
Extracellular Vesicles in Human Milk


1
Laboratory of Experimental Clinical Chemistry and Vesicle Observation Center, Amsterdam University Medical Center, University of Amsterdam, Meibergdreef 9, 1105 AZ Amsterdam, The Netherlands
2
Biomedical Engineering & Physics, Amsterdam University Medical Center, University of Amsterdam, Meibergdreef 9, 1105 AZ Amsterdam, The Netherlands
3
Clinical Division of Haematology and Haemostaseology, Department of Medicine I, Medical University of Vienna, Währinger Gürtel 18–20, 1090 Vienna, Austria
*
Author to whom correspondence should be addressed.
Pharmaceuticals 2021, 14(10), 1050; https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101050
Submission received: 6 July 2021
/
Revised: 5 October 2021
/
Accepted: 11 October 2021
/
Published: 15 October 2021
(This article belongs to the Special Issue Extracellular Vesicles: Biology and Emerging Therapeutic Opportunities)
Abstract
:Milk supports the growth and development of infants. An increasing number of mostly recent studies have demonstrated that milk contains a hitherto undescribed component called extracellular vesicles (EVs). This presents questions regarding why milk contains EVs and what their function is. Recently, we showed that EVs in human milk expose tissue factor, the protein that triggers coagulation or blood clotting, and that milk-derived EVs promote coagulation. Because bovine milk, which also contains EVs, completely lacks this coagulant activity, important differences are present in the biological functions of human milk-derived EVs between species. In this review, we will summarize the current knowledge regarding the presence and biochemical composition of milk EVs, their function(s) and potential clinical applications such as in probiotics, and the unique problems that milk EVs encounter in vivo, including survival of the gastrointestinal conditions encountered in the newborn. The main focus of this review will be human milk-derived EVs, but when available, we will also include information regarding non-human milk for comparison.
1. Introduction
Milk is produced by mammary glands and is the “natural food” of infants [1]. Milk contains such nutrients as carbohydrates, lipids, minerals, proteins, and vitamins [2,3,4]. The composition of milk differs between species; for example, human milk is relatively rich in lactose compared to milk from cows, goats and sheep, whereas milk from cows, goats and sheep contains more protein than human milk [5].
Breast feeding is associated with beneficial effects for the infant. Infants fed with human breast milk have less intestinal complications than infants fed with formula [6]. However, milk may have more beneficial effects. A famous example dates back to the 1930s, when Alphons Sole, an Austrian pediatrician, reported that external bleeding of boys suffering from haemophilia, a hereditary bleeding disorder, is effectively stopped by pressing human milk-soaked tamponades on the bleeding spots [7,8]. At that time, the bioactive component in milk responsible for this effect was not identified.
The question is, which beneficial effects of milk can be attributed to which components?
An increasing number of publications suggests that at least some of these beneficial effects can be attributed to the presence of extracellular vesicles (EVs). “Extracellular vesicles” is an umbrella term for all phospholipid bilayer-enclosed particles that are released by cells into their environment. The term “extracellular vesicles” includes exosomes and microvesicles or microparticles, also known as ectosomes. Body fluids concurrently contain different types of EVs, but since there are no unique biochemical and/or physical properties known yet to distinguish one type of EV from the other, we prefer to use the term “extracellular vesicles”. Consequently, even when, for example, the term “exosomes” is used in a manuscript, we will use the term “extracellular vesicles” to circumvent confusion and misinterpretation. All body fluids, including amniotic fluid, blood, saliva, and urine, contain EVs under physiological as well as pathological conditions [9,10,11,12]. The presence of EVs in human milk was first shown by electron microscopy in 1980s [13], and since then, the presence of EVs has been demonstrated in milk from camels, cows, goats, and pigs [14,15,16,17]. In contrast, EVs are absent in formula milk [18].
The International Society for Extracellular Vesicles (ISEV) introduced the term “extracellular vesicles” and has worked to actively improve the rigor, standardization and credibility of EV research by publishing the Minimal Information for Studies on Extracellular Vesicles and accompanying checklists [19]. Since 2019, the ISEV has maintained a Rigor and Standardization Subcommittee (www.isev.org), and recently, an editorial was published in which ongoing efforts were summarized with the overall goal of improving the quality of EV research [20].
In the following section, we will summarize the thus far reported physical properties and cellular origin of EVs in milk.
2. Physical Properties and Cellular Origin of Extracellular Vesicles in Milk
The reported size range of EVs in human and bovine milk are comparable, as shown in Table 1. EVs in human milk are reported to range in diameter from 50 to 265 nm [21,22,23], and in bovine milk from 44 to 200 nm [24,25]. The absolute concentration of EVs in milk is unknown, because until now, only the total concentration of “particles”, i.e., including EVs plus non-EV particles, has been measured in cell-free fractions of human [26,27] and bovine milk [27,28].
The cellular origin of milk EVs is mostly unexplored. Human milk contains EVs exposing epithelial cell adhesion molecule (EpCAM), and such EVs likely originate from the mammary epithelium [33]. Bovine milk also contains such EVs [34]. Recent publications showed that milk contains immune cells [35], and thus, one would expect to observe immune cell-derived EVs. However, in our hands, using a sensitive flow cytometry capable of detecting single EVs with a diameter >160 nm, we were hardly able to detect EVs from leukocytes, monocytes, B-cells, and T-cells. Platelet-derived EVs were also not detectable, indicating that both immune cell-derived and blood cell-derived EVs are scarce or absent [33]. Whether these EVs are indeed hardly present, or whether most are still undetected due to their small size and the limited number of exposed antigens one needs to label and identify such (single) EVs, awaits further studies. Milk also contains bacteria [36,37,38], and bacterial EVs may present a fraction of EVs present in milk [38].
The mammary epithelium differentiates during lactation [39,40]. The changes in histology are reflected by changes in the composition of milk, which shifts from colostrum to mature milk [41]. It has been reported that colostrum EVs are superior compared to EVs from mature milk with regards to protection against intestinal inflammation due to the higher concentrations of immune- and growth-related proteins associated with colostrum EVs [42,43].
3. Isolation of Extracellular Vesicles from Milk
To gain insight into the functions of EVs present in milk, it is essential to isolate EVs to study their biochemical composition and/or study their function(s) directly. The current isolation methods that have been used to isolate milk EVs were adopted from those developed to isolate EVs from plasma or conditioned cultured medium. The 125 articles that were retrieved on PubMed (using the terms “milk and (extracellular vesicles or exosomes or microvesicles)”) showed that milk EVs are isolated by commonly used EV isolation methods as (ultra)centrifugation, density gradient centrifugation, commercial precipitation kits, and size exclusion chromatography (SEC). These methods have been extensively described elsewhere, including their advantages and disadvantages, but with regard to milk, a few additional points should be discussed and taken into account due to the complex composition of milk, including the presence of milk fat globules and casein micelles, as shown in Figure 1.
3.1. Removal of Cells and Cell Fragments
In most studies, raw milk is centrifuged at approximately 2000× g to remove cells and cell fragments. In none of these studies, however, has the efficacy of this procedure been confirmed by measuring the concentration of remaining cells and/or cell fragments after centrifugation. Because residual cells and cell fragments will co-isolate with all current isolation procedures applied to EVs, this is a potential confounder in most, if not all, studies published thus far.
3.2. Removal of Milk Fat Globules
During the centrifugation step to remove cells and cell fragments, the milk fat globules will start to float and form a layer. Skimming after centrifugation removes most milk fat globules, but neither the efficacy of skimming nor the presence of residual milk fat globules has been quantified. In some studies, as a second step, filtration, is applied to remove the remaining milk fat globules, but neither the efficacy of filtration nor the possible loss of EVs has been quantified thus far. Because size ranges of milk fat globules and EVs overlap, fractions of “isolated EVs” from milk are likely to be contaminated by milk fat globules. Their presence can be demonstrated by measuring the presence of milk fat globule–epidermal growth factor–factor 8 (MFG-E8). From the results of the reported protein composition of isolated milk-derived EVs, shown in Section 4.1. Proteins -, it is clear that “isolated EVs” do contain MFG-E8 and, thus, are contaminated with milk fat globules.
3.3. Removal of Casein Micelles
Another major hurdle is the presence of casein. Casein is the most common milk protein, accounting for up to 80% of total cow milk protein [44] and 35% of total human milk protein [28,45]. Casein forms spherical colloidal aggregates which are called casein micelles, and which range in size from 20 nm to 600 nm [31], and thus also overlap in size with EVs. The diameter of casein micelles is temperature dependent, and, for example, the casein micelles in human milk have an average diameter of 100 nm at 37 °C but 570 nm at 4 °C [46].
Multiple methods have been reported to remove casein (micelles) from milk, including centrifugation, acid precipitation, calcium ion chelation, and chymosin treatment. To which extent these procedures also affect the presence and (functional) integrity of EVs present in milk, however, is largely unexplored.
Centrifugation at 20,000–75,000× g removes a fraction of casein micelles. At higher g forces, e.g., at 340,000× g, casein micelles will crosslink and form a solid gel, which can be removed. However, even at this high centrifugation force, still not all casein can be removed [27,47], and such procedures may result in EV loss due to cross-linking of casein micelles and EVs [48]. Acid precipitation may damage EVs [49]. Since the casein micelle structure is calcium ion-dependent, chelation of calcium ions induces dissociation of casein micelles [50]. At high concentrations of ethylenediaminetetraacetic acid (EDTA), i.e., up to 50 mmol/L, casein micelles dissociate [51]. However, at such high concentrations of EDTA, EVs may also dissociate (R. Nieuwland, personal communication). Finally, chymosin hydrolyzes and degrades casein, and casein is not detectable in EVs isolated by ultracentrifugation after chymosin treatment [48]. Taken together, the methods to remove casein are diverse, but to which extent such methods affect the presence and functional activities of EVs in milk needs further investigation.
4. The Biochemical Composition of Extracellular Vesicles: Proteins and RNA
As explained in the previous section, the isolation of EVs is essential to gain insight into their biochemical composition and thus also their function(s). At present, EVs are widely considered to be vehicles supporting intercellular communication by delivering biologically active cargoes, e.g., proteins and RNAs, from the parent cell to recipient cells [52]. An overview of the composition of isolated EVs from human and non-human origin are shown for proteins (Table 2 and Table 3) and RNA (Table 4 and Table 5).
