activity by ~50% and these results were confirmed by hematoxylin and eosine (H&E) examination of colonic slides. They also proposed a non-receptor-dependent immunoregulatory effect of KPV, mediated by a transporter normally expressed in the small bowel and induced in colitis [113]. Another similar study showed that KPV treatment leads earlier recovery and significantly stronger regain of body weight (to 87.8% ± 2.7%, versus 73.9% ± 3.5% of the original body weight in the intervention and control mice). Interestingly, on day 14 after the administration of DSS, the body weight of KPV-treated mice had standardized to 102.4–0.9%. treatment with KPV, significantly reduced myeloperoxidase activity (881.7 ± 215.9 U/mg protein) compared to control group (1835.9 ± 283.8 U/mg protein) (p < 0.05) [114]. Recently, Xiao et al. tested the effects of targeted hyaluronic acid-based KPV delivery (HA-NP) in experimental ulcerative colitis mouse model. They proposed that Hyaluronic acid lysine-proline-valine nanoparticles (HA-KPV-NPs) applies combined mechanisms against ulcerative colitis by both enhancing mucosal healing and regulating inflammatory responses. Furthermore, oral HA-KPV-NPs encapsulated in a hydrogel exhibited a higher potency to prevent epithelial injury [115]. Targeted theranostic NDP-α-MSH delivery in IBD has further opened up possibilities for therapeutic and selective use in other inflammatory disease, such as the lung inflammation found in COVID-19 [116].
3.5. α-MSH and Nervous System
Min et al. generated mature monocyte-derived dendritic cells (MoDCs), using TNF-α. Then, they treated MoDCs with different dosages of α-MSH (10−14–10−6 M) to assess the regulatory impact of α-MSH on TNF-α-DCs. The downregulation of CD86, CD83, IL-12 and over-expression of IL-10 was observed in all doses after treatment with α-MSH. They showed upregulation of Annexin A1 after administration of α-MSH, suggesting an inhibitory effect of α-MSH on TNF-α-induced MoDC maturation via the upregulation of Annexin A1 [117]. In vitro studies favor a possible neuroprotective role for melanocortins, as they suppressed NF-kB activation in TNF-α-activated Schwann cells or lipopolysaccharide-activated glioma cells [29,118,119,120]. Mykicki et al. showed that mice treated systemically with α-MSH in two-day intervals were immune from developing clinical signs of experimental autoimmune encephalitis (EAE), and this effect was associated with reduced inflammatory foci and decreased central nervous system demyelination (p < 0.0001, p = 0.0051). T-helper 1 and T-helper 17 cells were diminished in the CNS and in the cervical lymphatics from α-MSH-treated animals compared with controls. They found that NDP-MSH induced functional regulatory T cells through MC1R signaling, leading to alleviation of EAE in treated mice [121]. Carniglia et al. evaluated the effect of NDP-α-MSH on PPAR-β and PPAR-γ expression in rat’s astrocytes and microglial cells. They recognized that microglial cells of rat express MC4R and treatment with NDP-α-MSH strongly enhances PPAR-γ expression and decreases PPAR-β expression in both microglia and astrocytes [122]. Wang et al. produced a novel Tat protein(TAT)-human serum albumin (HAS)--α-MSH fusion protein. They showed that the fusion protein TAT-HSA-α-MSH can successfully cross the blood-brain barrier after intraperitoneal injection. The NF-κB driven reporter assay in vitro showed that TAT-HSA-α-MSH strongly suppressed NF-κB in the glioma cell line. In LPS-induced CNS inflammation in mice, HSA-α-MSH, when given intraperitoneally, markedly attenuated TNF-α production. These results confirmed that the fusion protein TAT-HSA-α-MSH exerts dominant anti-inflammatory activities in the nervous system after being delivered into the animal’s brain [29]. Effectiveness of melanocortins in neuroinflammation is further confirmed by other studies [30,118,119,123,124].
3.6. α-MSH and Skin
Kleiner et al. showed that the analogues of MC1R and MC5R have regulatory effects on IgE-mediated allergic inflammation [127]. Other studies showed similar therapeutic effects of MC1R agonists on atopic dermatitis mouse model, and induction of regulatory T-cells in vitro and in vivo which led to inhibitory effect on psoriasis progression in a mouse model [128,129,130].
3.7. α-MSH and the Musculoskeletal System
Capsoni et al. evaluated the role of melanocortin in regulating production of inflammatory cytokines, metalloproteinases (MMPs), tissue inhibitors of MMPs (TIMPs), iNOS, and nitric oxide (NO) in response to IL-1β and TNF-α in synovial chondrocytes. They demonstrated increased TIMP-3 gene expression and downmodulation of TNF-α-induced stimulation of synovial chondrocytes [131]. Immuno-regulatory and anti-degenerative actions of melanocortins in joints have been extensively studied [33,132,133,134,135].
Synovial fluid α-MSH levels have shown a negative independent correlation with disease severity in individuals with post-traumatic osteoarthritis and application of local α-MSH has been suggested as a potential adjuvant therapy [136]. Interestingly, two studies have revealed a therapeutic property of melanocortins (including ACTH), in osteonecrosis of bones [137,138].
3.8. Other Anti-Inflammatory Actions
Various studies have proposed that α-MSH protects against ischemia and reperfusion injuries in kidney, testes, myocardium, intestines, and CNS [139,140,141,142,143].
The melanocortins show therapeutic characteristics in atherosclerosis by preventing plaque rupture and improving endothelial cell function, which may suggest a novel therapeutic target for atherosclerosis [135].
Beneficial effects of α-MSH on endothelial production of pro-inflammatory substances for applications in implantable intravascular devices such as pacemakers, have been reported [144,145]. One study reported allograft protection in organ recipient mice [146].
Liu et al. attempted interpreting the role of α-MSH in adipose tissue inflammation and the interactions with forkhead box proteins. By suppressing forkhead box protein expression, α-MSH could dampen LPS-induced inflammation accompanied with increased anti-inflammatory mediator release and decreased inflammatory products [147].
4. Mechanisms of Anti-Inflammatory Effects of α-MSH
The mechanisms of anti-inflammatory actions have been extensively studied (briefly listed below and demonstrated in Figure 1).
4.1. Inhibition of NF-κB
Manna and Aggrawal first described that α-MSH nullified TNF-mediated NF-κB activation in a concentration-dependent fashion and opposed NF-κB activation induced by LPS. NF-κB is an evolutionarily conserved transcription factor that regulates immune responses. Its role as a master regulator of the inflammatory response stems from its critical function in regulating the expression of hundreds of immune relevant genes, particularly those encoding proinflammatory mediators, in addition to other genes important for the development of the immune system. The inhibitory effect of α-MSH appears to be performed through cAMP production, as inhibitors of adenylyl cyclase and of PKA antagonized its anti-inflammatory effects [148]. Subsequently, similar effects on various cell types, including pulmonary epithelial cells, were reported (Reviewed in [4]). Several mechanisms have been proposed to explain how cAMP interferes with the NF-κB signaling cascade. These mechanisms include effects of cAMP signaling on IκB kinase activation and cytoplasmic IκB levels, posttranslational cAMP-induced Rel proteins modification induced, effects of cAMP on NF-κB dimer composition, etc., (reviewed in [149]). cAMP-independent inhibition of nuclear translocation of NF-κB through MC1R signaling has also been explained [150].
4.2. Suppression of Proinflammatory Cytokines
α-MSH acts as an anti-inflammatory substance by suppression of proinflammatory mediators such as TNF-α, interferon-γ (IFN-γ), IL-1, IL-6, IL-8 and induction of cytokine suppression by production of IL-10 [51,151,152]. The present evidence indicates that α-MSH has a key role in the regulation of TNF-α and nitric oxide in monocytes and macrophages. Antibodies against MC1R increased TNF-α in non-challenged macrophages, blunted the hindering effect of α-MSH, and enhanced TNF-α production in LPS-challenged cells [51]. IL-8 gene transcription demands activation of the combination of both NF-κB and activating protein-1 (AP-1), or that of NF-κB, and another transcription factor, NFκβ/Interleukin-6 [153]. It has been shown that AP-1 in dermal fibroblasts can be modified by α-MSH, and that this effect appears to be co-stimulus-dependent [154]. Suppression of IL-1 and IL-6 mRNA expression by α-MSH might be due to suppression of the NF-κB signaling pathway, which is the key factor in production of these proinflammatory cytokines [155]. IL-10 is an anti-inflammatory cytokine that plays a key role in maintaining the balance of immune responses and resolves inflammation and blunts unwanted tissue injury [156]. Toll like receptor (TLR) signaling results in stimulation of NF-κB and mitogen-activated protein kinase (MAPK) pathways (ERK1/2 and p38), which subsequently lead to production of IL-10. MAPKs activate mitogen- and stress-activated kinase 1 (MSK1) and MSK2 and phosphorylate the transcription factors AP-1 and CREB, which subsequently result in IL-10 expression [157]. Additionally, a cAMP signaling cascade can lead to CREB phosphorylation and transcription of a plethora of genes besides IL-10 [149]. Therefore, CREB has a central role in the production of IL-10. α-MSH-mediated cAMP cascade signaling and subsequent CREB phosphorylation is a possible mechanism for IL-10 production.
4.3. Inhibition of Adhesion Molecules
The ability of α-MSH to suppress expression of intercellular adhesion molecule-1 (ICAM-1) has been explained in murine mast cells. The inhibition of vascular cell adhesion molecule-1 (VCAM-1) and E-selectin expression were also identified in endothelial cells. Moreover, α-MSH regulates the expression of co-stimulatory molecules essential for antigen presentation (CD40 and CD86) in monocytes and DCs [107,158,159,160,161].
4.4. Suppression of Non-Cytokine Inflammatory Mediators
α-MSH has been reported to suppress proinflammatory non-cytokine regulators such as nitric oxide (NO), prostaglandin E (PGE), and ROS. The capacity of α-MSH in damping stimulated nitric oxide synthesis and iNOS expression was first reported in MC1R expressing mice macrophages [162]. Thereafter, similar results have been described in Raw 264.7 cells, helper T-cells, PBMCs, melanoma cells, mice microglia, and astrocytes [151,152,163,164]. In Mandrika et al.’s experiment, forskolin (a pharmacological agent that raises intracellular cAMP) was also able to inhibit nitric oxide production without affecting the translocation of active IκB-free NF-κB, suggesting that cAMP generation may inhibit NO synthesis, independently of NF-kB signaling [150]. Oktar et al. showed that non-selective cyclooxynegase (COX) inhibitor indomethacin antagonized the effect of melanocortins, at a dose which did not influence the high lucigenin (a chemiluminescent probe used to detect superoxide production) chemiluminescence value of stimulated PMNs. However, the inhibitory effects of α-MSH in lucigenin values were not altered in cells treated with more selective COX inhibitors, like ketorolac or nimesulide. Although the mechanisms of interaction between α-MSH and COX are not clear yet, this study proposed that α-MSH prevents superoxide synthesis PMNs and that both COX1 and COX2 are involved in this effect [165].
4.5. Induction of Regulatory T Cells (Tregs)
α-MSH has been reported to induce CD4 and CD25 positive regulatory T cells, which have a central role in maintenance of immune tolerance [166,167,168]. α-MSH-treated Tregs have been shown to inhibit IFN-γ and IL-10 synthesis but increase TGF-β1 synthesis [82]. α-MSH-induced immune regulation arises from converting effector T lymphocytes. These regulatory cells are CD25+ CD4+, CTLA4+, CD44+, CD62L+, and latency associated peptide (LAP) positive. α-MSH induces TGF-β synthesis but does not exert immunoregulation in naive T-cells. This indicates that the immunoregulatory actions of α-MSH on T-cells are confined to antigen-experienced effector T-cells. Thus, it has been suggested that it is possible to use melanocortins to induce antigen-specific regulatory T-cells which aim autoimmune diseases [82,168,169,170]. The immunomodulatory effects of α-MSH in T-cells is mediated through MC5R, which subsequently activates the Janus kinase 2 (JAK2), signal transducer and activator of transcription 1 (STAT1), or ERK pathways in immune cells and lead to cell differentiation and cytokine production [81]. Emerging evidence suggests a role for CREB in TGF-β/FoxP3–dependent Treg induction and maintenance (reviewed in [157]).