4.1. Proteins
Several databases contain reported proteomes of milk EV proteins, including ExoCarta (http://exocarta.org/index.html) accessed on 20 March 2021, and Vesiclepedia (http://microvesicles.org/index.html) accessed on 20 March 2021. Most of the identified surface proteins are involved in peptide transport, membrane biosynthesis, and proteolysis, whereas the identified cytosolic proteins are involved in functional cell signaling pathways [53]. Several proteomes have identified tetraspannins, such as CD9, CD63 and CD81, which are claimed to be specific EV markers [27] (Table 2). Because transmembrane proteins require a phospholipid bilayer membrane, which is provided only by EVs in milk and not by casein micelles or milk fat globules, the presence of tetraspannins confirms that indeed EVs were isolated and studied, although it does not rule out the presence of non-EV particles. Of note, tissue factor, which is abundantly present in human milk and exclusively associated with EVs, is lacking in the published proteomes, illustrating that the current databases are certainly still incomplete [54]. Additionally, the fact that there are considerable differences in the reported protein composition of milk-derived EVs, even within the milk of one species, indicates that validation is essential. At present, there are also no studies yet that demonstrate or confirm the functions of the identified proteins in milk-derived isolated EVs.
Recently, EVs from human plasma and serum were shown to be covered by a set of proteins. Such proteins, called “corona proteins”, may be involved in various functions [55], and it seems reasonable to assume that also EVs that are present in other body fluids may be covered with such corona proteins, although the composition of such a corona may be body fluid- and perhaps disease (state)-dependent.

Table 2.
Main proteins identified in isolated extracellular vesicles from human milk.
| Isolation Procedure | Proteins | Detection | Study |
|---|---|---|---|
| Ultracentrifugation | CXCL5, SOD1, PRSS8, EPHA2, MIA, FR-alpha, MET, CD69, PDGF subunit B, CCL19 | PEA | [56] |
| Ultracentrifugation | MHC class II, CD81, MUC1, HSPs, MFG-E8 | FCM, WB, MS | [21] |
| Ultracentrifugation, size exclusive chromatography | TF, CD9, CD63 | WB | [33] |
| Density gradient centrifugation | Colostrum: LCP1, IGHG1, IGKV3-20, LAMP1, PRDX1 Mature milk: SLC1A5, RPL10, Tetraspanin, GMPPA | LC-MS/MS | [57] |
| Density gradient centrifugation | CD9, CD63, MHC-class II, FLOT-1 | LC-MS/MS, WB | [58] |
| Density gradient centrifugation | CD36, CD63, MUC1 | FCM | [59] |
Table 3.
Main proteins identified in isolated extracellular vesicles from livestock milk.
| Milk source | Isolation Procedure | Protein | Detection | Study |
|---|---|---|---|---|
| Bovine | Ultracentrifugation | CD63, HSP70, CD9, CD81 | WB | [60] |
| Bovine | Ultracentrifugation | CD9, CD81, CD63, CD82, CD47, MHC class I, syndecan, NT5E, CD59 | LC-MS/MS | [48] |
| Bovine | Ultracentrifugation | FASN, XDH, BTN1A1, HSPA8, PLIN2, MFG-E8, IDH1, GDI2 | LC-MS/MS | [61] |
| Bovine | Ultracentrifugation | CD63, CD81, TSG101, CD9 | WB | [62] |
| Bovine | Ultracentrifugation | LGB, PAEP, CSN1S1 | LC-MS/MS | [63] |
| Bovine (yak) | Ultracentrifugation | CD63, HSP70, TSG101 | WB | [64] |
| Bovine | Density gradient centrifugation | CD63, HSP70, MFG-E8, BTN1A1 | WB | [65] |
| Bovine | Density gradient centrifugation | Colostrum: B2M, Clusterin, PDGFC, CCN1 Mature milk: LTF, ANG1, LPO, QSOX1 | LC-MS/MS | [57] |
| Horse | Ultracentrifugation | CD81, CD63, LGB, MFG-E8 | MALDI-MS, MS/MS | [66] |
4.2. RNA
Isolated fractions of EVs are enriched in various types of RNAs, including micro RNAs (miRNAs), as shown in Table 4 and Table 5. Additionally, Vesiclepedia contains a section on miRNAs in milk EVs. At present, by using combinations of methods to isolate EVs, the previously reported and often assumed strong association between the presence of (mi)RNAs and EVs is becoming less clear and may have been overestimated in many studies [67]. Whether this also holds true for milk needs to be studied, but again, differences between study outcomes suggests that the RNA composition also needs validation.
Table 4.
Main microRNAs identified in isolated extracellular vesicles from human milk.
| Isolation Procedure | RNA | Detection | Study |
|---|---|---|---|
| Ultracentrifugation | hsa-miR-30d-5p, hsa-let-7b-5p, hsa-let-7a-5p, hsa-miR-125a-5p, hsa-miR-21–5p, hsa-miR-423–5p, hsa-let-7g-5p, hsa-let-7f-5p, hsa-miR-30a-5p, hsa-miR-146b-5p | RT-qPCR | [18] |
| Ultracentrifugation | Milk from mother who delivered pre-term infant: hsa-miR-22-3p, hsa-miR-148a-3p, hsa-miR-141-3p, hsa-miR-181a-5p, hsa-miR-320a, hsa-miR-378a-3p, hsa-miR-30d-5p, hsa-miR-30a-5p, hsa-miR-26a-5p, hsa-miR-191-5p; Milk from mother who delivered term infant: hsa-miR-22a-3p, hsa-miR-181a-5p, hsa-miR-148a-3p, hsa-miR-141a-3p, hsa-miR-30a-5p, hsa-miR-99b-5p, hsa-miR-191-5p, hsa-miR-378a-3p, hsa-miR-146b-5p, hsa-miR-30d-5p | UHTS | [68] |
| Ultracentrifugation | hsa-let-7c, hsa-miR-21, hsa-miR-34a, hsa-miR-146b, and hsa-miR-200b. | HTS | [69] |
| Density gradient centrifugation | hsa-miR-30d-5p, hsa-miR-148a-3p, hsa-miR-200a-3p, hsa-miR-200c-3p, hsa-let-7a-5p, hsa-miR-21-5p, hsa-let-7b-5p, hsa-let-7f-5p, hsa-miR-30a-5p, hsa-let-7g-5p | UHTS, RT-qPCR | [70] |
| ExoQuick-TC precipitation solution | hsa-miR-148a-3p, hsa-miR-22-3p, hsa-miR-30d-5p, hsa-let-7b-5p, hsa-miR-200a-3p | HTS | [71] |
| ExoQuick precipitation solution | hsa-miR-148a-3p, hsa-miR-30b-5p, hsa-let-7f-1-5p & -2-5p, hsa-miR-146b-5p, hsa-miR-29a-3p, hsa-let-7a-2-5p & -3-5p, hsa-miR-141-3p, hsa-miR-182-5p, hsa-miR-200a-3p, hsa-miR-378-3p | qPCR | [72] |
| ExoQuick-TC precipitation solution | hsa-miR-22-3p, hsa-miR-30d-5p, hsa-miR-148a-3p, hsa-miR-181a-5p, hsa-miR-141-3p, hsa-miR-30b-5p, hsa-miR-26a-5p, hsa-miR-92a-3p, hsa-miR-375, hsa-miR-30a-5p | UHTS | [73] |
| ExoQuick-TC precipitation solution | hsa-miR-148a-3p, hsa-miR-6073, hsa-miR-200c, hsa-miR-200b, hsa-miR-99a-5p, hsa-miR-30a-5p, hsa-miR-30d, hsa-miR-320-3p, hsa-let-7a-5p, hsa-miR-26a-5p | WGS | [74] |
| Total exosomes isolation reagent | miRNA from HIV-infected breast milk vs. uninfected; upregulated: hsa-miR-320e; hsa-miR-630; hsa-miR-148a-3p; hsa-miR-23a-3p; hsa-miR-378g; hsa-miR-30a-5p; hsa-miR-93-5p; hsa-miR-497-5p; hsa-miR-200b-3p; hsa-miR-200a-3p | NGS | [75] |
| ExoEasy maxi kit | hsa-miR-99b-3p, hsa-miR -96-5p, hsa-miR-550a-5p, hsa-miR-616-5p, hsa-miR-155-5p, hsa-miR-604 | qPCR | [76] |
Table 5.
Main microRNAs identified in isolated extracellular vesicles from livestock milk.
| Milk Source | Isolation Procedure | RNA | Detection | Study |
|---|---|---|---|---|
| Bovine | Ultracentrifugation | bta-miR-223, bta-miR125b | RT-qPCR | [61] |
| Bovine | Ultracentrifugation | Colostrum: bta-miR-24, bta-miR-30d, bta-miR-93, bta-miR-106a, bta-miR-181a, bta-miR-200a, bta-miR451 | RT-qPCR | [77] |
| Bovine | Ultracentrifugation | bta-miR-10b, bta-miR-143, bta-miR-10a, bta-miR-26a, bta-let-7a, bta-miR-21, bta-let-7f, bta-miR-222, bta-miR-99b, bta-let-7i | RT-qPCR | [78] |
| Bovine | Ultracentrifugation | bta-let-7a-5p, bta-let-7b, bta-let-7c, bta-let-7e and bta-let-7f, bta-miR-30a-5p, bta-miR-30d and bta-miR-30e-5p, bta-miR-148a, bta-miR-26a | HTS | [79] |
| Bovine | Ultracentrifugation, Density gradient centrifugation | bta-miR-223, bta-miR125b | RT-qPCR | [25] |
| Bovine | Total exosome isolation kit | bta-miR-26a, bta-miR-191, bta-miR-423-5p, bta-let-7f, bta-miR-30d, bta-let-7a-5p, bta-miR-27b, bta-let-7b, bta-miR-92a, bta-miR-125a | WGS | [80] |
| Bovine | ExoQuick reagent | - bta-miR-21, bta-miR-30a, bta-miR-92a, bta-miR-99a, bta-miR-223 | qPCR | [81] |
| Porcine | Ultracentrifugation | ssc-miR-193a-3p, ssc-miR-423-5p, ssc-miR-320, ssc-miR-181a, ssc-miR-30a3p, ssc-miR-378, ssc-miR-191, ssc-let-7a, ssc-let-7f, ssc-let-7c. | HTS | [82] |
| Porcine | Density gradient centrifugation | ssc-let-7a-5p, ssc-miR-30a-5p, ssc-miR-191-5p, ssc-miR-21-5p, ssc-miR-30d-5p, ssc-let-7f-5p, ssc-let-7c, ssc-miR-200c-3p, ssc-let-7g-5p, ssc-miR-320a-3p | HTS | [70] |
| Porcine | ExoQuick exosome precipitation solution | ssc-miR-148a-3p, ssc-miR-182-5p, ssc-miR-200c-3p, ssc-miR-25-3p, ssc-miR-30a-5p, ssc-miR-30d-5p, ssc-miR-574-3p | HTS | [83] |
| Caprinae (sheep) | Total exosome isolation kit | oar-miR-26a, oar-miR-191, oar-let-7f, oar-let-7b, oar-miR-10b, oar-miR-148a, oar-let-7b, oar-let-7a, oar-miR-21, oar-let-7c | HTS, PCR | [84] |
5. Functions of Extracellular Vesicles in Milk
Before introducing the reported functions of milk-derived EVs, which include coagulation, regulation of intestinal epithelium barrier function, anti-viral activity, and microbiome composition, it should be clear that such EVs can potentially have effects both on the mother and the infant. We will also give an overview of how the reported beneficial functions of milk-derived EVs have been explored, mostly in vitro and in animal models, for potential clinical utility, including osteoarthritis, cancer, and drug delivery.