4.6. Promotion of Efferocytosis
Effective clearance of apoptotic cells by macrophages and other phagocytes is a fundamental component in homeostasis and resolution of inflammation, termed efferocytosis [171]. For many decades, resolution of inflammation was regarded as a passive process, simply including removal of inflammatory stimulus, stopping production of inflammatory mediators, and the inhibition of further chemotaxis to injury site. Later, Sehran and Savill proposed that resolution of inflammation is an active process which also consists of signaling pathways associated with apoptosis, efferocytosis, and reprogramming of macrophages to ensure a regaining of the preinflammatory status [172] Montero–Mendelez and co-workers evaluated the effect of AP214 (α-MSH analogue with a higher affinity to MC1R and MC3R) on phagocytosis and apoptotic neutrophils in mouse peritonitis model. They showed that 400–800 µg/kg body weight of i.p. AP214, can increase phagocytosis of apoptotic neutrophils by 70 and 30% in In Vitro and In Vivo, respectively. They proposed a role for MC3R in efferocytosis [65].
5. α-MSH and Tissue Repair and Remodeling
Replacement of damaged tissue with new living tissue is referred to as tissue repair (healing). The basic cellular and molecular mechanisms underlying restoration of tissue architecture and function after an injury and its failure to heal are still poorly understood, and treatments are dissatisfying. Defective tissue repair after trauma, surgery, and acute or chronic disease states affect millions of people worldwide each year and arises from malregulation of tissue repair responses, including inflammation, angiogenesis, matrix deposition and degradation, and cell recruitment. Impaired repair can lead to fibrosis and organ dysfunction. Several possible anti-fibrotic properties of α-MSH are discussed below (summarized in Figure 2).
Over the past few decades, transforming growth factor-β (TGF-β) may have been the best studied cytokine in fibrosis and has been a prototypical “profibrotic” mediator [173]. TGF-β1 regulates fibroblast recruitment to sites of tissue injury and mediates fibroblast-to-myofibroblast differentiation [174]. Bohm et al. showed that α-MSH reduces the extracellular levels of procollagens I, III, and V by 70% in human dermal fibroblasts and strongly reverses the encouraging actions of TGF-β1 on the extracellular matrix (ECM) collagen levels, with the most predominant effects on collagens I and III and more blunted effects on collagen V [175]. α-MSH inhibits IL-1β-mediated IL-8 secretion [154], exerts cytoprotective effects [176], and suppresses experimentally induced cutaneous fibrosis in dermal fibroblasts [177]. Kokot et al. developed an animal model of scleroderma induced by subcutaneous injection of bleomycin and treated the mice with 5 µg/day of subcutaneous α-MSH for 21 days. The administration of α-MSH inhibited expression of type I and type III collagens induced by bleomycin [177]. Bleomycin-treated mice with defective α-MSH and MC1R signaling showed increased cutaneous collagen type I mRNA levels accompanying cutaneous fibrosis [178]. In addition, MC1R mRNA expression levels in keloid fibroblast cell lines were reduced to less than 50%, in comparison with the normal fibroblasts [157], and α-MSH administration to keloid fibroblasts did not inhibit TGF-β1-mediated collagen synthesis and myofibroblast differentiation as much as in the control group, seemingly because of defective expression of MC1R in keloid fibroblasts [179]. de Souza et al. demonstrated that intraperitoneal administration of 1 mg/kg α-MSH immediately before skin wounding significantly reduces the quantity of leukocytes, mast cells, and fibroblasts at the site of injury. α-MSH reduced scar area and enhanced the orientation of the collagen fibers, suggesting it may command the healing process to a more regeneration and less scar formation pathways [180].
Hepatic fibrosis arises from escalating deposition of extracellular matrix components in the hepatic parenchyma due to recurring tissue injury. Lee et al. developed a mouse model of hepatic fibrosis with administration of carbon tetrachloride (CCl4) for 10 weeks. α-MSH expression vector was delivered via electro-permeabilization after full-blown liver fibrosis. Histologic examination and assessment of extracellular matrix contents of the livers revealed that transfected animals markedly reversed CCL4-induced fibrosis, compared to untreated animals (collagen content in intervention group was 23.7 ± 4.7 vs. 59.7 ± 5.0 μg/mg in untreated animals, p < 0.01). The over-expression of TGF-β1, collagen 1, fibronectin, TNF-α, ICAM-1, and VCAM-1 mRNA were reported in the experimental models of hepatic fibrosis. Gene therapy with α-MSH significantly attenuated this over-expression. They further showed that the intervention reversed established hepatic cirrhosis by increasing MMP activity and decrease in their tissue inhibitors (TIMP), suggesting that extracellular matrix metabolism modification might play a role in the tissue repair properties of α-MSH [181]. Wang et al. introduced a liver fibrosis model induced by chronic thioacetamide (selective hepatotoxin) administration and investigated the effects of α-MSH gene therapy on tissue remodeling. Hepatic ECM collagen content in the treated animals was 32.2 ± 6.2 μg/mg while it was 71.6 ± 10.0 μg/mg in control group (p < 0.01). Treatment significantly inhibited TGF-β1, procollagen I, TNF-α, ICAM-1, VCAM-1 and TIMP-1 mRNA over-expression in intervention group. They proposed that the collagenolytic actions of α-MSH can be due to MMP and TIMP balance modulation [182]. Lonati et al. aimed to experiment if treatment with melanocortin adjusts tissue remodeling after performing partial hepatectomy (PH) or sham procedure in rats. Immediately prior to surgery the intervention group received a single dose of NDP-MSH, while controls received only saline. RT-PCR analyses demonstrated that NDP-MSH altered the expression of a substantial proportion of transcripts, including multiple cytokines and their receptors. The critical signaling pathway IL-6/STAT/SOC was significantly enhanced by the α-MSH agonist [183]. Another older study had shown the regenerative effects of α-MSH on hepatectomized rats [184].
Xu and co-workers investigated the anti-fibrotic properties of an α-MSH analogue (STY39) on a bleomycin-induced lung fibrosis murine model. Mice received STY39 (0.625, 1.25, or 2.5 mg/kg, IP) once daily for 2 weeks. multiple items associated with inflammatory pathways, extracellular matrix (ECM) components, myofibroblast proliferation, and tissue remodeling were assessed. They found that α-MSH analogue predominantly improved the survival rates of animals with severe bleomycin-induced lung fibrosis, opposed weight loss, reduced the expression of types I and III procollagen mRNA, blunted myofibroblast differentiation and proliferation, and reduced pulmonary fibrosis. Further evaluation showed that STY39 administration inhibited neutrophil migration into the lungs, inhibited the production of local TNF-α, IL-6, macrophage inflammatory protein 2, and TGF-β, and modified MMP-1/TIMP-1 ratio [185].
Lee et al. evaluated the anti-fibrotic properties of an α-MSH agonist (STY39) on a cyclosporine-induced tubulointerstitial fibrosis rat model. STY39 counterbalanced the Bax and TGF-α increase and induced synthesis of anti-apoptotic Bcl2 protein, as well as inhibition of inflammation and tubulointerstitial renal fibrosis [186].
Verhaagen et al. have explained the effects of α-MSH in nerve regeneration [187]. This is also confirmed by Dekker et al., who injected 10 µg of α-MSH into rats every 48 h after a sciatic nerve crush and tested the number of myelinated axons in cross sections of sciatic nerve at several time points and observed that α-MSH increased the number and diameter of axons after nerve injury [188]. Later, the effectiveness of α-MSH on peripheral nerve regeneration was further established [189,190,191].
Bonfiglio et al. investigated the effects of KPV on corneal wound re-epithelization in rabbits and the potential role of nitric oxide. Denuded corneas of rabbits were treated four times a day with KPV 1, 5, or 10 mg/mL (30 mL) or sodium nitroprusside (NO donor) instantaneously after corneal abrasion while control group only received normal saline. Then, 60 hours later, 100% of the corneas treated with KPV and SP were fully re-epithelized while none from untreated rabbits were re-epithelized. They concluded that the availability of nitric oxide might be of specific importance in therapeutic efficacy of topical KPV in experimental corneal abrasion model [161]. Pavan et al. evaluated the influence of topical α-MSH on the healing of corneal wound healing in rats. Topical α-MSH eye-drop in a concentration of 1 × 10−4 mg/mL improved corneal wound healing significantly, while non from control group were healed [192]. Zhang and co-workers tested the anti-fibrotic effect of α-MSH on TGF-β1-stimulated human Tenon’s capsule fibroblasts (HTFs) since these fibroblasts play a central role in the initiation and handling of wound healing and tissue remodeling after trabeculectomy. α-MSH inhibited the proliferation of TGF-β1-induced HTFs in a concentration-dependent fashion and demonstrated inhibitory effect on the mRNA expression of type I collagen, TNF-α, ICAM-1, and VCAM-1, which were upregulated by TGF-β1. They proposed an opposite effect of α-MSH on the disparity between MMPs and TIMPs compared with TGF-β1 [193].
6. Future Perspectives
Tissue injury and inflammation are crucial triggers for either regeneration or fibrosis. Melanocortin agonists might have favorable effects on the processes leading from injury to tissue inflammation, from inflammation to tissue fibrosis, and from fibrosis to organ dysfunction. α-MSH may have significant potentials in inflammation control and repairment process in numerous inflammatory lung diseases including sarcoidosis, interstitial lung disease, and COVID-19 related pulmonary fibrosis, with fewer safety concerns than other immunomodulatory medications. Validation via further investigation is recommended to prove the therapeutic properties of MSH agonists in lung diseases.
Author Contributions
Conceptualization, M.M.; methodology and data collection, R.D.; writing—original draft preparation, R.D.; writing—review and editing, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
Authors would like to thank April Mann for critical review and edits.
Conflicts of Interest
Dinparastisaleh declares no conflict of interest. Mirsaeidi is Advisor Board of Mallinckrodt and received grant to study sarcoidosis.