5.1. Coagulation
Human milk triggers blood clotting [8], and recently, we demonstrated that the coagulant activity of human milk is due to the presence of tissue factor, which is present on EVs [33]. Tissue factor is a transmembrane protein. EVs are a suitable vehicle because they are surrounded by a phospholipid bilayer membrane, and because coagulation also requires a membrane surface [85]. To date, there is no evidence available that human milk contains coagulation factors other than TF.
Substantial nipple skin damage occurs in most breastfeeding women [86]. Fast activation of coagulation will quickly “seal the wound”, thereby reducing the risk of infection. Thus, one may speculate that tissue factor-exposing EVs in human milk may prevent infection and thus help to secure the mother’s health. In our study, surprisingly, bovine milk completely lacks this hemostasis-promoting property, and bovine milk-derived EVs are completely devoid of this tissue factor-coagulant activity, thus revealing a possibly functionally important difference between the milk of different mammalian species.
5.2. Intestinal Epithelium Barrier Function
The intestinal epithelial layer is essentially a barrier separating the body from bacteria that are present in the gastrointestinal tract. Regulation of tight junctions in this epithelial cell layer regulates the barrier permeability. When this barrier function becomes impaired, bacterial translocation can occur from the intestines into the blood, which leads to infection and sepsis [87].
The gastrointestinal tract of the pre-term infant is immature. After birth, nutrients such as oligosaccharides and immunoglobulins in milk facilitate the maturation of the gastrointestinal system [88]. The contribution of EVs to this barrier function has been studied in physiological and pathological models. With regards to the described effects of EVs to this barrier function, we can distinguish three main effects, i.e., the effects of EVs on proliferation, cell (tight) junctions, and mucin.
Firstly, in physiological models, proliferation of intestinal epithelial cells enhances the epithelial barrier function, and administration of milk-derived EVs promotes intestinal epithelial proliferation in rats [22] and pigs [89]. Furthermore, daily administration of porcine milk-derived EVs facilitated intestinal cell proliferation and intestinal tract development [85], with a possible involvement of the MAPK pathway [90].
In pathological (inflammatory) models, administration of human milk-derived EVs increased cell proliferation and decreased apoptosis in intestinal cells in vitro, and reduced intestinal epithelial injury and inflammation [91,92,93]. Additionally, rat and human milk-derived EVs promoted intestinal epithelial cell viability, enhanced proliferation of intestinal cells, and stimulated intestinal stem cell activity, thereby restoring the intestinal barrier function of newborns [22,23]. In a mouse colitis model, isolated EVs from bovine and human milk had an anti-inflammatory effect [94], whereas porcine milk EVs attenuated damage by promoting cell proliferation and tight junction formation via down-regulation of p53 [95]. Additionally, a diet with EV-depleted bovine milk induced intestinal inflammation in a mouse model [96]. Thus, the EVs present in milk seem to support intestinal integrity by inducing proliferation and regulation of permeability, but the underlying molecular mechanisms are unknown.
Secondly, the integrity of the intestinal epithelium depends on the junctions between cells [97]. There is evidence that milk EVs promotes the epithelial barrier function and reduces inflammation. Mice lacking a tight junction protein, kindlin-2, developed ulcerative colitis, which was reversed by treatment with bovine milk-derived EVs [98]. These findings suggest that milk EVs affect the tight junction function, and thus are involved in regulation of the permeability of the intestinal barrier.
Thirdly, the intestinal epithelium is covered by a layer of mucin, a protective additional barrier that is produced by goblet cells. Loss of mucin and reduced numbers of goblet cells are both associated with disease development of intestinal inflammation [99]. In an inflammation mucosal injury model, human milk-derived EVs increased the mucus production [93,99]. Treatment with bovine milk-derived EVs induced an increased expression of mucin-2; similarly, the addition of bovine milk-derived EVs to goblet cells stimulated mucin production and secretion, and prevented the development of intestinal inflammation [60]. In addition to this in vivo model of mucosal injury, intestinal mucosal injury and inflammation were attenuated by the administration of human milk EVs [93,99].
Although all evidence thus far confirms that milk-derived EVs promote epithelial barrier function, one must keep in mind that most, if not all, of these studies have used impure EVs and experiments were performed in mixed-model species, which hampers the interpretation of the reported results.
5.3. Anti-Viral Activity
Milk-derived EVs affect the immune responses of the mother by regulating the activity of immune cells [58,100]. For example, milk-derived EVs promoted the uptake of human immunodeficiency virus (HIV)-1 by macrophages and inhibited uptake by T cells [21,26]. Binding of EVs to antigen-presenting cells inhibited HIV infection of both dendritic cells and CD4+ T cells, which in turn may explain why an HIV-infected mother does not infect a breast-feeding baby [101]. Such an anti-viral activity was also reported for cytomegalovirus (CMV). CMV is regularly transmitted from the mother’s lactation epithelium to infants. Although transmitted CMV can be of great risk to the preterm infant, the reported cases are rare, suggesting that there are one or more protective mechanisms present in breast milk. EVs isolated from colostrum impaired the attachment of CMV to human foreskin fibroblast-1 cells, which was abolished by trypsin pretreatment of EVs, indicating a possible involvement of EV surface proteins [102].
5.4. Microbiome
The gastrointestinal epithelium forms a barrier, and thus prevents the entry of bacteria and bacterial toxins into the intestinal mucosa, and, eventually, into the systemic circulation [103]. This barrier function is dynamic and regulated by multiple stimuli [104]. Balanced microbial communities in the intestinal tract itself also support this barrier function. Bovine milk-derived EVs facilitated the gut microbiota composition, modulated bacterial metabolites, and regulated local intestinal immunity. For example, loss of Lachnospiraceae, a family of anaerobic, spore-forming bacteria, is associated with development of intestinal bowel disease [105,106], and the presence of Lachnospiraceae is supported by EV-enriched diets in mice [107]. Thus, potentially, milk EVs may be a valuable nutritional probiotic for human infants.
6. Therapeutic Application of Milk-Derived Extracellular Vesicles
6.1. Osteoporosis
Osteoporosis is a systemic skeletal disease characterized by weak and brittle bones, resulting in bone fragility and fracture susceptibility. EVs from bovine milk were reported to have both direct and indirect effects on bone mineral density. Regarding the direct effect, the addition of bovine milk-derived EVs regulated osteoblast and osteoclast differentiation and proliferation in vitro [108,109,110]. However, because milk-derived EVs will be present within the gastrointestinal tract, this direct effect seems unlikely to occur in vivo, because intact EVs would need to pass the epithelial barrier, travel to bones via the blood, and then regulate osteoblast and osteoclast differentiation and proliferation. Regarding the indirect effect, mice suffering from osteoporosis receiving bovine colostrum-derived EVs had higher bone mineral density compared to control mice. Because these changes were paralleled by changes in the microbiome, these authors speculated that bone mineral density may be affected by (indirect) EV-induced changes to the microbiome [111].
6.2. Arthritis
Consumption of bovine milk-derived EVs by mice suffering from arthritis protected cartilage and reduced inflammation of the bone marrow [112]. Interestingly, this protective effect of milk-derived EVs was paralleled by changes in the microbiome, again supporting the observation that milk-derived EVs may be part of the “gut-bone axis” [113,114].
6.3. Cancer
Oral administration of camel milk, or of camel milk-derived EVs orally or by local injection, reduced breast tumor progression in mice by inducing apoptosis of cancer tissue, increasing antioxidant enzyme activity, decreasing lipid peroxidation, and increasing oxidative stress in tumor tissue compared to the control group [115]. In contrast, human milk-derived EVs enhanced the epithelial-mesenchymal transition of breast cancer in vitro [116]. More work needs to be done to confirm how purified milk-derived EVs affect tumor growth and metastasis.
6.4. Drug Delivery
Studies are ongoing to investigate whether milk-derived EVs can be used as a cheap drug delivery vehicle instead of liposomes [117,118]. Most of these studies, however, suffer from multiple shortcomings. Firstly, milk EVs are difficult to isolate, and because there are no standard operating procedures, contamination with milk fat globules, casein micelles and soluble components cannot be excluded. Secondly, milk contains bacteria and viruses, and thus milk is not a sterile body fluid. Although there are procedures to disinfect milk, these procedures may also damage EVs. Especially for intravenous applications, sterilization of milk seems essential, whereas for oral applications it is recommended. Thirdly, milk EVs from non-human mammals other than cows and possibly relatives may expose tissue factor and thus trigger blood clotting. If so, intravenous administration of such EVs would directly trigger thrombus formation, which may even be lethal. Due to these limitations, administration of milk-derived EVs seems unsafe and requires additional studies.
The idea behind using (milk-derived) EVs for drug administration is to lower the dose of toxic anti-cancer drugs by improving their bioavailability. Thus, EVs are used as lipid carriers instead of liposomes, and by loading toxic drugs into milk-derived EVs, the total dose can be lowered [119]. For example, the therapeutic effectivity of paclitaxel, cisplatin, and doxorubicin was enhanced by packaging these drugs into EVs [120,121], and nude mice bearing human lung tumors were treated successfully with intravenously administered, paclitaxel-loaded bovine milk-derived EVs with low systemic and immunologic toxicities [122]. Whereas paclitaxel, cisplatin and doxorubicin are hydrophilic, milk-derived EVs have also been used as carriers of hydrophobic compounds such as curcumin, a substance isolated from the root of the turmeric plant reported to have anti-cancer potential [123]. When curcumin is incorporated into EVs from buffalo milk, curcumin had improved bioavailability compared to free curcumin [124]. Once loaded into EVs, curcumin was more easily taken up by cells [125], and tumor growth inhibition was stronger than non-packaged curcumin [120]. Finally, milk-derived EVs have also been used as carriers of siRNAs [126]. Taken together, milk-derived EVs may have potential for drug delivery, but one has to be careful because milk is not sterile and EVs may trigger blood clotting. EVs from mesenchymal progenitor cells or other mesenchymal cell types may be better for intravenous administration [127,128].
7. Survival of Extracellular Vesicles from Mothers’ Milk in the Infants’ Digestive Tract
Milk directly enters the gastrointestinal tract of the infant and thus faces digestion and degradation. Digestion affects the presence of bioactive components, including those associated with EVs. Thus, the question is whether EVs or the bioactive components of EVs may survive under physiological conditions of oral administration.
Digestion encompasses food entering the mouth, stomach, small intestines and colon. During digestion, food turns into macromolecules, which are absorbed as nutrients by the blood, and waste. In the oral digestion phase, milk is mixed with a minor volume of saliva. Then, in the gastric digestion phase (stomach), the EVs encounter an acidic environment, which is followed by enzymatic degradation in the small intestines, etc. Infants, especially pre-term infants, have an immature digestive system that is characterized by less harsh conditions than those encountered in the mature digestive system. For example, the pH of the gastric fluid of infants is higher than in adults [68,129,130], and concentrations of degrading enzymes, such as amylase, trypsin and lipase [131], and bile salts [130] are lower. In vitro, milk-derived EVs survive simulated gastric digestion and/or simulated intestinal digestion [73,130,132], which supports data indicating that endogenous milk EVs are beneficial, at least in infants [25,68,73,133].
8. Possible Uptake of Milk-Derived Extracellular Vesicles
As explained, drinking milk brings EVs into the infant’s digestive tract, and such EVs are likely to survive the relatively mild environmental conditions. To be taken up by the intestinal epithelial cells, the first barrier milk EVs have to pass is a layer of mucus covering the intestinal epithelium. Whether milk EVs are trapped in this layer, or how they can move across this layer, is unknown. In vitro studies have claimed that EVs bind to and are taken up by intestinal epithelial cells [73,112,134], vascular endothelial cells [135], macrophages [44], and splenocytes [112], and in a mouse model, milk-derived EVs were shown to be absorbed by the gut and to accumulate in several organs [136]. However, whether uptake of EVs by epithelial cells is affected by a layer of mucus in vitro is obscure. Furthermore, in the mouse model, EVs were labeled with a lipid membrane dye, and using such dyes is more often than not a source of artefacts [137].
9. Summary
Milk is an important body fluid as it is both consumable and the recommended food for newborns, since milk reduces intestinal complications. There is compelling evidence that milk contains EVs. At least some of the beneficial effects of milk may be attributed to the presence of EVs (Figure 2). Due to the lack of proper methods to purify EVs from milk, most results thus far regarding their biochemical composition and function awaits confirmation.
10. Outlook
The recommended practice of exclusively breastfeeding to at least 6 months after birth is not applied to all infants [138], and the benefits of human milk compared to formula encourages the current efforts of establishing donor milk banks for wider applications of human milk feeding to infants. To establish such biobanks, donor milk must be collected, transported, and stored. Because milk contains potential pathogens, collected donor milk needs to be disinfected. Although several disinfection methods are available, including pasteurization, at present it is unknown to which extent such methods affect the functions of milk and endogenous EVs. One may hypothesize that the coagulant activity of milk, which can be solely attributed to EVs present in milk, can be used as a read-out to evaluate the effects of collection and handling, including disinfection, and at the same time may provide insight into the intactness and functionality of at least a fraction of EVs present in such milk samples.
Author Contributions
Writing—original draft preparation, Y.H.; writing—review and editing, Y.H., J.T. and R.N.; visualization, Y.H.; supervision, J.T. and R.N. All authors have read and agreed to the published version of the manuscript.
Funding
Y.H. was funded by China Scholarship Council, grant number 201708320341.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| hsa | Homo sapiens |
| bta | Bos taurus |
| ssc | Sus scrofa |
| oar | Ovis aries |
| ANG1 | Angiogenin-1 |
| B2M | Beta-2-microglobulin |
| BTN1A1 | Butyrophilin, subfamily 1, member A1 |
| CCL19 | C-C motif chemokine ligand 19 |
| CCN1 | Cellular communication network factor 1 |
| CD | Cluster of differentiation |
| CSN1S1 | Casein alpha S1 |
| CXCL5 | C-X-C motif chemokine 5 |
| EVs | Extracellular vesicles |
| EPHA2 | EPH receptor A2 |
| FASN | Fatty acid synthase |
| FCM | Flow cytometry |
| FLOT-1 | Flotillin-1 |
| FR-alpha | Folate receptor 1 |
| GDI2 | Rab GDP dissociation inhibitor beta |
| GMPPA | Mannose-1-phosphate guanyltransferase alpha |
| HSP | Heat shock protein |
| HTS | High throughput sequencing |
| IDH1 | Isocitrate dehydrogenase |
| IGHG1 | Ig gamma-1 chain C region |
| IGKV3-20 | Ig kappa chain V-III region |
| LAMP1 | Lysosome-associated membrane glycoprotein 1 |
| LC-MS/MS | Liquid chromatography-tandem mass spectrometry analysis |
| LCP1 | Lymphocyte cytosolic protein 1 |
| LGB | Beta-lactoglobulin |
| LPO | Lactoperoxidase |
| LTF | Lactoferrin |
| MALDI | Matrix-assisted laser desorption/ionization |
| MET | Hepatocyte growth factor receptor |
| MFG-E8 | Milk fat globule-EGF factor 8 protein (lactadherin) |
| MHC | Major histocompatibility complex |
| MIA | Melanoma inhibitory activity |
| MS | Mass spectrometry |
| MS/MS | Tandem mass spectrometry |
| MUC1 | Mucin 1 |
| NGS | Next-generation sequencing |
| NT5E | 5′-nucleotidase |
| PAEP | Progestogen-associated endometrial protein |
| PEA | Proximity extension assay |
| PDGF | Plate-derived growth factor |
| PLIN2 | Perilipin 2 |
| PRDX1 | Peroxiredoxin-1 |
| PRSS8 | Serine protease 8 |
| qPCR | Quantitative polymerase chain reaction |
| QSOX1 | Quiescin sulfhydryl oxidase 1 |
| RPL10 | Ribosomal protein L10 |
| RT-qPCR | Quantitative reverse transcription polymerase chain reaction |
| SLC1A5 | Solute carrier family 1 member 5 |
| SOD1 | Superoxide dismutase 1 |
| TF | Tissue factor |
| TSG101 | Tumor susceptibility gene 101 |
| UHTS | Ultra-high throughput sequencing |
| WB | Western blot |
| WGS | Whole-genome sequencing |
| XDH | Xanthine dehydrogenase/oxidase |
References
- Corpeleijn, W.E.; Kouwenhoven, S.M.; Paap, M.C.; van Vliet, I.; Scheerder, I.; Muizer, Y.; Helder, O.K.; van Goudoever, J.B.; Vermeulen, M.J. Intake of own mother’s milk during the first days of life is associated with decreased morbidity and mortality in very low birth weight infants during the first 60 days of life. Neonatology 2012, 102, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Sosa-Castillo, E.; Rodriguez-Cruz, M.; Molto-Puigmarti, C. Genomics of lactation: Role of nutrigenomics and nutrigenetics in the fatty acid composition of human milk. Br. J. Nutr. 2017, 118, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.C. Milk nutritional composition and its role in human health. Nutrition 2014, 30, 619–627. [Google Scholar] [CrossRef]
- De la Torre Gomez, C.; Goreham, R.V.; Bech Serra, J.J.; Nann, T.; Kussmann, M. “Exosomics”-A Review of Biophysics, Biology and Biochemistry of Exosomes with a Focus on Human Breast Milk. Front. Genet. 2018, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.W.; Juárez, M.; Ramos, M.; Haenlein, G.F.W. Physico-chemical characteristics of goat and sheep milk. Small Rumin. Res. 2007, 68, 88–113. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, S.; Schanler, R.J.; Kim, J.H.; Patel, A.L.; Trawoger, R.; Kiechl-Kohlendorfer, U.; Chan, G.M.; Blanco, C.L.; Abrams, S.; Cotten, C.M.; et al. An exclusively human milk-based diet is associated with a lower rate of necrotizing enterocolitis than a diet of human milk and bovine milk-based products. J. Pediatr. 2010, 156, 562–567.e561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sole, A. Die Muttermilch als Blutstillungsmittel. Wien. Klin. Wochenschr. 1935, 38, 1354–1359. [Google Scholar] [CrossRef]
- Glanzmann, E. Einführung in die Kinderheilkunde; Springer: Vienna, Austria, 1934. [Google Scholar]
- Wolf, P. The nature and significance of platelet products in human plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Hell, L.; Wisgrill, L.; Ay, C.; Spittler, A.; Schwameis, M.; Jilma, B.; Pabinger, I.; Altevogt, P.; Thaler, J. Procoagulant extracellular vesicles in amniotic fluid. Transl. Res. 2017, 184, 12–20.e11. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Gool, E.; Berckmans, R.J.; Coumans, F.A.W.; Barendrecht, A.D.; Maas, C.; van der Wel, N.N.; Altevogt, P.; Sturk, A.; Nieuwland, R. Extracellular vesicles from human saliva promote hemostasis by delivering coagulant tissue factor to activated platelets. J. Thromb. Haemost. 2018, 16, 1153–1163. [Google Scholar] [CrossRef] [Green Version]