References
- Tao, Y.X. Melanocortin receptors. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2411–2413. [Google Scholar] [CrossRef] [PubMed]
- Gallo-Payet, N. 60 YEARS OF POMC: Adrenal and extra-adrenal functions of ACTH. J. Mol. Endocrinol. 2016, 56, T135–T156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hench, P.S.; Kendall, E.C.; Slocumb, C.H.; Polley, H.F. The effect of a hormone of the adrenal cortex (17-hydroxy-11-dehydrocorticosterone: Compound E) and of pituitary adrenocortical hormone in arthritis: Preliminary report. Ann. Rheum. Dis. 1949, 8, 97–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Guo, D.Y.; Lin, Y.J.; Tao, Y.X. Melanocortin Regulation of Inflammation. Front. Endocrinol. 2019, 10, 683. [Google Scholar] [CrossRef]
- Smith, P.C.; Martínez, C.; Martínez, J.; McCulloch, C.A. Role of Fibroblast Populations in Periodontal Wound Healing and Tissue Remodeling. Front. Physiol. 2019, 10, 270. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Ma, Q. Myofibroblasts and lung fibrosis induced by carbon nanotube exposure. Part. Fibre Toxicol. 2016, 13, 60. [Google Scholar] [CrossRef] [Green Version]
- Wójcik-Pszczoła, K.; Jakieła, B.; Plutecka, H.; Koczurkiewicz, P.; Madeja, Z.; Michalik, M.; Sanak, M. Connective tissue growth factor regulates transition of primary bronchial fibroblasts to myofibroblasts in asthmatic subjects. Cytokine 2018, 102, 187–190. [Google Scholar] [CrossRef]
- Toda, C.; Santoro, A.; Kim, J.D.; Diano, S. POMC Neurons: From Birth to Death. Annu. Rev. Physiol. 2017, 79, 209–236. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Xu, B.; Wang, M.; Liu, S.; Zhang, Y.; Zhang, C. Pharmacological modulation of MRAP2 protein on melanocortin receptors in the sea lamprey. Endocr. Connect. 2019, 8, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Maddila, S.C.; Busch-Dienstfertig, M.; Stein, C. B Lymphocytes Express Pomc mRNA, Processing Enzymes and β-Endorphin in Painful Inflammation. J. Neuroimmune Pharmacol. 2017, 12, 180–186. [Google Scholar] [CrossRef]
- Chrétien, M.; Mbikay, M. 60 YEARS OF POMC: From the prohormone theory to pro-opiomelanocortin and to proprotein convertases (PCSK1 to PCSK9). J. Mol. Endocrinol. 2016, 56, T49–T62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cawley, N.X.; Li, Z.; Loh, Y.P. 60 YEARS OF POMC: Biosynthesis, trafficking, and secretion of pro-opiomelanocortin-derived peptides. J. Mol. Endocrinol. 2016, 56, T77–T97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, P. 60 YEARS OF POMC: Purification and biological characterisation of melanotrophins and corticotrophins. J. Mol. Endocrinol. 2016, 56, T1–T12. [Google Scholar] [CrossRef] [Green Version]
- Winget, M.D.; Ericson, M.D.; Freeman, K.T.; Haskell-Luevano, C. Single Nucleotide Polymorphisms in the Melanocortin His-Phe-Arg-Trp Sequences Decrease Tetrapeptide Potency and Efficacy. ACS Med. Chem. Lett. 2020, 11, 272–277. [Google Scholar] [CrossRef]
- Mowlazadeh Haghighi, S.; Zhou, Y.; Dai, J.; Sawyer, J.R.; Hruby, V.J.; Cai, M. Replacement of Arg with Nle and modified D-Phe in the core sequence of MSHs, Ac-His-D-Phe-Arg-Trp-NH(2), leads to hMC1R selectivity and pigmentation. Eur. J. Med. Chem. 2018, 151, 815–823. [Google Scholar] [CrossRef]
- Ghaddhab, C.; Vuissoz, J.M.; Deladoëy, J. From Bioinactive ACTH to ACTH Antagonist: The Clinical Perspective. Front. Endocrinol. 2017, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Lindner, J.M.; Suhr, A.C.; Grimm, S.H.; Möhnle, P.; Vogeser, M.; Briegel, J. The dynamics of a serum steroid profile after stimulation with intravenous ACTH. J. Pharm Biomed. Anal. 2018, 151, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, C.; Lalli, E. Impact of ACTH Signaling on Transcriptional Regulation of Steroidogenic Genes. Front. Endocrinol. 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, A.E.; Aesoy, R.; Bakke, M. Role of EPAC in cAMP-Mediated Actions in Adrenocortical Cells. Front. Endocrinol. 2016, 7, 63. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Saarinen, A.M.; Campbell, L.E.; De Filippis, E.A.; Liu, J. Regulation of Lipolytic Response and Energy Balance by Melanocortin 2 Receptor Accessory Protein (MRAP) in Adipocytes. Diabetes 2018, 67, 222–234. [Google Scholar] [CrossRef]
- Betz, M.J.; Hatiboglu, N.; Mauracher, B.; Hadaschik, D.; Sauter, A.; Demmelmair, H.; Koletzko, B.; Beuschlein, F.; Slawik, M. Mc2 receptor knockdown modulates differentiation and lipid composition in adipocytes. Horm. Metab. Res. 2012, 44, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Gioldasi, S.; Karvela, A.; Rojas-Gil, A.P.; Rodi, M.; de Lastic, A.L.; Thomas, I.; Spiliotis, B.E.; Mouzaki, A. Metabolic Association between Leptin and the Corticotropin Releasing Hormone. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, F.; Vahednia, E.; Naderi Meshkin, H.; Kerachian, M.A. The effect of adrenocorticotropic hormone on alpha-2-macroglobulin in osteoblasts derived from human mesenchymal stem cells. J. Cell. Mol. Med. 2020, 24, 4784–4790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belaya, Z.E.; Grebennikova, T.A.; Melnichenko, G.A.; Nikitin, A.G.; Solodovnikov, A.G.; Brovkina, O.I.; Grigoriev, A.U.; Rozhinskaya, L.Y.; Dedov, I.I. Effects of endogenous hypercortisolism on bone mRNA and microRNA expression in humans. Osteoporos. Int. 2018, 29, 211–221. [Google Scholar] [CrossRef]
- Belotti, E.M.; Amweg, A.N.; Matiller, V.; Varela, M.L.; Stassi, A.F.; Velázquez, M.M.L.; Ortega, H.H.; Rey, F.; Salvetti, N.R. Effects of adrenocorticotrophic hormone on the expression of matrix metalloproteinases and their inhibitors in the bovine ovary. Reprod. Fertil. Dev. 2020, 32, 748–762. [Google Scholar] [CrossRef]
- Tishevskaya, N.V.; Gevorkyan, N.M.; Kozlova, N.I. Sensitivity of T-Lymphocytes to Hormones of the Anterior Pituitary Gland. Uspekhi Fiziol. Nauk 2017, 48, 80–90. [Google Scholar]
- Si, J.; Ge, Y.; Zhuang, S.; Wang, L.J.; Chen, S.; Gong, R. Adrenocorticotropic hormone ameliorates acute kidney injury by steroidogenic-dependent and -independent mechanisms. Kidney Int. 2013, 83, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhu, B.; Yin, C.; Liu, W.; Han, C.; Chen, B.; Liu, T.; Li, X.; Chen, X.; Li, C.; et al. Palmitoylation-dependent activation of MC1R prevents melanomagenesis. Nature 2017, 549, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhi, D.; Wang, H.; Ru, Y.; Ren, H.; Wang, N.; Liu, Y.; Li, Y.; Li, H. TAT-HSA-α-MSH fusion protein with extended half-life inhibits tumor necrosis factor-α in brain inflammation of mice. Appl. Microbiol. Biotechnol. 2016, 100, 5353–5361. [Google Scholar] [CrossRef]
- Ramírez, D.; Saba, J.; Turati, J.; Carniglia, L.; Imsen, M.; Mohn, C.; Scimonelli, T.; Durand, D.; Caruso, C.; Lasaga, M. NDP-MSH reduces oxidative damage induced by palmitic acid in primary astrocytes. J. Neuroendocrinol. 2019, 31, e12673. [Google Scholar] [CrossRef]
- Zhang, C.; Chery, S.; Lazerson, A.; Altman, N.H.; Jackson, R.; Holt, G.; Campos, M.; Schally, A.V.; Mirsaeidi, M. Anti-inflammatory effects of α-MSH through p-CREB expression in sarcoidosis like granuloma model. Sci. Rep. 2020, 10, 7277. [Google Scholar] [CrossRef] [PubMed]
- Nieto, J.E.; Casanova, I.; Serna-Ojeda, J.C.; Graue-Hernández, E.O.; Quintana, G.; Salazar, A.; Jiménez-Martinez, M.C. Increased Expression of TLR4 in Circulating CD4+T Cells in Patients with Allergic Conjunctivitis and In Vitro Attenuation of Th2 Inflammatory Response by Alpha-MSH. Int. J. Mol. Sci. 2020, 21, 7861. [Google Scholar] [CrossRef] [PubMed]
- Can, V.C.; Locke, I.C.; Kaneva, M.K.; Kerrigan, M.J.P.; Merlino, F.; De Pascale, C.; Grieco, P.; Getting, S.J. Novel anti-inflammatory and chondroprotective effects of the human melanocortin MC1 receptor agonist BMS-470539 dihydrochloride and human melanocortin MC3 receptor agonist PG-990 on lipopolysaccharide activated chondrocytes. Eur. J. Pharmacol. 2020, 872, 172971. [Google Scholar] [CrossRef] [PubMed]
- Lužnik, Z.; Sun, Z.; Nakagawa, H.; Taylor, A.W.; Jurkunas, U.V.; Yin, J.; Dana, R. Association of α-Melanocyte-Stimulating Hormone With Corneal Endothelial Cell Survival During Oxidative Stress and Inflammation-Induced Cell Loss in Donor Tissue. JAMA Ophthalmol. 2020, 138, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kadiri, J.J.; Thapa, K.; Kaipio, K.; Cai, M.; Hruby, V.J.; Rinne, P. Melanocortin 3 receptor activation with [D-Trp8]-γ-MSH suppresses inflammation in apolipoprotein E deficient mice. Eur. J. Pharmacol. 2020, 880, 173186. [Google Scholar] [CrossRef] [PubMed]
- Getting, S.J.; Allcock, G.H.; Flower, R.; Perretti, M. Natural and synthetic agonists of the melanocortin receptor type 3 possess anti-inflammatory properties. J. Leukoc. Biol. 2001, 69, 98–104. [Google Scholar]
- Gómez-SanMiguel, A.B.; Villanúa, M.; Martín, A.I.; López-Calderón, A. D-TRP(8)-γMSH Prevents the Effects of Endotoxin in Rat Skeletal Muscle Cells through TNFα/NF-KB Signalling Pathway. PLoS ONE 2016, 11, e0155645. [Google Scholar] [CrossRef]
- Gómez-SanMiguel, A.B.; Martín, A.I.; Nieto-Bona, M.P.; Fernández-Galaz, C.; Villanúa, M.; López-Calderón, A. The melanocortin receptor type 3 agonist d-Trp(8)-γMSH decreases inflammation and muscle wasting in arthritic rats. J. Cachexia Sarcopenia Muscle 2016, 7, 79–89. [Google Scholar] [CrossRef]
- Madeira, M.F.; Queiroz-Junior, C.M.; Montero-Melendez, T.; Werneck, S.M.; Corrêa, J.D.; Soriani, F.M.; Garlet, G.P.; Souza, D.G.; Teixeira, M.M.; Silva, T.A.; et al. Melanocortin agonism as a viable strategy to control alveolar bone loss induced by oral infection. FASEB J. 2016, 30, 4033–4041. [Google Scholar] [CrossRef] [Green Version]
- Dores, R.M.; Liang, L.; Davis, P.; Thomas, A.L.; Petko, B. 60 YEARS OF POMC: Melanocortin receptors: Evolution of ligand selectivity for melanocortin peptides. J. Mol. Endocrinol. 2016, 56, T119–T133. [Google Scholar] [CrossRef] [Green Version]
- Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Cone, R.D. The cloning of a family of genes that encode the melanocortin receptors. Science 1992, 257, 1248–1251. [Google Scholar] [CrossRef]