- Rikkert, L.G.; de Rond, L.; van Dam, A.; van Leeuwen, T.G.; Coumans, F.A.W.; de Reijke, T.M.; Terstappen, L.; Nieuwland, R. Detection of extracellular vesicles in plasma and urine of prostate cancer patients by flow cytometry and surface plasmon resonance imaging. PLoS ONE 2020, 15, e0233443. [Google Scholar] [CrossRef]
- Brooker, B.E. The epithelial cells and cell fragments in human milk. Cell Tissue Res. 1980, 210, 321–332. [Google Scholar] [CrossRef]
- Baddela, V.S.; Nayan, V.; Rani, P.; Onteru, S.K.; Singh, D. Physicochemical Biomolecular Insights into Buffalo Milk-Derived Nanovesicles. Appl. Biochem. Biotechnol. 2016, 178, 544–557. [Google Scholar] [CrossRef]
- Mecocci, S.; Gevi, F.; Pietrucci, D.; Cavinato, L.; Luly, F.R.; Pascucci, L.; Petrini, S.; Ascenzioni, F.; Zolla, L.; Chillemi, G.; et al. Anti-Inflammatory Potential of Cow, Donkey and Goat Milk Extracellular Vesicles as Revealed by Metabolomic Profile. Nutrients 2020, 12, 2908. [Google Scholar] [CrossRef]
- Ibrahim, H.M.; Mohammed-Geba, K.; Tawfic, A.A.; El-Magd, M.A. Camel milk exosomes modulate cyclophosphamide-induced oxidative stress and immuno-toxicity in rats. Food Funct. 2019, 10, 7523–7532. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.Y.; Hou, L.J.; Sun, J.J.; Zeng, B.; Xi, Q.Y.; Luo, J.Y.; Chen, T.; Zhang, Y.L. Porcine Milk Exosome MiRNAs Attenuate LPS-Induced Apoptosis through Inhibiting TLR4/NF-κB and p53 Pathways in Intestinal Epithelial Cells. J. Agric. Food Chem. 2019, 67, 9477–9491. [Google Scholar] [CrossRef] [PubMed]
- Leiferman, A.; Shu, J.; Upadhyaya, B.; Cui, J.; Zempleni, J. Storage of Extracellular Vesicles in Human Milk, and MicroRNA Profiles in Human Milk Exosomes and Infant Formulas. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Royo, F.; Théry, C.; Falcón-Pérez, J.M.; Nieuwland, R.; Witwer, K.W. Methods for Separation and Characterization of Extracellular Vesicles: Results of a Worldwide Survey Performed by the ISEV Rigor and Standardization Subcommittee. Cells 2020, 9, 1955. [Google Scholar] [CrossRef]
- Admyre, C.; Johansson, S.M.; Qazi, K.R.; Filen, J.J.; Lahesmaa, R.; Norman, M.; Neve, E.P.; Scheynius, A.; Gabrielsson, S. Exosomes with immune modulatory features are present in human breast milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef]
- Hock, A.; Miyake, H.; Li, B.; Lee, C.; Ermini, L.; Koike, Y.; Chen, Y.; Maattanen, P.; Zani, A.; Pierro, A. Breast milk-derived exosomes promote intestinal epithelial cell growth. J. Pediatr. Surg. 2017, 52, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, X.; Zhang, L.; Cai, J.; Zhou, Y.; Liu, H.; Hu, Y.; Chen, W.; Xu, S.; Liu, P.; et al. Identification and Peptidomic Profiling of Exosomes in Preterm Human Milk: Insights into Necrotizing Enterocolitis Prevention. Mol. Nutr. Food Res. 2019, 63, e1801247. [Google Scholar] [CrossRef]
- Yu, S.; Zhao, Z.; Xu, X.; Li, M.; Li, P. Characterization of three different types of extracellular vesicles and their impact on bacterial growth. Food Chem. 2019, 272, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Benmoussa, A.; Lee, C.H.; Laffont, B.; Savard, P.; Laugier, J.; Boilard, E.; Gilbert, C.; Fliss, I.; Provost, P. Commercial Dairy Cow Milk microRNAs Resist Digestion under Simulated Gastrointestinal Tract Conditions. J. Nutr. 2016, 146, 2206–2215. [Google Scholar] [CrossRef]
- Sims, B.; Farrow, A.L.; Williams, S.D.; Bansal, A.; Krendelchtchikov, A.; Gu, L.; Matthews, Q.L. Role of TIM-4 in exosome-dependent entry of HIV-1 into human immune cells. Int. J. Nanomed. 2017, 12, 4823–4833. [Google Scholar] [CrossRef] [Green Version]
- Blans, K.; Hansen, M.S.; Sorensen, L.V.; Hvam, M.L.; Howard, K.A.; Moller, A.; Wiking, L.; Larsen, L.B.; Rasmussen, J.T. Pellet-free isolation of human and bovine milk extracellular vesicles by size-exclusion chromatography. J. Extracell. Vesicles 2017, 6, 1294340. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Shimizu, K.; Rahman, M.; Ishikawa, H.; Takase, H.; Ugawa, S.; Okada, A.; Inoshima, Y. Efficient method for isolation of exosomes from raw bovine milk. Drug Dev. Ind. Pharm. 2019, 45, 359–364. [Google Scholar] [CrossRef]
- Kunz, C.; Lönnerdal, B. Casein micelles and casein subunits in human milk. In Protein and Non-Protein Nitrogen in Human Milk; CRC Press: Boca Raton, FL, USA, 1989; pp. 9–27. [Google Scholar]
- Veenstra, C.; Lenferink, A.; Petersen, W.; Steenbergen, W.; Bosschaart, N. Optical properties of human milk. Biomed. Opt. Express 2019, 10, 4059–4074. [Google Scholar] [CrossRef]
- Kamigaki, T.; Ito, Y.; Nishino, Y.; Miyazawa, A. Microstructural observation of casein micelles in milk by cryo-electron microscopy of vitreous sections (CEMOVIS). Microscopy 2018, 67, 164–170. [Google Scholar] [CrossRef]
- Michalski, M.C.; Briard, V.; Michel, F.; Tasson, F.; Poulain, P. Size distribution of fat globules in human colostrum, breast milk, and infant formula. J. Dairy Sci. 2005, 88, 1927–1940. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Hell, L.; Kendlbacher, R.A.; Hajji, N.; Hau, C.; van Dam, A.; Berckmans, R.J.; Wisgrill, L.; Ay, C.; Pabinger, I.; et al. Human milk triggers coagulation via tissue factor-exposing extracellular vesicles. Blood Adv. 2020, 4, 6274–6282. [Google Scholar] [CrossRef]
- Zhang, M.; Ma, Z.; Li, R.; Guo, S.; Qiu, Y.; Gao, X. Proteomic Analysis Reveals Proteins and Pathways Associated with Lactation in Bovine Mammary Epithelial Cell-Derived Exosomes. J. Proteome Res. 2020, 19, 3211–3219. [Google Scholar] [CrossRef]
- Li, S.; Zhang, L.; Zhou, Q.; Jiang, S.; Yang, Y.; Cao, Y. Characterization of Stem Cells and Immune Cells in Preterm and Term Mother’s Milk. J. Hum. Lact. 2019, 35, 528–534. [Google Scholar] [CrossRef]
- Chen, P.W.; Lin, Y.L.; Huang, M.S. Profiles of commensal and opportunistic bacteria in human milk from healthy donors in Taiwan. J. Food Drug Anal. 2018, 26, 1235–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jost, T.; Lacroix, C.; Braegger, C.; Chassard, C. Impact of human milk bacteria and oligosaccharides on neonatal gut microbiota establishment and gut health. Nutr. Rev. 2015, 73, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Yi, D.Y. Analysis of the human breast milk microbiome and bacterial extracellular vesicles in healthy mothers. Exp. Mol. Med. 2020, 52, 1288–1297. [Google Scholar] [CrossRef] [PubMed]
- Alex, A.; Bhandary, E.; McGuire, K.P. Anatomy and Physiology of the Breast during Pregnancy and Lactation. Adv. Exp. Med. Biol. 2020, 1252, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Honvo-Houéto, E.; Truchet, S. Indirect Immunofluorescence on Frozen Sections of Mouse Mammary Gland. J. Vis. Exp. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. North. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Zhang, R.; Qian, T.; Peng, X.; He, W.; Zheng, S.; Cao, Y.; Pierro, A.; Shen, C. A comparison of exosomes derived from different periods breast milk on protecting against intestinal organoid injury. Pediatr. Surg. Int. 2019, 35, 1363–1368. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.; Chisanga, D.; Liem, M.; Keerthikumar, S.; Anand, S.; Ang, C.S.; Adda, C.G.; Versteegen, E.; Jois, M.; Mathivanan, S. Bovine milk-derived exosomes from colostrum are enriched with proteins implicated in immune response and growth. Sci. Rep. 2017, 7, 5933. [Google Scholar] [CrossRef]
- Somiya, M.; Yoshioka, Y.; Ochiya, T. Biocompatibility of highly purified bovine milk-derived extracellular vesicles. J. Extracell. Vesicles 2018, 7, 1440132. [Google Scholar] [CrossRef] [Green Version]
- Hernell, O. Human milk vs. cow’s milk and the evolution of infant formulas. Nestle Nutr. Workshop Ser. Pediatr. Program. 2011, 67, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sood, S.M.; Herbert, P.J.; Slattery, C.W. Structural studies on casein micelles of human milk: Dissociation of beta-casein of different phosphorylation levels induced by cooling and ethylenediaminetetraacetate. J. Dairy Sci. 1997, 80, 628–633. [Google Scholar] [CrossRef]
- Stewart, P.S.; Puppione, D.L.; Patton, S. The presence of microvilli and other membrane fragments in the non-fat phase of bovine milk. Z Zellforsch. Mikrosk. Anat. 1972, 123, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Hao, H.; Zhang, X.; Zhang, Z.; Lv, Y.; Zhang, L.; Yi, H. Oral Administration of Bovine Milk-Derived Extracellular Vesicles Alters the Gut Microbiota and Enhances Intestinal Immunity in Mice. Mol. Nutr. Food Res. 2020, 64, e1901251. [Google Scholar] [CrossRef]
- Rahman, M.M.; Shimizu, K.; Yamauchi, M.; Takase, H.; Ugawa, S.; Okada, A.; Inoshima, Y. Acidification effects on isolation of extracellular vesicles from bovine milk. PLoS ONE 2019, 14, e0222613. [Google Scholar] [CrossRef] [Green Version]
- Griffin, M.C.; Lyster, R.L.; Price, J.C. The disaggregation of calcium-depleted casein micelles. Eur. J. Biochem. 1988, 174, 339–343. [Google Scholar] [CrossRef]
- Udabage, P.; McKinnon, I.R.; Augustin, M.A. Mineral and casein equilibria in milk: Effects of added salts and calcium-chelating agents. J. Dairy Res. 2000, 67, 361–370. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Gohar, M.; Gilois, N.; Graveline, R.; Garreau, C.; Sanchis, V.; Lereclus, D. A comparative study of Bacillus cereus, Bacillus thuringiensis and Bacillus anthracis extracellular proteomes. Proteomics 2005, 5, 3696–3711. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Sen, P. Structural modulation of factor VIIa by full-length tissue factor (TF(1-263)): Implication of novel interactions between EGF2 domain and TF. J. Biomol. Struct. Dyn. 2018, 36, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Palviainen, M.; Saraswat, M.; Varga, Z.; Kitka, D.; Neuvonen, M.; Puhka, M.; Joenväärä, S.; Renkonen, R.; Nieuwland, R.; Takatalo, M.; et al. Extracellular vesicles from human plasma and serum are carriers of extravesicular cargo-Implications for biomarker discovery. PLoS ONE 2020, 15, e0236439. [Google Scholar] [CrossRef] [PubMed]
- Larssen, P.; Wik, L.; Czarnewski, P.; Eldh, M.; Lof, L.; Ronquist, K.G.; Dubois, L.; Freyhult, E.; Gallant, C.J.; Oelrich, J.; et al. Tracing Cellular Origin of Human Exosomes Using Multiplex Proximity Extension Assays. Mol. Cell Proteom. 2017, 16, 502–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Song, D.; Cao, X.; Wu, R.; Liu, B.; Ye, W.; Wu, J.; Yue, X. Comparative proteomic analysis of milk-derived exosomes in human and bovine colostrum and mature milk samples by iTRAQ-coupled LC-MS/MS. Food Res. Int. 2017, 92, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Van Herwijnen, M.J.; Zonneveld, M.I.; Goerdayal, S.; Nolte-‘t Hoen, E.N.; Garssen, J.; Stahl, B.; Maarten Altelaar, A.F.; Redegeld, F.A.; Wauben, M.H. Comprehensive Proteomic Analysis of Human Milk-derived Extracellular Vesicles Unveils a Novel Functional Proteome Distinct from Other Milk Components. Mol. Cell Proteom. 2016, 15, 3412–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torregrosa Paredes, P.; Gutzeit, C.; Johansson, S.; Admyre, C.; Stenius, F.; Alm, J.; Scheynius, A.; Gabrielsson, S. Differences in exosome populations in human breast milk in relation to allergic sensitization and lifestyle. Allergy 2014, 69, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Hock, A.; Wu, R.Y.; Minich, A.; Botts, S.R.; Lee, C.; Antounians, L.; Miyake, H.; Koike, Y.; Chen, Y.; et al. Bovine milk-derived exosomes enhance goblet cell activity and prevent the development of experimental necrotizing enterocolitis. PLoS ONE 2019, 14, e0211431. [Google Scholar] [CrossRef]
- Benmoussa, A.; Ly, S.; Shan, S.T.; Laugier, J.; Boilard, E.; Gilbert, C.; Provost, P. A subset of extracellular vesicles carries the bulk of microRNAs in commercial dairy cow’s milk. J. Extracell. Vesicles 2017, 6, 1401897. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Niu, M.; Hao, Z.; Liu, M.; Tong, C.; Zhao, X. Selective packaged circular RNAs in milk extracellular vesicles during Staphylococcus aureus infection may have potential against bacterial infection. RNA Biol. 2021, 18, 818–831. [Google Scholar] [CrossRef]
- Benmoussa, A.; Gotti, C.; Bourassa, S.; Gilbert, C.; Provost, P. Identification of protein markers for extracellular vesicle (EV) subsets in cow’s milk. J. Proteom. 2019, 192, 78–88. [Google Scholar] [CrossRef]
- Gao, H.N.; Guo, H.Y.; Zhang, H.; Xie, X.L.; Wen, P.C.; Ren, F.Z. Yak-milk-derived exosomes promote proliferation of intestinal epithelial cells in an hypoxic environment. J. Dairy Sci. 2019, 102, 985–996. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, B.; Buschmann, D.; Paul, V.; Pfaffl, M.W. Postprandial transfer of colostral extracellular vesicles and their protein and miRNA cargo in neonatal calves. PLoS ONE 2020, 15, e0229606. [Google Scholar] [CrossRef]
- Sedykh, S.E.; Purvinish, L.V.; Monogarov, A.S.; Burkova, E.E.; Grigor’eva, A.E.; Bulgakov, D.V.; Dmitrenok, P.S.; Vlassov, V.V.; Ryabchikova, E.I.; Nevinsky, G.A. Purified horse milk exosomes contain an unpredictable small number of major proteins. Biochim. Open 2017, 4, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Albanese, M.; Chen, Y.-F.A.; Hüls, C.; Gärtner, K.; Tagawa, T.; Mejias-Perez, E.; Keppler, O.T.; Göbel, C.; Zeidler, R.; Shein, M.; et al. Micro RNAs are minor constituents of extracellular vesicles and are hardly delivered to target cells. bioRxiv 2020, 2020.05.20.106393. [Google Scholar] [CrossRef]
- Kahn, S.; Liao, Y.; Du, X.; Xu, W.; Li, J.; Lonnerdal, B. Exosomal MicroRNAs in Milk from Mothers Delivering Preterm Infants Survive in Vitro Digestion and Are Taken Up by Human Intestinal Cells. Mol. Nutr. Food Res. 2018, 62, e1701050. [Google Scholar] [CrossRef] [PubMed]
- Mirza, A.H.; Kaur, S.; Nielsen, L.B.; Størling, J.; Yarani, R.; Roursgaard, M.; Mathiesen, E.R.; Damm, P.; Svare, J.; Mortensen, H.B.; et al. Breast Milk-Derived Extracellular Vesicles Enriched in Exosomes from Mothers with Type 1 Diabetes Contain Aberrant Levels of microRNAs. Front. Immunol. 2019, 10, 2543. [Google Scholar] [CrossRef]
- Van Herwijnen, M.J.C.; Driedonks, T.A.P.; Snoek, B.L.; Kroon, A.M.T.; Kleinjan, M.; Jorritsma, R.; Pieterse, C.M.J.; Hoen, E.; Wauben, M.H.M. Abundantly Present miRNAs in Milk-Derived Extracellular Vesicles Are Conserved Between Mammals. Front. Nutr. 2018, 5, 81. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.R.; Brede, G.; Johansen, J.; Johnsen, R.; Storrø, O.; Sætrom, P.; Øien, T. Human Breast Milk miRNA, Maternal Probiotic Supplementation and Atopic Dermatitis in Offspring. PLoS ONE 2015, 10, e0143496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Li, M.; Wang, X.; Li, Q.; Wang, T.; Zhu, Q.; Zhou, X.; Wang, X.; Gao, X.; Li, X. Immune-related microRNAs are abundant in breast milk exosomes. Int. J. Biol. Sci. 2012, 8, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Du, X.; Li, J.; Lonnerdal, B. Human milk exosomes and their microRNAs survive digestion in vitro and are taken up by human intestinal cells. Mol. Nutr. Food Res. 2017, 61, 1700082. [Google Scholar] [CrossRef]
- Golan-Gerstl, R.; Elbaum Shiff, Y.; Moshayoff, V.; Schecter, D.; Leshkowitz, D.; Reif, S. Characterization and biological function of milk-derived miRNAs. Mol. Nutr. Food Res. 2017, 61, 1700009. [Google Scholar] [CrossRef]
- Zahoor, M.A.; Yao, X.D.; Henrick, B.M.; Verschoor, C.P.; Abimiku, A.; Osawe, S.; Rosenthal, K.L. Expression profiling of human milk derived exosomal microRNAs and their targets in HIV-1 infected mothers. Sci. Rep. 2020, 10, 12931. [Google Scholar] [CrossRef]
- Bozack, A.K.; Colicino, E.; Rodosthenous, R.; Bloomquist, T.R.; Baccarelli, A.A.; Wright, R.O.; Wright, R.J.; Lee, A.G. Associations between maternal lifetime stressors and negative events in pregnancy and breast milk-derived extracellular vesicle microRNAs in the programming of intergenerational stress mechanisms (PRISM) pregnancy cohort. Epigenetics 2021, 16, 389–404. [Google Scholar] [CrossRef]
- Sun, Q.; Chen, X.; Yu, J.; Zen, K.; Zhang, C.Y.; Li, L. Immune modulatory function of abundant immune-related microRNAs in microvesicles from bovine colostrum. Protein Cell 2013, 4, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Benmoussa, A.; Laugier, J.; Beauparlant, C.J.; Lambert, M.; Droit, A.; Provost, P. Complexity of the microRNA transcriptome of cow milk and milk-derived extracellular vesicles isolated via differential ultracentrifugation. J. Dairy Sci. 2020, 103, 16–29. [Google Scholar] [CrossRef]
- Chen, Z.; Xie, Y.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Milk exosome-derived miRNAs from water buffalo are implicated in immune response and metabolism process. BMC Vet. Res. 2020, 16, 123. [Google Scholar] [CrossRef] [PubMed]
- Quan, S.Y.; Nan, X.M.; Wang, K.; Zhao, Y.G.; Jiang, L.S.; Yao, J.H.; Xiong, B.H. Replacement of forage fiber with non-forage fiber sources in dairy cow diets changes milk extracellular vesicle-miRNA expression. Food Funct. 2020, 11, 2154–2162. [Google Scholar] [CrossRef] [PubMed]
- Pieters, B.C.; Arntz, O.J.; Bennink, M.B.; Broeren, M.G.; van Caam, A.P.; Koenders, M.I.; van Lent, P.L.; van den Berg, W.B.; de Vries, M.; van der Kraan, P.M.; et al. Commercial cow milk contains physically stable extracellular vesicles expressing immunoregulatory TGF-beta. PLoS ONE 2015, 10, e0121123. [Google Scholar] [CrossRef]