- Chhajlani, V.; Wikberg, J.E. Molecular cloning and expression of the human melanocyte stimulating hormone receptor cDNA. FEBS Lett. 1992, 309, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Cone, R.D. Studies on the physiological functions of the melanocortin system. Endocr. Rev. 2006, 27, 736–749. [Google Scholar] [CrossRef] [PubMed]
- Irani, B.G.; Haskell-Luevano, C. Feeding effects of melanocortin ligands—A historical perspective. Peptides 2005, 26, 1788–1799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, C.; Liu, Y.; Liu, J.; Wang, H.Y.; Liu, A.F.; He, D.Q. Agouti signalling protein (ASIP) gene: Molecular cloning, sequence characterisation and tissue distribution in domestic goose. Br. Poult. Sci. 2016, 57, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Almathen, F.; Elbir, H.; Bahbahani, H.; Mwacharo, J.; Hanotte, O. Polymorphisms in MC1R and ASIP Genes are Associated with Coat Color Variation in the Arabian Camel. J. Hered. 2018, 109, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wikberg, J.E.; Chhajlani, V. Expression of melanocortin 1 receptor in periaqueductal gray matter. Neuroreport 1995, 6, 2193–2196. [Google Scholar] [CrossRef]
- Zhang, Y.; Fang, C.; Wang, R.E.; Wang, Y.; Guo, H.; Guo, C.; Zhao, L.; Li, S.; Li, X.; Schultz, P.G.; et al. A tumor-targeted immune checkpoint blocker. Proc. Natl. Acad. Sci. USA 2019, 116, 15889–15894. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Ju, W.; Tin, T.D.; Kim, J.; Lee, J.S.; Park, C.H.; Kwak, S.H. Effect of BMS-470539 on lipopolysaccharide-induced neutrophil activation. Korean J. Anesthesiol. 2020, 73, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Jang, E.A.; Kim, J.Y.; Tin, T.D.; Song, J.A.; Lee, S.H.; Kwak, S.H. The effects of BMS-470539 on lipopolysaccharide-induced acute lung injury. Acute Crit. Care 2019, 34, 133–140. [Google Scholar] [CrossRef]
- Taherzadeh, S.; Sharma, S.; Chhajlani, V.; Gantz, I.; Rajora, N.; Demitri, M.T.; Kelly, L.; Zhao, H.; Ichiyama, T.; Catania, A.; et al. alpha-MSH and its receptors in regulation of tumor necrosis factor-alpha production by human monocyte/macrophages. Am. J. Physiol. 1999, 276, R1289–R1294. [Google Scholar] [CrossRef] [PubMed]
- Neumann Andersen, G.; Nagaeva, O.; Mandrika, I.; Petrovska, R.; Muceniece, R.; Mincheva-Nilsson, L.; Wikberg, J.E. MC(1) receptors are constitutively expressed on leucocyte subpopulations with antigen presenting and cytotoxic functions. Clin. Exp. Immunol. 2001, 126, 441–446. [Google Scholar] [CrossRef]
- Goldenberg, A.J.; Gehrand, A.L.; Waples, E.; Jablonski, M.; Hoeynck, B.; Raff, H. Effect of a melanocortin type 2 receptor (MC2R) antagonist on the corticosterone response to hypoxia and ACTH stimulation in the neonatal rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R128–R133. [Google Scholar] [CrossRef] [PubMed]
- Nensey, N.K.; Bodager, J.; Gehrand, A.L.; Raff, H. Effect of Novel Melanocortin Type 2 Receptor Antagonists on the Corticosterone Response to ACTH in the Neonatal Rat Adrenal Gland In Vivo and In Vitro. Front. Endocrinol 2016, 7, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grässel, S.; Opolka, A.; Anders, S.; Straub, R.H.; Grifka, J.; Luger, T.A.; Böhm, M. The melanocortin system in articular chondrocytes: Melanocortin receptors, pro-opiomelanocortin, precursor proteases, and a regulatory effect of alpha-melanocyte-stimulating hormone on proinflammatory cytokines and extracellular matrix components. Arthritis Rheum. 2009, 60, 3017–3027. [Google Scholar] [CrossRef]
- Böhm, M.; Grässel, S. Role of proopiomelanocortin-derived peptides and their receptors in the osteoarticular system: From basic to translational research. Endocr. Rev. 2012, 33, 623–651. [Google Scholar] [CrossRef] [Green Version]
- Roselli-Rehfuss, L.; Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Low, M.J.; Tatro, J.B.; Entwistle, M.L.; Simerly, R.B.; Cone, R.D. Identification of a receptor for gamma melanotropin and other proopiomelanocortin peptides in the hypothalamus and limbic system. Proc. Natl. Acad. Sci. USA 1993, 90, 8856–8860. [Google Scholar] [CrossRef] [Green Version]
- Pei, H.; Patterson, C.M.; Sutton, A.K.; Burnett, K.H.; Myers, M.G., Jr.; Olson, D.P. Lateral Hypothalamic Mc3R-Expressing Neurons Modulate Locomotor Activity, Energy Expenditure, and Adiposity in Male Mice. Endocrinology 2019, 160, 343–358. [Google Scholar] [CrossRef] [Green Version]
- Butler, A.A.; Girardet, C.; Mavrikaki, M.; Trevaskis, J.L.; Macarthur, H.; Marks, D.L.; Farr, S.A. A Life without Hunger: The Ups (and Downs) to Modulating Melanocortin-3 Receptor Signaling. Front. Neurosci. 2017, 11, 128. [Google Scholar] [CrossRef] [Green Version]
- Eerola, K.; Virtanen, S.; Vähätalo, L.; Ailanen, L.; Cai, M.; Hruby, V.; Savontaus, M.; Savontaus, E. Hypothalamic γ-melanocyte stimulating hormone gene delivery reduces fat mass in male mice. J. Endocrinol. 2018, 239, 19–31. [Google Scholar] [CrossRef]
- Getting, S.J.; Riffo-Vasquez, Y.; Pitchford, S.; Kaneva, M.; Grieco, P.; Page, C.P.; Perretti, M.; Spina, D. A role for MC3R in modulating lung inflammation. Pulm. Pharmacol. Ther. 2008, 21, 866–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montero-Melendez, T.; Forfar, R.A.; Cook, J.M.; Jerman, J.C.; Taylor, D.L.; Perretti, M. Old drugs with new skills: Fenoprofen as an allosteric enhancer at melanocortin receptor 3. Cell Mol. Life Sci. 2017, 74, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Juárez-Rebollar, D.; Rios, C.; Nava-Ruíz, C.; Méndez-Armenta, M. Metallothionein in Brain Disorders. Oxidative Med. Cell. Longev. 2017, 2017, 5828056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristensen, J.; Jonassen, T.E.; Rehling, M.; Tønnesen, E.; Sloth, E.; Nielsen, S.; Frøkiaer, J. The α-MSH analogue AP214 attenuates rise in pulmonary pressure and fall in ejection fraction in lipopolysaccharide-induced systemic inflammatory response syndrome in pigs. Clin. Physiol. Funct. Imaging 2011, 31, 54–60. [Google Scholar] [CrossRef]
- Montero-Melendez, T.; Patel, H.B.; Seed, M.; Nielsen, S.; Jonassen, T.E.; Perretti, M. The melanocortin agonist AP214 exerts anti-inflammatory and proresolving properties. Am. J. Pathol. 2011, 179, 259–269. [Google Scholar] [CrossRef]
- Montero-Melendez, T.; Gobbetti, T.; Cooray, S.N.; Jonassen, T.E.; Perretti, M. Biased agonism as a novel strategy to harness the proresolving properties of melanocortin receptors without eliciting melanogenic effects. J. Immunol. 2015, 194, 3381–3388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Getting, S.J.; Gibbs, L.; Clark, A.J.; Flower, R.J.; Perretti, M. POMC gene-derived peptides activate melanocortin type 3 receptor on murine macrophages, suppress cytokine release, and inhibit neutrophil migration in acute experimental inflammation. J. Immunol. 1999, 162, 7446–7453. [Google Scholar]
- King, S.H.; Mayorov, A.V.; Balse-Srinivasan, P.; Hruby, V.J.; Vanderah, T.W.; Wessells, H. Melanocortin receptors, melanotropic peptides and penile erection. Curr. Top. Med. Chem. 2007, 7, 1098–1106. [Google Scholar]
- Gantz, I.; Miwa, H.; Konda, Y.; Shimoto, Y.; Tashiro, T.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning, expression, and gene localization of a fourth melanocortin receptor. J. Biol. Chem. 1993, 268, 15174–15179. [Google Scholar] [CrossRef]
- Gantz, I.; Fong, T.M. The melanocortin system. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E468–E474. [Google Scholar] [CrossRef]
- Méquinion, M.; Foldi, C.J.; Andrews, Z.B. The Ghrelin-AgRP Neuron Nexus in Anorexia Nervosa: Implications for Metabolic and Behavioral Adaptations. Front. Nutr. 2019, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Morgan, D.A.; Cui, H.; Rahmouni, K. Activation of hypothalamic AgRP and POMC neurons evokes disparate sympathetic and cardiovascular responses. Am. J. Physiol. Circ. Physiol. 2020, 319, H1069–H1077. [Google Scholar] [CrossRef] [PubMed]
- Kishi, T.; Aschkenasi, C.J.; Lee, C.E.; Mountjoy, K.G.; Saper, C.B.; Elmquist, J.K. Expression of melanocortin 4 receptor mRNA in the central nervous system of the rat. J. Comp. Neurol. 2003, 457, 213–235. [Google Scholar] [CrossRef] [PubMed]
- Michael, N.J.; Caron, A.; Lee, C.E.; Castorena, C.M.; Lee, S.; Zigman, J.M.; Williams, K.W.; Elmquist, J.K. Melanocortin regulation of histaminergic neurons via perifornical lateral hypothalamic melanocortin 4 receptors. Mol. Metab. 2020, 35, 100956. [Google Scholar] [CrossRef]
- Demont, E.H.; Bailey, J.M.; Bit, R.A.; Brown, J.A.; Campbell, C.A.; Deeks, N.; Dowell, S.J.; Eldred, C.; Gaskin, P.; Gray, J.R.; et al. Discovery of Tetrahydropyrazolopyridine as Sphingosine 1-Phosphate Receptor 3 (S1P3)-Sparing S1P1 Agonists Active at Low Oral Doses. J. Med. Chem. 2016, 59, 1003–1020. [Google Scholar] [CrossRef]
- Muceniece, R.; Zvejniece, L.; Vilskersts, R.; Liepinsh, E.; Baumane, L.; Kalvinsh, I.; Wikberg, J.E.; Dambrova, M. Functional evaluation of THIQ, a melanocortin 4 receptor agonist, in models of food intake and inflammation. Basic Clin. Pharmacol. Toxicol. 2007, 101, 416–420. [Google Scholar] [CrossRef]
- Wessells, H.; Gralnek, D.; Dorr, R.; Hruby, V.J.; Hadley, M.E.; Levine, N. Effect of an alpha-melanocyte stimulating hormone analog on penile erection and sexual desire in men with organic erectile dysfunction. Urology 2000, 56, 641–646. [Google Scholar] [CrossRef]
- Molinoff, P.B.; Shadiack, A.M.; Earle, D.; Diamond, L.E.; Quon, C.Y. PT-141: A melanocortin agonist for the treatment of sexual dysfunction. Ann. N. Y. Acad. Sci. 2003, 994, 96–102. [Google Scholar] [CrossRef]
- Gantz, I.; Shimoto, Y.; Konda, Y.; Miwa, H.; Dickinson, C.J.; Yamada, T. Molecular cloning, expression, and characterization of a fifth melanocortin receptor. Biochem. Biophys. Res. Commun. 1994, 200, 1214–1220. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Guan, X.; Zhou, R.; Gong, R. Melanocortin 5 receptor signaling pathway in health and disease. Cell Mol. Life Sci. 2020, 77, 3831–3840. [Google Scholar] [CrossRef]