- Chen, T.; Xi, Q.Y.; Ye, R.S.; Cheng, X.; Qi, Q.E.; Wang, S.B.; Shu, G.; Wang, L.N.; Zhu, X.T.; Jiang, Q.Y.; et al. Exploration of microRNAs in porcine milk exosomes. BMC Genom. 2014, 15, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Li, M.; Wang, T.; Liang, Y.; Zhong, Z.; Wang, X.; Zhou, Q.; Chen, L.; Lang, Q.; He, Z.; et al. Lactation-related microRNA expression profiles of porcine breast milk exosomes. PLoS ONE 2012, 7, e43691. [Google Scholar] [CrossRef]
- Quan, S.; Nan, X.; Wang, K.; Jiang, L.; Yao, J.; Xiong, B. Characterization of Sheep Milk Extracellular Vesicle-miRNA by Sequencing and Comparison with Cow Milk. Animals 2020, 10, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripisciano, C.; Weiss, R.; Eichhorn, T.; Spittler, A.; Heuser, T.; Fischer, M.B.; Weber, V. Different Potential of Extracellular Vesicles to Support Thrombin Generation: Contributions of Phosphatidylserine, Tissue Factor, and Cellular Origin. Sci. Rep. 2017, 7, 6522. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Asaka, Y.; Ogawara, T.; Yorozu, Y. Nipple Skin Trauma in Breastfeeding Women During Postpartum Week One. Breastfeed. Med. 2018, 13, 479–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foitzik, T.; Kruschewski, M.; Kroesen, A.J.; Hotz, H.G.; Eibl, G.; Buhr, H.J. Does glutamine reduce bacterial translocation? A study in two animal models with impaired gut barrier. Int. J. Colorectal Dis. 1999, 14, 143–149. [Google Scholar] [CrossRef]
- Nolan, L.S.; Parks, O.B.; Good, M. A Review of the Immunomodulating Components of Maternal Breast Milk and Protection Against Necrotizing Enterocolitis. Nutrients 2019, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Xie, M.Y.; Sun, J.J.; Ye, R.S.; Cheng, X.; Sun, R.P.; Wei, L.M.; Li, M.; Lin, D.L.; Jiang, Q.Y.; et al. Porcine milk-derived exosomes promote proliferation of intestinal epithelial cells. Sci. Rep. 2016, 6, 33862. [Google Scholar] [CrossRef]
- Yu, S.; Zhao, Z.; Sun, L.; Li, P. Fermentation Results in Quantitative Changes in Milk-Derived Exosomes and Different Effects on Cell Growth and Survival. J. Agric. Food Chem. 2017, 65, 1220–1228. [Google Scholar] [CrossRef]
- Pisano, C.; Galley, J.; Elbahrawy, M.; Wang, Y.; Farrell, A.; Brigstock, D.; Besner, G.E. Human Breast Milk-Derived Extracellular Vesicles in the Protection Against Experimental Necrotizing Enterocolitis. J. Pediatr. Surg. 2020, 55, 54–58. [Google Scholar] [CrossRef] [Green Version]
- Reif, S.; Elbaum Shiff, Y.; Golan-Gerstl, R. Milk-derived exosomes (MDEs) have a different biological effect on normal fetal colon epithelial cells compared to colon tumor cells in a miRNA-dependent manner. J. Transl. Med. 2019, 17, 325. [Google Scholar] [CrossRef]
- Miyake, H.; Lee, C.; Chusilp, S.; Bhalla, M.; Li, B.; Pitino, M.; Seo, S.; O’Connor, D.L.; Pierro, A. Human breast milk exosomes attenuate intestinal damage. Pediatr. Surg. Int. 2020, 36, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Reif, S.; Elbaum-Shiff, Y.; Koroukhov, N.; Shilo, I.; Musseri, M.; Golan-Gerstl, R. Cow and Human Milk-Derived Exosomes Ameliorate Colitis in DSS Murine Model. Nutrients 2020, 12, 2589. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.Y.; Chen, T.; Xi, Q.Y.; Hou, L.J.; Luo, J.Y.; Zeng, B.; Li, M.; Sun, J.J.; Zhang, Y.L. Porcine milk exosome miRNAs protect intestinal epithelial cells against deoxynivalenol-induced damage. Biochem. Pharmacol. 2020, 175, 113898. [Google Scholar] [CrossRef]
- Wu, D.; Kittana, H.; Shu, J.; Kachman, S.D.; Cui, J.; Ramer-Tait, A.E.; Zempleni, J. Dietary Depletion of Milk Exosomes and Their MicroRNA Cargos Elicits a Depletion of miR-200a-3p and Elevated Intestinal Inflammation and Chemokine (C-X-C Motif) Ligand 9 Expression in Mdr1a(−/−) Mice. Curr. Dev. Nutr. 2019, 3, nzz122. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight Junction in the Intestinal Epithelium: Its Association with Diseases and Regulation by Phytochemicals. J. Immunol. Res. 2018, 2018, 2645465. [Google Scholar] [CrossRef] [Green Version]
- Stremmel, W.; Weiskirchen, R.; Melnik, B.C. Milk Exosomes Prevent Intestinal Inflammation in a Genetic Mouse Model of Ulcerative Colitis: A Pilot Experiment. Inflamm. Intest. Dis. 2020, 5, 117–123. [Google Scholar] [CrossRef]
- Wu, R.Y.; Li, B.; Koike, Y.; Maattanen, P.; Miyake, H.; Cadete, M.; Johnson-Henry, K.C.; Botts, S.R.; Lee, C.; Abrahamsson, T.R.; et al. Human Milk Oligosaccharides Increase Mucin Expression in Experimental Necrotizing Enterocolitis. Mol. Nutr. Food Res. 2019, 63, e1800658. [Google Scholar] [CrossRef]
- Komine-Aizawa, S.; Ito, S.; Aizawa, S.; Namiki, T.; Hayakawa, S. Cow milk exosomes activate NK cells and γδT cells in human PBMCs in vitro. Immunol. Med. 2020, 43, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Naslund, T.I.; Paquin-Proulx, D.; Paredes, P.T.; Vallhov, H.; Sandberg, J.K.; Gabrielsson, S. Exosomes from breast milk inhibit HIV-1 infection of dendritic cells and subsequent viral transfer to CD4+ T cells. Aids 2014, 28, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Donalisio, M.; Cirrincione, S.; Rittà, M.; Lamberti, C.; Civra, A.; Francese, R.; Tonetto, P.; Sottemano, S.; Manfredi, M.; Lorenzato, A.; et al. Extracellular Vesicles in Human Preterm Colostrum Inhibit Infection by Human Cytomegalovirus In Vitro. Microorganisms 2020, 8, 1087. [Google Scholar] [CrossRef]
- Shukla, P.K.; Gangwar, R.; Manda, B.; Meena, A.S.; Yadav, N.; Szabo, E.; Balogh, A.; Lee, S.C.; Tigyi, G.; Rao, R. Rapid disruption of intestinal epithelial tight junction and barrier dysfunction by ionizing radiation in mouse colon in vivo: Protection by N-acetyl-l-cysteine. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G705–G715. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Yang, S.; Qiu, Y.; Chen, G.; Wang, W.; Xu, C.; Cai, W.; Sun, L.; Xiao, W.; Yang, H. Par-3 modulates intestinal epithelial barrier function through regulating intracellular trafficking of occludin and myosin light chain phosphorylation. J. Gastroenterol. 2015, 50, 1103–1113. [Google Scholar] [CrossRef]
- Baumgart, M.; Dogan, B.; Rishniw, M.; Weitzman, G.; Bosworth, B.; Yantiss, R.; Orsi, R.H.; Wiedmann, M.; McDonough, P.; Kim, S.G.; et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn’s disease involving the ileum. ISME J. 2007, 1, 403–418. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.N.; St Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Paz, H.A.; Sadri, M.; Cui, J.; Kachman, S.D.; Fernando, S.C.; Zempleni, J. Dietary bovine milk exosomes elicit changes in bacterial communities in C57BL/6 mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G618–g624. [Google Scholar] [CrossRef]
- Oliveira, M.C.; Di Ceglie, I.; Arntz, O.J.; van den Berg, W.B.; van den Hoogen, F.H.; Ferreira, A.V.; van Lent, P.L.; van de Loo, F.A. Milk-Derived Nanoparticle Fraction Promotes the Formation of Small Osteoclasts but Reduces Bone Resorption. J. Cell Physiol. 2017, 232, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.C.; Arntz, O.J.; Blaney Davidson, E.N.; van Lent, P.L.; Koenders, M.I.; van der Kraan, P.M.; van den Berg, W.B.; Ferreira, A.V.; van de Loo, F.A. Milk extracellular vesicles accelerate osteoblastogenesis but impair bone matrix formation. J. Nutr. Biochem. 2016, 30, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.C.; Pieters, B.C.H.; Guimarães, P.B.; Duffles, L.F.; Heredia, J.E.; Silveira, A.L.M.; Oliveira, A.C.C.; Teixeira, M.M.; Ferreira, A.V.M.; Silva, T.A.; et al. Bovine Milk Extracellular Vesicles Are Osteoprotective by Increasing Osteocyte Numbers and Targeting RANKL/OPG System in Experimental Models of Bone Loss. Front. Bioeng. Biotechnol. 2020, 8, 891. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.; Maburutse, B.E.; Kang, M.; Park, M.R.; Park, D.J.; Kim, Y.; Oh, S. Short communication: Dietary bovine milk-derived exosomes improve bone health in an osteoporosis-induced mouse model. J. Dairy Sci. 2020. [Google Scholar] [CrossRef]
- Arntz, O.J.; Pieters, B.C.; Oliveira, M.C.; Broeren, M.G.; Bennink, M.B.; de Vries, M.; van Lent, P.L.; Koenders, M.I.; van den Berg, W.B.; van der Kraan, P.M.; et al. Oral administration of bovine milk derived extracellular vesicles attenuates arthritis in two mouse models. Mol. Nutr. Food Res. 2015, 59, 1701–1712. [Google Scholar] [CrossRef]
- Guerreiro, C.S.; Calado, Â.; Sousa, J.; Fonseca, J.E. Diet, Microbiota, and Gut Permeability-The Unknown Triad in Rheumatoid Arthritis. Front. Med. 2018, 5, 349. [Google Scholar] [CrossRef] [Green Version]