- Buggy, J.J. Binding of alpha-melanocyte-stimulating hormone to its G-protein-coupled receptor on B-lymphocytes activates the Jak/STAT pathway. Biochem. J. 1998, 331, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.; Namba, K. In vitro induction of CD25+ CD4+ regulatory T cells by the neuropeptide alpha-melanocyte stimulating hormone (alpha-MSH). Immunol. Cell Biol. 2001, 79, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Trotta, M.C.; Maisto, R.; Alessio, N.; Hermenean, A.; D’Amico, M.; Di Filippo, C. The Melanocortin MC5R as a New Target for Treatment of High Glucose-Induced Hypertrophy of the Cardiac H9c2 Cells. Front. Physiol. 2018, 9, 1475. [Google Scholar] [CrossRef] [PubMed]
- Glyn, J.R.; Lipton, J.M. Hypothermic and antipyretic effects of centrally administered ACTH (1--24) and alpha-melanotropin. Peptides 1981, 2, 177–187. [Google Scholar] [CrossRef]
- Kandasamy, S.B.; Williams, B.A. Hypothermic and antipyretic effects of ACTH (1-24) and alpha-melanotropin in guinea-pigs. Neuropharmacology 1984, 23, 49–53. [Google Scholar] [CrossRef]
- Shih, S.T.; Lipton, J.M. Intravenous alpha-MSH reduces fever in the squirrel monkey. Peptides 1985, 6, 685–687. [Google Scholar] [CrossRef]
- Bitto, A.; Polito, F.; Altavilla, D.; Irrera, N.; Giuliani, D.; Ottani, A.; Minutoli, L.; Spaccapelo, L.; Galantucci, M.; Lodi, R.; et al. Melanocortins protect against multiple organ dysfunction syndrome in mice. Br. J. Pharmacol. 2011, 162, 917–928. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, D.; Mioni, C.; Bazzani, C.; Zaffe, D.; Botticelli, A.R.; Capolongo, S.; Sabba, A.; Galantucci, M.; Iannone, A.; Grieco, P.; et al. Selective melanocortin MC4 receptor agonists reverse haemorrhagic shock and prevent multiple organ damage. Br. J. Pharmacol. 2007, 150, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Ottani, A.; Neri, L.; Canalini, F.; Calevro, A.; Rossi, R.; Cappelli, G.; Ballestri, M.; Giuliani, D.; Guarini, S. Protective effects of the melanocortin analog NDP-α-MSH in rats undergoing cardiac arrest. Eur. J. Pharmacol. 2014, 745, 108–116. [Google Scholar] [CrossRef]
- Vecsernyes, M.; Szokol, M.; Bombicz, M.; Priksz, D.; Gesztelyi, R.; Fulop, G.A.; Varga, B.; Juhasz, B.; Haines, D.; Tosaki, A. Alpha-Melanocyte-stimulating Hormone Induces Vasodilation and Exerts Cardioprotection Through the Heme-Oxygenase Pathway in Rat Hearts. J. Cardiovasc. Pharmacol. 2017, 69, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Qiang, X.; Liotta, A.S.; Shiloach, J.; Gutierrez, J.C.; Wang, H.; Ochani, M.; Ochani, K.; Yang, H.; Rabin, A.; LeRoith, D.; et al. New melanocortin-like peptide of E. coli can suppress inflammation via the mammalian melanocortin-1 receptor (MC1R): Possible endocrine-like function for microbes of the gut. NPJ Biofilms Microbiomes 2017, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Dong, B.; Li, C.; Zhang, F.; Ji, Y.; Zhang, J.; Yin, C. Dynamic changes of α-melanocyte-stimulating hormone levels in the serum of patients with craniocerebral trauma. Exp. Ther. Med. 2017, 14, 2511–2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patruno, S.; Garrido-Mesa, J.; Romano, M.; Perretti, M.; Montero-Melendez, T. Ligand Bias and Its Association With Pro-resolving Actions of Melanocortin Drugs. Front. Pharmacol. 2018, 9, 919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipton, J.M.; Ceriani, G.; Macaluso, A.; McCoy, D.; Carnes, K.; Biltz, J.; Catania, A. Antiinflammatory effects of the neuropeptide alpha-MSH in acute, chronic, and systemic inflammation. Ann. N. Y. Acad. Sci. 1994, 741, 137–148. [Google Scholar] [CrossRef]
- Deng, J.; Hu, X.; Yuen, P.S.; Star, R.A. Alpha-melanocyte-stimulating hormone inhibits lung injury after renal ischemia/reperfusion. Am. J. Respir. Crit. Care Med. 2004, 169, 749–756. [Google Scholar] [CrossRef]
- Miao, Y.L.; Deng, X.M.; Li, J.B.; Zhao, X.K.; Cao, J.M.; Tian, Y.P. Effect of alpha-melanocyte stimulating hormone on the apoptosis of the vascular endothelial cell of the lung in two-hit acute respiratory distress syndrome in rat. Zhongguo Wei Zhong Bing Ji Jiu Yi Xue 2004, 16, 596–598. [Google Scholar]
- Colombo, G.; Gatti, S.; Sordi, A.; Turcatti, F.; Carlin, A.; Rossi, C.; Lonati, C.; Catania, A. Production and effects of alpha-melanocyte-stimulating hormone during acute lung injury. Shock 2007, 27, 326–333. [Google Scholar] [CrossRef]
- Raap, U.; Brzoska, T.; Sohl, S.; Päth, G.; Emmel, J.; Herz, U.; Braun, A.; Luger, T.; Renz, H. Alpha-melanocyte-stimulating hormone inhibits allergic airway inflammation. J. Immunol. 2003, 171, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Webering, S.; Lunding, L.P.; Vock, C.; Schröder, A.; Gaede, K.I.; Herzmann, C.; Fehrenbach, H.; Wegmann, M. The alpha-melanocyte-stimulating hormone acts as a local immune homeostasis factor in experimental allergic asthma. Clin. Exp. Allergy 2019, 49, 1026–1039. [Google Scholar] [CrossRef]
- Böhm, M.; Luger, T. Are melanocortin peptides future therapeutics for cutaneous wound healing? Exp. Dermatol. 2019, 28, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Land, S.C. Inhibition of cellular and systemic inflammation cues in human bronchial epithelial cells by melanocortin-related peptides: Mechanism of KPV action and a role for MC3R agonists. Int. J. Physiol. Pathophysiol. Pharmacol. 2012, 4, 59–73. [Google Scholar] [PubMed]
- Clemson, C.M.; Yost, J.; Taylor, A.W. The Role of Alpha-MSH as a Modulator of Ocular Immunobiology Exemplifies Mechanistic Differences between Melanocortins and Steroids. Ocul. Immunol. Inflamm. 2017, 25, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.J.; Biros, D.J.; Taylor, A.W. Injection of an alpha-melanocyte stimulating hormone expression plasmid is effective in suppressing experimental autoimmune uveitis. Int. Immunopharmacol. 2009, 9, 1079–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.W.; Ng, T.F. Negative regulators that mediate ocular immune privilege. J. Leukoc. Biol. 2018, 103, 1179–1187. [Google Scholar] [CrossRef]
- Lee, D.J.; Taylor, A.W. Both MC5r and A2Ar are required for protective regulatory immunity in the spleen of post-experimental autoimmune uveitis in mice. J. Immunol. 2013, 191, 4103–4111. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Yang, Q.; Hou, M.; Han, Q.; Zhang, H.; Wang, J.; Qi, C.; Bo, Q.; Ru, Y.; Yang, W.; et al. A-Melanocyte-Stimulating Hormone Protects Early Diabetic Retina from Blood-Retinal Barrier Breakdown and Vascular Leakage via MC4R. Cell Physiol. Biochem. 2018, 45, 505–522. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, L.; Liu, X.; Jiang, Y.; Zhang, L.; Zhang, X.; Li, X.; Zhang, Y. α-Melanocyte-stimulating hormone protects retinal vascular endothelial cells from oxidative stress and apoptosis in a rat model of diabetes. PLoS ONE 2014, 9, e93433. [Google Scholar] [CrossRef]
- Zhang, Z.; Yao, K.; Ma, J.; Tang, X. Anti-inflammatory effect of the alpha-melanocyte stimulating hormone in animal eyes undergoing extracapsular lens extraction. Mol. Biol. 2011, 45, 272–281. [Google Scholar] [CrossRef]
- Laroui, H.; Dalmasso, G.; Nguyen, H.T.; Yan, Y.; Sitaraman, S.V.; Merlin, D. Drug-loaded nanoparticles targeted to the colon with polysaccharide hydrogel reduce colitis in a mouse model. Gastroenterology 2010, 138, 843–853.e2. [Google Scholar] [CrossRef]
- Zeng, M.; Shao, A.; Li, H.; Tang, Y.; Li, Q.; Guo, Z.; Wu, C.; Cheng, Y.; Tian, H.; Zhu, W.H. Peptide Receptor-Targeted Fluorescent Probe: Visualization and Discrimination between Chronic and Acute Ulcerative Colitis. ACS Appl. Mater. Interfaces 2017, 9, 13029–13036. [Google Scholar] [CrossRef]
- Váradi, J.; Harazin, A.; Fenyvesi, F.; Réti-Nagy, K.; Gogolák, P.; Vámosi, G.; Bácskay, I.; Fehér, P.; Ujhelyi, Z.; Vasvári, G.; et al. Alpha-Melanocyte Stimulating Hormone Protects against Cytokine-Induced Barrier Damage in Caco-2 Intestinal Epithelial Monolayers. PLoS ONE 2017, 12, e0170537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, P.; Yang, Y.; Liu, Z.; Huang, J.; Gong, Y.; Sun, H. Oral Bifidobacterium longum expressing alpha-melanocyte-stimulating hormone to fight experimental colitis. Drug Deliv. 2016, 23, 2058–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmasso, G.; Charrier-Hisamuddin, L.; Nguyen, H.T.; Yan, Y.; Sitaraman, S.; Merlin, D. PepT1-mediated tripeptide KPV uptake reduces intestinal inflammation. Gastroenterology 2008, 134, 166–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannengiesser, K.; Maaser, C.; Heidemann, J.; Luegering, A.; Ross, M.; Brzoska, T.; Bohm, M.; Luger, T.A.; Domschke, W.; Kucharzik, T. Melanocortin-derived tripeptide KPV has anti-inflammatory potential in murine models of inflammatory bowel disease. Inflamm. Bowel Dis. 2008, 14, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Xu, Z.; Viennois, E.; Zhang, Y.; Zhang, Z.; Zhang, M.; Han, M.K.; Kang, Y.; Merlin, D. Orally Targeted Delivery of Tripeptide KPV via Hyaluronic Acid-Functionalized Nanoparticles Efficiently Alleviates Ulcerative Colitis. Mol. Ther. 2017, 25, 1628–1640. [Google Scholar] [CrossRef] [Green Version]
- Peñate-Medina, T.; Damoah, C.; Benezra, M.; Will, O.; Kairemo, K.; Humbert, J.; Sebens, S.; Peñate-Medina, O. Alpha-MSH Targeted Liposomal Nanoparticle for Imaging in Inflammatory Bowel Disease (IBD). Curr. Pharm. Des. 2020, 26, 3840–3846. [Google Scholar] [CrossRef]
- Min, Y.; Han, D.; Fu, Z.; Wang, H.; Liu, L.; Tian, Y. α-MSH inhibits TNF-α-induced maturation of human dendritic cells in vitro through the up-regulation of ANXA1. Acta Biochim. Biophys. Sin. 2011, 43, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Flores-Bastías, O.; Adriasola-Carrasco, A.; Karahanian, E. Activation of Melanocortin-4 Receptor Inhibits Both Neuroinflammation Induced by Early Exposure to Ethanol and Subsequent Voluntary Alcohol Intake in Adulthood in Animal Models: Is BDNF the Key Mediator? Front. Cell. Neurosci. 2020, 14, 5. [Google Scholar] [CrossRef]
- Flores-Bastías, O.; Gómez, G.I.; Orellana, J.A.; Karahanian, E. Activation of Melanocortin-4 Receptor by a Synthetic Agonist Inhibits Ethanolinduced Neuroinflammation in Rats. Curr. Pharm. Des. 2019, 25, 4799–4805. [Google Scholar] [CrossRef]
- Wu, X.; Fu, S.; Liu, Y.; Luo, H.; Li, F.; Wang, Y.; Gao, M.; Cheng, Y.; Xie, Z. NDP-MSH binding melanocortin-1 receptor ameliorates neuroinflammation and BBB disruption through CREB/Nr4a1/NF-κB pathway after intracerebral hemorrhage in mice. J. Neuroinflam. 2019, 16, 192. [Google Scholar] [CrossRef]
- Mykicki, N.; Herrmann, A.M.; Schwab, N.; Deenen, R.; Sparwasser, T.; Limmer, A.; Wachsmuth, L.; Klotz, L.; Köhrer, K.; Faber, C.; et al. Melanocortin-1 receptor activation is neuroprotective in mouse models of neuroinflammatory disease. Sci. Transl. Med. 2016, 8, 362ra146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carniglia, L.; Durand, D.; Caruso, C.; Lasaga, M. Effect of NDP-α-MSH on PPAR-γ and -β expression and anti-inflammatory cytokine release in rat astrocytes and microglia. PLoS ONE 2013, 8, e57313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, G.; Feng, J.; Zhang, S.; Tong, Y.; Zhang, Q.; Yang, X.; Zhang, H. Altered levels of α-melanocyte stimulating hormone in cerebrospinal fluid and plasma of patients with traumatic brain injury. Brain Res. 2018, 1696, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Kamermans, A.; Verhoeven, T.; van Het Hof, B.; Koning, J.J.; Borghuis, L.; Witte, M.; van Horssen, J.; de Vries, H.E.; Rijnsburger, M. Setmelanotide, a Novel, Selective Melanocortin Receptor-4 Agonist Exerts Anti-inflammatory Actions in Astrocytes and Promotes an Anti-inflammatory Macrophage Phenotype. Front. Immunol. 2019, 10, 2312. [Google Scholar] [CrossRef]
- Chen, S.; Zhao, L.; Sherchan, P.; Ding, Y.; Yu, J.; Nowrangi, D.; Tang, J.; Xia, Y.; Zhang, J.H. Activation of melanocortin receptor 4 with RO27-3225 attenuates neuroinflammation through AMPK/JNK/p38 MAPK pathway after intracerebral hemorrhage in mice. J. Neuroinflamm. 2018, 15, 106. [Google Scholar] [CrossRef]
- Chen, S.; Zuo, Y.; Huang, L.; Sherchan, P.; Zhang, J.; Yu, Z.; Peng, J.; Zhang, J.; Zhao, L.; Doycheva, D.; et al. The MC(4) receptor agonist RO27-3225 inhibits NLRP1-dependent neuronal pyroptosis via the ASK1/JNK/p38 MAPK pathway in a mouse model of intracerebral haemorrhage. Br. J. Pharmacol. 2019, 176, 1341–1356. [Google Scholar] [CrossRef]
- Kleiner, S.; Braunstahl, G.J.; Rüdrich, U.; Gehring, M.; Eiz-Vesper, B.; Luger, T.A.; Steelant, B.; Seys, S.F.; Kapp, A.; Böhm, M.; et al. Regulation of melanocortin 1 receptor in allergic rhinitis in vitro and in vivo. Clin. Exp. Allergy 2016, 46, 1066–1074. [Google Scholar] [CrossRef]
- Etori, M.; Yonekubo, K.; Sato, E.; Mizukami, K.; Hirahara, K.; Karasuyama, H.; Maeda, H.; Yamashita, M. Melanocortin receptors 1 and 5 might mediate inhibitory effects of α-melanocyte-stimulating hormone on antigen-induced chronic allergic skin inflammation in IgE transgenic mice. J. Investig. Dermatol. 2012, 132, 1925–1927. [Google Scholar] [CrossRef] [Green Version]
- Andoh, T.; Akasaka, C.; Shimizu, K.; Lee, J.B.; Yoshihisa, Y.; Shimizu, T. Involvement of α-Melanocyte-Stimulating Hormone-Thromboxane A(2) System on Itching in Atopic Dermatitis. Am. J. Pathol. 2019, 189, 1775–1785. [Google Scholar] [CrossRef]
- Mehta, D.; Granstein, R.D. Immunoregulatory Effects of Neuropeptides on Endothelial Cells: Relevance to Dermatological Disorders. Dermatology 2019, 235, 175–186. [Google Scholar] [CrossRef]
- Capsoni, F.; Ongari, A.M.; Lonati, C.; Accetta, R.; Gatti, S.; Catania, A. α-Melanocyte-stimulating-hormone (α-MSH) modulates human chondrocyte activation induced by proinflammatory cytokines. BMC Musculoskelet. Disord. 2015, 16, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montero-Melendez, T.; Nagano, A.; Chelala, C.; Filer, A.; Buckley, C.D.; Perretti, M. Therapeutic senescence via GPCR activation in synovial fibroblasts facilitates resolution of arthritis. Nat. Commun. 2020, 11, 745. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yao, Z.; He, M.; Tian, C. Synovial Fluid α-MSH Levels are Inversely Correlated with Articular Cartilage Degeneration in Anterior Cruciate Ligament Deficient Knees. Clin. Lab. 2015, 61, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Li, Y.; Ding, L.B.; Liu, G.Y.; Zheng, X.F.; Xue, W.; Wang, H.J. Synovial fluid alpha-melanocyte-stimulating hormone may act as a protective biomarker for primary knee osteoarthritis. Discov. Med. 2019, 27, 17–26. [Google Scholar]
- Böhm, M.; Apel, M.; Lowin, T.; Lorenz, J.; Jenei-Lanzl, Z.; Capellino, S.; Dosoki, H.; Luger, T.A.; Straub, R.H.; Grässel, S. α-MSH modulates cell adhesion and inflammatory responses of synovial fibroblasts from osteoarthritis patients. Biochem. Pharmacol. 2016, 116, 89–99. [Google Scholar] [CrossRef]
- Liu, G.; Chen, Y.; Wang, G.; Niu, J. Decreased Synovial Fluid α-Melanocyte-Stimulating-Hormone (α-MSH) Levels Reflect Disease Severity in Patients with Posttraumatic Ankle Osteoarthritis. Clin. Lab. 2016, 62, 1491–1500. [Google Scholar] [CrossRef]
- Zaidi, M.; Sun, L.; Robinson, L.J.; Tourkova, I.L.; Liu, L.; Wang, Y.; Zhu, L.L.; Liu, X.; Li, J.; Peng, Y.; et al. ACTH protects against glucocorticoid-induced osteonecrosis of bone. Proc. Natl. Acad. Sci. USA 2010, 107, 8782–8787. [Google Scholar] [CrossRef] [Green Version]
- Mao, Z.; Liu, G.; Chen, J.J.; Liu, D.; Xu, M.P.; Zhao, C.; Yang, H.T.; Yue, Y.B. Serum α-melanocyte-stimulating hormone may act as a protective biomarker for non-traumatic osteonecrosis of the femoral head. Ann. Clin. Biochem. 2018, 55, 453–460. [Google Scholar] [CrossRef]
- Holloway, P.M.; Durrenberger, P.F.; Trutschl, M.; Cvek, U.; Cooper, D.; Orr, A.W.; Perretti, M.; Getting, S.J.; Gavins, F.N. Both MC1 and MC3 Receptors Provide Protection From Cerebral Ischemia-Reperfusion-Induced Neutrophil Recruitment. Arter. Thromb. Vasc. Biol. 2015, 35, 1936–1944. [Google Scholar] [CrossRef] [Green Version]
- Ottani, A.; Giuliani, D.; Neri, L.; Calevro, A.; Canalini, F.; Vandini, E.; Cainazzo, M.M.; Ruberto, I.A.; Barbieri, A.; Rossi, R.; et al. NDP-α-MSH attenuates heart and liver responses to myocardial reperfusion via the vagus nerve and JAK/ERK/STAT signaling. Eur. J. Pharmacol. 2015, 769, 22–32. [Google Scholar] [CrossRef]
- Hassoun, H.T.; Zou, L.; Moore, F.A.; Kozar, R.A.; Weisbrodt, N.W.; Kone, B.C. Alpha-melanocyte-stimulating hormone protects against mesenteric ischemia-reperfusion injury. Am. J. Physiol. Liver Physiol. 2002, 282, G1059–G1068. [Google Scholar] [CrossRef]
- Minutoli, L.; Bitto, A.; Squadrito, F.; Irrera, N.; Rinaldi, M.; Nicotina, P.A.; Arena, S.; Magno, C.; Marini, H.; Spaccapelo, L.; et al. Melanocortin 4 receptor activation protects against testicular ischemia-reperfusion injury by triggering the cholinergic antiinflammatory pathway. Endocrinology 2011, 152, 3852–3861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skott, M.; Nørregaard, R.; Birke-Sørensen, H.; Palmfeldt, J.; Kwon, T.H.; Jonassen, T.; Frøkiær, J.; Nielsen, S. Development of intestinal ischemia/reperfusion-induced acute kidney injury in rats with or without chronic kidney disease: Cytokine/chemokine response and effect of α-melanocyte-stimulating hormone. Kidney Res. Clin. Pract. 2014, 33, 79–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuutinen, S.; Ailanen, L.; Savontaus, E.; Rinne, P. Melanocortin overexpression limits diet-induced inflammation and atherosclerosis in LDLR(-/-) mice. J. Endocrinol. 2018, 236, 111–123. [Google Scholar] [CrossRef]
- Le Saux, G.; Plawinski, L.; Nlate, S.; Ripoche, J.; Buffeteau, T.; Durrieu, M.C. Beneficial Effect of Covalently Grafted α-MSH on Endothelial Release of Inflammatory Mediators for Applications in Implantable Devices. PLoS ONE 2016, 11, e0150706. [Google Scholar] [CrossRef]
- Gatti, S.; Colombo, G.; Buffa, R.; Turcatti, F.; Garofalo, L.; Carboni, N.; Ferla, L.; Fassati, L.R.; Lipton, J.M.; Catania, A. alpha-Melanocyte-stimulating hormone protects the allograft in experimental heart transplantation. Transplantation 2002, 74, 1678–1684. [Google Scholar] [CrossRef]
- Liu, G.; Li, M.; Saeed, M.; Xu, Y.; Ren, Q.; Sun, C. αMSH inhibits adipose inflammation via reducing FoxOs transcription and blocking Akt/JNK pathway in mice. Oncotarget 2017, 8, 47642–47654. [Google Scholar] [CrossRef] [Green Version]
- Manna, S.K.; Aggarwal, B.B. Alpha-melanocyte-stimulating hormone inhibits the nuclear transcription factor NF-kappa B activation induced by various inflammatory agents. J. Immunol. 1998, 161, 2873–2880. [Google Scholar]
- Gerlo, S.; Kooijman, R.; Beck, I.M.; Kolmus, K.; Spooren, A.; Haegeman, G. Cyclic AMP: A selective modulator of NF-κB action. Cell Mol. Life Sci. 2011, 68, 3823–3841. [Google Scholar] [CrossRef]
- Mandrika, I.; Muceniece, R.; Wikberg, J.E. Effects of melanocortin peptides on lipopolysaccharide/interferon-gamma-induced NF-kappaB DNA binding and nitric oxide production in macrophage-like RAW 264.7 cells: Evidence for dual mechanisms of action. Biochem. Pharmacol. 2001, 61, 613–621. [Google Scholar] [CrossRef]
- Delgado, R.; Carlin, A.; Airaghi, L.; Demitri, M.T.; Meda, L.; Galimberti, D.; Baron, P.; Lipton, J.M.; Catania, A. Melanocortin peptides inhibit production of proinflammatory cytokines and nitric oxide by activated microglia. J. Leukoc. Biol. 1998, 63, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Galimberti, D.; Baron, P.; Meda, L.; Prat, E.; Scarpini, E.; Delgado, R.; Catania, A.; Lipton, J.M.; Scarlato, G. Alpha-MSH peptides inhibit production of nitric oxide and tumor necrosis factor-alpha by microglial cells activated with beta-amyloid and interferon gamma. Biochem. Biophys. Res. Commun. 1999, 263, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Mukaida, N.; Okamoto, S.; Ishikawa, Y.; Matsushima, K. Molecular mechanism of interleukin-8 gene expression. J. Leukoc. Biol. 1994, 56, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Böhm, M.; Schulte, U.; Kalden, H.; Luger, T.A. Alpha-melanocyte-stimulating hormone modulates activation of NF-kappa B and AP-1 and secretion of interleukin-8 in human dermal fibroblasts. Ann. N. Y. Acad. Sci. 1999, 885, 277–286. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The role of the transcription factor CREB in immune function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef] [Green Version]
- Kalden, D.H.; Scholzen, T.; Brzoska, T.; Luger, T.A. Mechanisms of the antiinflammatory effects of alpha-MSH. Role of transcription factor NF-kappa B and adhesion molecule expression. Ann. N. Y. Acad. Sci. 1999, 885, 254–261. [Google Scholar] [CrossRef]
- Scholzen, T.E.; Sunderkötter, C.; Kalden, D.H.; Brzoska, T.; Fastrich, M.; Fisbeck, T.; Armstrong, C.A.; Ansel, J.C.; Luger, T.A. Alpha-melanocyte stimulating hormone prevents lipopolysaccharide-induced vasculitis by down-regulating endothelial cell adhesion molecule expression. Endocrinology 2003, 144, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Becher, E.; Mahnke, K.; Brzoska, T.; Kalden, D.H.; Grabbe, S.; Luger, T.A. Human peripheral blood-derived dendritic cells express functional melanocortin receptor MC-1R. Ann. N. Y. Acad. Sci. 1999, 885, 188–195. [Google Scholar] [CrossRef]
- Bonfiglio, V.; Camillieri, G.; Avitabile, T.; Leggio, G.M.; Drago, F. Effects of the COOH-terminal tripeptide alpha-MSH(11-13) on corneal epithelial wound healing: Role of nitric oxide. Exp. Eye Res. 2006, 83, 1366–1372. [Google Scholar] [CrossRef] [PubMed]
- Star, R.A.; Rajora, N.; Huang, J.; Stock, R.C.; Catania, A.; Lipton, J.M. Evidence of autocrine modulation of macrophage nitric oxide synthase by alpha-melanocyte-stimulating hormone. Proc. Natl. Acad. Sci. USA 1995, 92, 8016–8020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajora, N.; Ceriani, G.; Catania, A.; Star, R.A.; Murphy, M.T.; Lipton, J.M. alpha-MSH production, receptors, and influence on neopterin in a human monocyte/macrophage cell line. J. Leukoc. Biol. 1996, 59, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.J.; Han, D.J.; Chang, S.H.; Lim, D.G.; Wee, Y.M.; Kim, J.H.; Kim, Y.H.; Koo, S.K.; Choi, M.; Kim, S.C. Protective effect of alpha-melanocyte-stimulating hormone on pancreas islet cell against peripheral blood mononuclear cell-mediated cytotoxicity in vitro. Transplant. Proc. 2007, 39, 1604–1606. [Google Scholar] [CrossRef] [PubMed]
- Oktar, B.K.; Yüksel, M.; Alican, I. The role of cyclooxygenase inhibition in the effect of alpha-melanocyte-stimulating hormone on reactive oxygen species production by rat peritoneal neutrophils. Prostaglandins Leukot. Essent. Fat. Acids 2004, 71, 1–5. [Google Scholar] [CrossRef]
- Taylor, A.W.; Lee, D.J. The alpha-melanocyte stimulating hormone induces conversion of effector T cells into treg cells. J. Transplant. 2011, 2011, 246856. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Han, D.; Hong, J.; Zhang, H.; Ying, Y.; Tian, Y.; Zhang, L.; Lin, J. SVα-MSH, a novel α-melanocyte stimulating hormone analog, ameliorates autoimmune encephalomyelitis through inhibiting autoreactive CD4(+) T cells activation. J. Neuroimmunol. 2014, 269, 9–19. [Google Scholar] [CrossRef]
- Namba, K.; Kitaichi, N.; Nishida, T.; Taylor, A.W. Induction of regulatory T cells by the immunomodulating cytokines alpha-melanocyte-stimulating hormone and transforming growth factor-beta2. J. Leukoc. Biol. 2002, 72, 946–952. [Google Scholar]
- Ng, T.F.; Kitaichi, N.; Taylor, A.W. In vitro generated autoimmune regulatory T cells enhance intravitreous allogeneic retinal graft survival. Investig. Opthalmol. Vis. Sci. 2007, 48, 5112–5117. [Google Scholar] [CrossRef]
- Nishida, T.; Miyata, S.; Itoh, Y.; Mizuki, N.; Ohgami, K.; Shiratori, K.; Ilieva, I.B.; Ohno, S.; Taylor, A.W. Anti-inflammatory effects of alpha-melanocyte-stimulating hormone against rat endotoxin-induced uveitis and the time course of inflammatory agents in aqueous humor. Int. Immunopharmacol. 2004, 4, 1059–1066. [Google Scholar] [CrossRef]
- Boada-Romero, E.; Martinez, J.; Heckmann, B.L.; Green, D.R. The clearance of dead cells by efferocytosis. Nat. Rev. Mol. Cell Biol. 2020, 21, 398–414. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Kim, T.J.; Peng, D.H.; Duan, D.; Gibbons, D.L.; Yamauchi, M.; Jackson, J.R.; Le Saux, C.J.; Calhoun, C.; Peters, J.; et al. Fibroblast-specific inhibition of TGF-β1 signaling attenuates lung and tumor fibrosis. J. Clin. Investig. 2017, 127, 3675–3688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thannickal, V.J.; Lee, D.Y.; White, E.S.; Cui, Z.; Larios, J.M.; Chacon, R.; Horowitz, J.C.; Day, R.M.; Thomas, P.E. Myofibroblast differentiation by transforming growth factor-beta1 is dependent on cell adhesion and integrin signaling via focal adhesion kinase. J. Biol. Chem. 2003, 278, 12384–12389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhm, M.; Raghunath, M.; Sunderkötter, C.; Schiller, M.; Ständer, S.; Brzoska, T.; Cauvet, T.; Schiöth, H.B.; Schwarz, T.; Luger, T.A. Collagen metabolism is a novel target of the neuropeptide alpha-melanocyte-stimulating hormone. J. Biol. Chem. 2004, 279, 6959–6966. [Google Scholar] [CrossRef] [Green Version]
- Hill, R.P.; Wheeler, P.; MacNeil, S.; Haycock, J.W. Alpha-melanocyte stimulating hormone cytoprotective biology in human dermal fibroblast cells. Peptides 2005, 26, 1150–1158. [Google Scholar] [CrossRef]
- Kokot, A.; Sindrilaru, A.; Schiller, M.; Sunderkötter, C.; Kerkhoff, C.; Eckes, B.; Scharffetter-Kochanek, K.; Luger, T.A.; Böhm, M. alpha-melanocyte-stimulating hormone suppresses bleomycin-induced collagen synthesis and reduces tissue fibrosis in a mouse model of scleroderma: Melanocortin peptides as a novel treatment strategy for scleroderma? Arthritis Rheum. 2009, 60, 592–603. [Google Scholar] [CrossRef]
- Böhm, M.; Stegemann, A. Bleomycin-induced fibrosis in MC1 signalling-deficient C57BL/6J-Mc1r(e/e) mice further supports a modulating role for melanocortins in collagen synthesis of the skin. Exp. Dermatol. 2014, 23, 431–433. [Google Scholar] [CrossRef]
- Luo, L.F.; Shi, Y.; Zhou, Q.; Xu, S.Z.; Lei, T.C. Insufficient expression of the melanocortin-1 receptor by human dermal fibroblasts contributes to excess collagen synthesis in keloid scars. Exp. Dermatol. 2013, 22, 764–766. [Google Scholar] [CrossRef]
- De Souza, K.S.; Cantaruti, T.A.; Azevedo, G.M., Jr.; Galdino, D.A.; Rodrigues, C.M.; Costa, R.A.; Vaz, N.M.; Carvalho, C.R. Improved cutaneous wound healing after intraperitoneal injection of alpha-melanocyte-stimulating hormone. Exp. Dermatol. 2015, 24, 198–203. [Google Scholar] [CrossRef]
- Lee, T.H.; Jawan, B.; Chou, W.Y.; Lu, C.N.; Wu, C.L.; Kuo, H.M.; Concejero, A.M.; Wang, C.H. Alpha-melanocyte-stimulating hormone gene therapy reverses carbon tetrachloride induced liver fibrosis in mice. J. Gene Med. 2006, 8, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Lee, T.H.; Lu, C.N.; Chou, W.Y.; Hung, K.S.; Concejero, A.M.; Jawan, B. Electroporative alpha-MSH gene transfer attenuates thioacetamide-induced murine hepatic fibrosis by MMP and TIMP modulation. Gene Ther. 2006, 13, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Lonati, C.; Carlin, A.; Leonardi, P.; Valenza, F.; Bosari, S.; Catania, A.; Gatti, S. Modulatory effects of NDP-MSH in the regenerating liver after partial hepatectomy in rats. Peptides 2013, 50, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Vándor, E.; Simon, G.; Anda, E.; Budavári, I. The effect of alpha-melanophor-stimulating hormone on liver regeneration and incorporation of amino acid in rats’ liver protein. Endokrinologie 1975, 66, 81–87. [Google Scholar] [PubMed]
- Xu, P.B.; Mao, Y.F.; Meng, H.B.; Tian, Y.P.; Deng, X.M. STY39, a novel alpha-melanocyte-stimulating hormone analogue, attenuates bleomycin-induced pulmonary inflammation and fibrosis in mice. Shock 2011, 35, 308–314. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jo, S.K.; Cho, W.Y.; Kim, H.K.; Won, N.H. The effect of alpha-melanocyte-stimulating hormone on renal tubular cell apoptosis and tubulointerstitial fibrosis in cyclosporine A nephrotoxicity. Transplantation 2004, 78, 1756–1764. [Google Scholar] [CrossRef]
- Verhaagen, J.; Edwards, P.M.; Jennekens, F.G.; Schotman, P.; Gispen, W.H. Alpha-melanocyte-stimulating hormone stimulates the outgrowth of myelinated nerve fibers after peripheral nerve crush. Exp. Neurol. 1986, 92, 451–454. [Google Scholar] [CrossRef]
- Dekker, A.J. Effect of alpha-melanocyte-stimulating hormone on peripheral nerve regeneration in the rat: Histological aspects and comparison with the effect of gangliosides. Exp. Neurol. 1988, 99, 490–497. [Google Scholar] [CrossRef]
- Laquerriere, A.; Peulve, P.; Jin, O.; Tiollier, J.; Tardy, M.; Vaudry, H.; Hemet, J.; Tadie, M. Effect of basic fibroblast growth factor and alpha-melanocytic stimulating hormone on nerve regeneration through a collagen channel. Microsurgery 1994, 15, 203–210. [Google Scholar] [CrossRef]
- Plantinga, L.C.; Verhaagen, J.; Edwards, P.M.; Hali, M.; Brakkee, J.H.; Gispen, W.H. Pharmacological evidence for the involvement of endogenous alpha-MSH-like peptides in peripheral nerve regeneration. Peptides 1995, 16, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Ter Laak, M.P.; Brakkee, J.H.; Adan, R.A.; Hamers, F.P.; Gispen, W.H. The potent melanocortin receptor agonist melanotan-II promotes peripheral nerve regeneration and has neuroprotective properties in the rat. Eur. J. Pharmacol. 2003, 462, 179–183. [Google Scholar] [CrossRef]
- Pavan, J.; Lukenda, A.; Stambuk, N.; Konjevoda, P.; Kastelan, S.; Curković, M. Effects of alpha-MSH on corneal epithelial lesions in rats. Coll. Antropol. 2012, 36, 1407–1411. [Google Scholar] [PubMed]
- Zhang, Z.; Ma, J.; Yao, K.; Yin, J. Alpha-melanocyte stimulating hormone suppresses the proliferation of human tenon’s capsule fibroblast proliferation induced by transforming growth factor beta 1. Mol. Biol. 2012, 46, 628–633. [Google Scholar] [CrossRef]
Figure 1.
Simplified role of MC1R signaling in anti-inflammatory actions of α-MSH. MC1R activates adenylyl cyclase and generates intracellular cAMP, which activates protein kinases (C and A). This leads to activation of MAPK and JAK and STAT pathways. In the nucleus, the catalytic subunits can phosphorylate different substrates, the best known of which is the transcription factor CREB. CREB is involved in transcription of anti-inflammatory mediators. Alternatively, protein kinase activation can lead to increased cytoplasmic inhibitor of κB (IκB) through blocking IκB kinase, inhibition of IκB ubiquitinylation, etc. This leads to inhibition of NF-κB and decreased expression of downstream proinflammatory genes.
Figure 1.
Simplified role of MC1R signaling in anti-inflammatory actions of α-MSH. MC1R activates adenylyl cyclase and generates intracellular cAMP, which activates protein kinases (C and A). This leads to activation of MAPK and JAK and STAT pathways. In the nucleus, the catalytic subunits can phosphorylate different substrates, the best known of which is the transcription factor CREB. CREB is involved in transcription of anti-inflammatory mediators. Alternatively, protein kinase activation can lead to increased cytoplasmic inhibitor of κB (IκB) through blocking IκB kinase, inhibition of IκB ubiquitinylation, etc. This leads to inhibition of NF-κB and decreased expression of downstream proinflammatory genes.
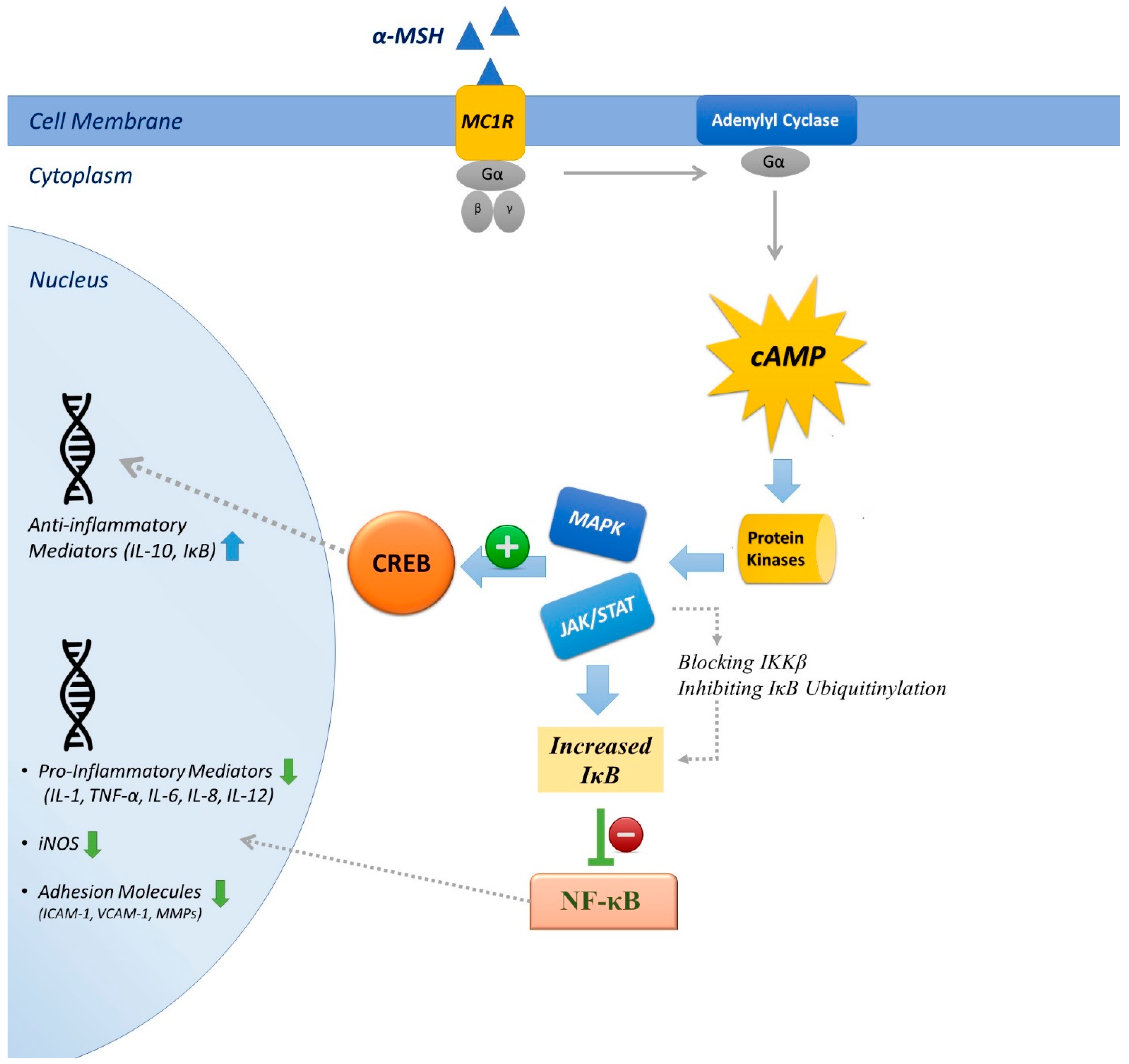
Figure 2.
Possible anti-fibrotic properties of α-MSH (see text).


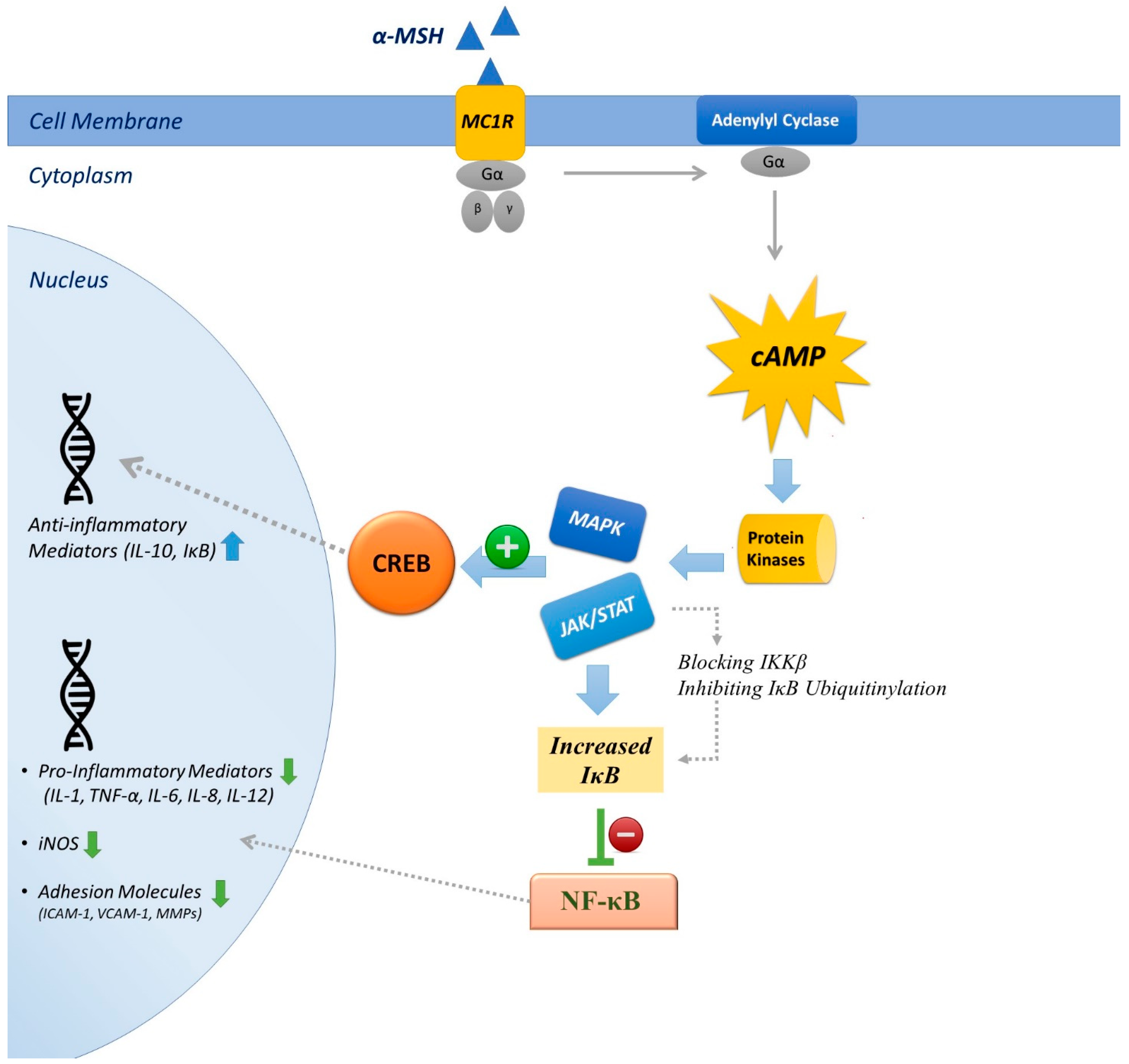{kind=link}
{kind=link}
Table 1.
Melanocortin receptors; tissue distribution, known agonists and antagonists, and their biological effects.
Table 1.
Melanocortin receptors; tissue distribution, known agonists and antagonists, and their biological effects.
| Receptor | Tissue Distribution | Species | Agonist | Biologic Effects | Antagonist | Biologic Effects |
|---|---|---|---|---|---|---|
| MC1R | Present in melanocytes/skin | Human [41,42] | α-MSH, ACTH, β-MSH, γ-MSH | Pigmentation, anti-inflammatory [43,44] | Agouti | Suppresses melanine production [45,46] |
| Present in grey matter | Rat [47] | NDP-MSH | Anti-inflammatory [48,49,50] | |||
| BMS-470539 | ||||||
| Present in monocyte, macrophage (including alveolar), lymphocyte, neutrophil | Human, murine [51,52] | AP-1189 | ||||
| MC2R | Present in adrenal cortex; absent in liver, lung, thyroid and kidney | Rhesus macaque [41] | ACTH | Induce steroidogenesis [18] | GPS1574 | Inhibits steroidogenesis [53,54] |
| Present in chondrocyte and osteoblast | Human [55,56] | |||||
| MC3R | Present in brain, placenta; absent in adrenal, kidney, liver | Rat [57] | γ-MSH ≥ ACTH = β-MSH = α-MSH | Energy equilibrium, cardiovascular [58,59,60] | SHU-9119 | Inhibits anti-inflammatory effects of γMSH [36] |
| Present in lung | Murine [61] | MTII | Anti-inflammatory [61,62,63,64,65,66] | |||
| D-Trp8-γMSH | ||||||
| Present in macrophage and monocytes | Murine [51,52,55,56,57,67] | AP-214 | AVM-127 | inhibits α-MSH-induced penile erection [68] | ||
| AP-1189 (biased agonist) | ||||||
| MC4R | Present in brain; absent in lung, liver, kidney, adrenal | Rat/Canine [69] | α-MSH = ACTH > β-MSH > γ-MSH | Energy balance, erectile function, cardiovascular effects [43,70] | AgRP | Inhibitory cardiovascular effects, increases food intake [71,72] |
| Present in thalamus and hypothalamus | Rat [73] | THIQ | Anti-inflammatory, inhibits food intake [74,75,76] | ML00253764 | ||
| Ro27-3225 | ||||||
| PT-141 | Induces erection [77,78] | |||||
| MC5R | Present in lung, skeletal muscle, brain; absent in adrenal | Murine/Human [79] | α-MSH, ACTH, β-MSH, | Anti-inflammatory [80] | ||
| Present in B-lymphocytes | Mouse [81] | |||||
| Present in T-lymphocytes | Mouse [82] | PG-901 | Inhibits glucose uptake [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dinparastisaleh, R.; Mirsaeidi, M. Antifibrotic and Anti-Inflammatory Actions of α-Melanocytic Hormone: New Roles for an Old Player. Pharmaceuticals 2021, 14, 45. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14010045
AMA Style
Dinparastisaleh R, Mirsaeidi M. Antifibrotic and Anti-Inflammatory Actions of α-Melanocytic Hormone: New Roles for an Old Player. Pharmaceuticals. 2021; 14(1):45. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14010045
Chicago/Turabian StyleDinparastisaleh, Roshan, and Mehdi Mirsaeidi. 2021. "Antifibrotic and Anti-Inflammatory Actions of α-Melanocytic Hormone: New Roles for an Old Player" Pharmaceuticals 14, no. 1: 45. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14010045
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.