- Aarts, J.; Boleij, A.; Pieters, B.C.H.; Feitsma, A.L.; van Neerven, R.J.J.; Ten Klooster, J.P.; M’Rabet, L.; Arntz, O.J.; Koenders, M.I.; van de Loo, F.A.J. Flood Control: How Milk-Derived Extracellular Vesicles Can Help to Improve the Intestinal Barrier Function and Break the Gut-Joint Axis in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 703277. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.; El-Magd, M.A.; AlSadrah, S.A. Therapeutic Effect of Camel Milk and Its Exosomes on MCF7 Cells In Vitro and In Vivo. Integr. Cancer Ther. 2018, 17, 1235–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, W.; Tsukasaki, Y.; Dasgupta, S.; Mukhopadhyay, N.; Ikebe, M.; Sauter, E.R. Exosomes in Human Breast Milk Promote EMT. Clin. Cancer Res. 2016, 22, 4517–4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, A.; Patel, T. Milk-derived Extracellular Vesicles for Therapeutic Delivery of Small Interfering RNAs. Methods Mol. Biol. 2018, 1740, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, A.; Moirangthem, A.; Angom, R.S.; Ishiguro, K.; Driscoll, J.; Yan, I.K.; Mukhopadhyay, D.; Patel, T. Safety of bovine milk derived extracellular vesicles used for delivery of RNA therapeutics in zebrafish and mice. J. Appl. Toxicol. 2019, 40, 706–718. [Google Scholar] [CrossRef]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Agrawal, A.K.; Gupta, R. Exosomes for the Enhanced Tissue Bioavailability and Efficacy of Curcumin. AAPS J. 2017, 19, 1691–1702. [Google Scholar] [CrossRef]
- Li, D.; Yao, S.; Zhou, Z.; Shi, J.; Huang, Z.; Wu, Z. Hyaluronan decoration of milk exosomes directs tumor-specific delivery of doxorubicin. Carbohydr. Res. 2020, 493, 108032. [Google Scholar] [CrossRef]
- Agrawal, A.K.; Aqil, F.; Jeyabalan, J.; Spencer, W.A.; Beck, J.; Gachuki, B.W.; Alhakeem, S.S.; Oben, K.; Munagala, R.; Bondada, S.; et al. Milk-derived exosomes for oral delivery of paclitaxel. Nanomedicine 2017, 13, 1627–1636. [Google Scholar] [CrossRef]
- Kunihiro, A.G.; Brickey, J.A.; Frye, J.B.; Luis, P.B.; Schneider, C.; Funk, J.L. Curcumin, but not curcumin-glucuronide, inhibits Smad signaling in TGFβ-dependent bone metastatic breast cancer cells and is enriched in bone compared to other tissues. J. Nutr. Biochem. 2019, 63, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Vashisht, M.; Rani, P.; Onteru, S.K.; Singh, D. Curcumin Encapsulated in Milk Exosomes Resists Human Digestion and Possesses Enhanced Intestinal Permeability in Vitro. Appl. Biochem. Biotechnol. 2017, 183, 993–1007. [Google Scholar] [CrossRef] [PubMed]
- Carobolante, G.; Mantaj, J.; Ferrari, E.; Vllasaliu, D. Cow Milk and Intestinal Epithelial Cell-derived Extracellular Vesicles as Systems for Enhancing Oral Drug Delivery. Pharmaceutics 2020, 12, 226. [Google Scholar] [CrossRef] [Green Version]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Agrawal, A.K.; Kyakulaga, A.H.; Wilcher, S.A.; Gupta, R.C. Milk exosomes—Natural nanoparticles for siRNA delivery. Cancer Lett. 2019, 449, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Laso-García, F.; Ramos-Cejudo, J.; Carrillo-Salinas, F.J.; Otero-Ortega, L.; Feliú, A.; Gómez-de Frutos, M.; Mecha, M.; Díez-Tejedor, E.; Guaza, C.; Gutiérrez-Fernández, M. Therapeutic potential of extracellular vesicles derived from human mesenchymal stem cells in a model of progressive multiple sclerosis. PLoS ONE 2018, 13, e0202590. [Google Scholar] [CrossRef] [Green Version]
- Nassar, W.; El-Ansary, M.; Sabry, D.; Mostafa, M.A.; Fayad, T.; Kotb, E.; Temraz, M.; Saad, A.N.; Essa, W.; Adel, H. Umbilical cord mesenchymal stem cells derived extracellular vesicles can safely ameliorate the progression of chronic kidney diseases. Biomater. Res. 2016, 20, 21. [Google Scholar] [CrossRef] [Green Version]
- Mason, S. Some aspects of gastric function in the newborn. Arch. Dis. Child. 1962, 37, 387–391. [Google Scholar] [CrossRef] [Green Version]
- Kamstrup, D.; Berthelsen, R.; Sassene, P.J.; Selen, A.; Mullertz, A. In Vitro Model Simulating Gastro-Intestinal Digestion in the Pediatric Population (Neonates and Young Infants). AAPS PharmSciTech 2017, 18, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Zoppi, G.; Andreotti, G.; Pajno-Ferrara, F.; Njai, D.M.; Gaburro, D. Exocrine pancreas function in premature and full term neonates. Pediatr. Res. 1972, 6, 880–886. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Ye, A.; Liu, W.; Liu, C.; Singh, H. Stability during in vitro digestion of lactoferrin-loaded liposomes prepared from milk fat globule membrane-derived phospholipids. J. Dairy Sci. 2013, 96, 2061–2070. [Google Scholar] [CrossRef] [Green Version]
- Hata, T.; Murakami, K.; Nakatani, H.; Yamamoto, Y.; Matsuda, T.; Aoki, N. Isolation of bovine milk-derived microvesicles carrying mRNAs and microRNAs. Biochem. Biophys. Res. Commun. 2010, 396, 528–533. [Google Scholar] [CrossRef]
- Wolf, T.; Baier, S.R.; Zempleni, J. The Intestinal Transport of Bovine Milk Exosomes Is Mediated by Endocytosis in Human Colon Carcinoma Caco-2 Cells and Rat Small Intestinal IEC-6 Cells. J. Nutr. 2015, 145, 2201–2206. [Google Scholar] [CrossRef] [Green Version]
- Kusuma, R.J.; Manca, S.; Friemel, T.; Sukreet, S.; Nguyen, C.; Zempleni, J. Human vascular endothelial cells transport foreign exosomes from cow’s milk by endocytosis. Am. J. Physiol. Cell Physiol. 2016, 310, C800–C807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betker, J.L.; Angle, B.M.; Graner, M.W.; Anchordoquy, T.J. The Potential of Exosomes from Cow Milk for Oral Delivery. J. Pharm. Sci. 2019, 108, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- De Rond, L.; van der Pol, E.; Hau, C.M.; Varga, Z.; Sturk, A.; van Leeuwen, T.G.; Nieuwland, R.; Coumans, F.A.W. Comparison of Generic Fluorescent Markers for Detection of Extracellular Vesicles by Flow Cytometry. Clin. Chem. 2018, 64, 680–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagci Bosi, A.T.; Eriksen, K.G.; Sobko, T.; Wijnhoven, T.M.; Breda, J. Breastfeeding practices and policies in WHO European Region Member States. Public Health Nutr. 2016, 19, 753–764. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Processing of milk before isolation of extracellular vesicles. Raw milk contains cells, cell fragments, milk fat globules, casein micelles, and extracellular vesicles (EVs). As a first step, cells, cell fragments and milk fat globules are mostly removed by centrifugation. Milk fat globules will float to the top due to their low density, and cells and cell fragments will form a pellet. The middle layer contains casein micelles and EVs. This middle layer is the starting material for the isolation of EVs. Casein micelles can be removed by acid-induced or enzyme-induced precipitation, centrifugation, or by EDTA-induced dissociation.
Figure 1.
Processing of milk before isolation of extracellular vesicles. Raw milk contains cells, cell fragments, milk fat globules, casein micelles, and extracellular vesicles (EVs). As a first step, cells, cell fragments and milk fat globules are mostly removed by centrifugation. Milk fat globules will float to the top due to their low density, and cells and cell fragments will form a pellet. The middle layer contains casein micelles and EVs. This middle layer is the starting material for the isolation of EVs. Casein micelles can be removed by acid-induced or enzyme-induced precipitation, centrifugation, or by EDTA-induced dissociation.

Figure 2.
Functions of milk-derived extracellular vesicles. Milk contains extracellular vesicles (EVs). (A) EVs in human milk expose tissue factor (TF), the protein that triggers blood clotting by binding and activating factor VII (FVII), thereby promoting haemostasis and wound healing. (B) Milk EVs may survive the gastrointestinal conditions of newborns, and such EVs are thought to participate in immune responses and homeostasis of the gastrointestinal tract. (C) Milk EVs attenuate virus infection with help from immune cells. (D) Milk EVs may kill harmful bacteria and promote the growth of beneficial bacteria within the gastrointestinal tract, which may have beneficial effects on growth and development of the newborn.
Figure 2.
Functions of milk-derived extracellular vesicles. Milk contains extracellular vesicles (EVs). (A) EVs in human milk expose tissue factor (TF), the protein that triggers blood clotting by binding and activating factor VII (FVII), thereby promoting haemostasis and wound healing. (B) Milk EVs may survive the gastrointestinal conditions of newborns, and such EVs are thought to participate in immune responses and homeostasis of the gastrointestinal tract. (C) Milk EVs attenuate virus infection with help from immune cells. (D) Milk EVs may kill harmful bacteria and promote the growth of beneficial bacteria within the gastrointestinal tract, which may have beneficial effects on growth and development of the newborn.




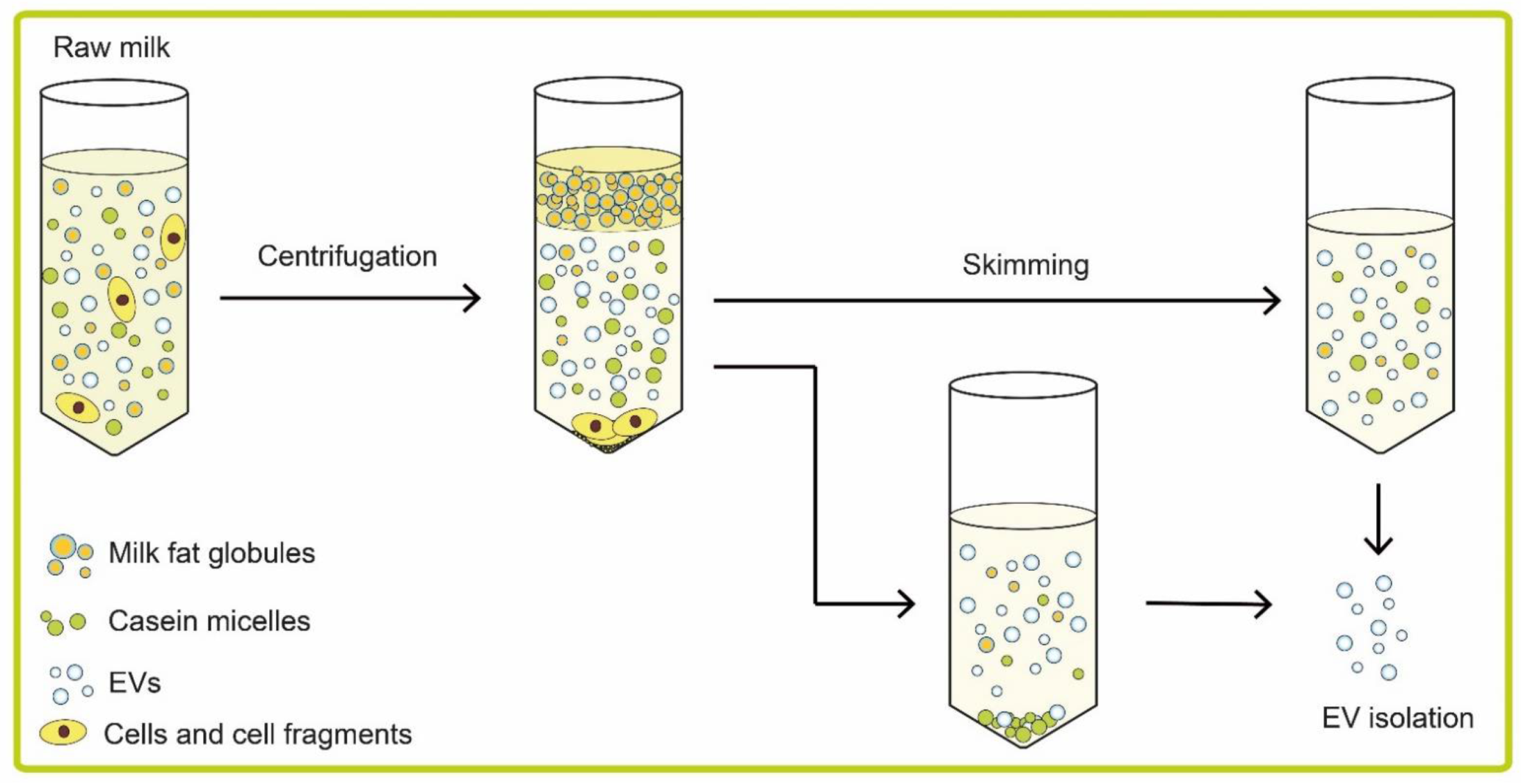{kind=link}
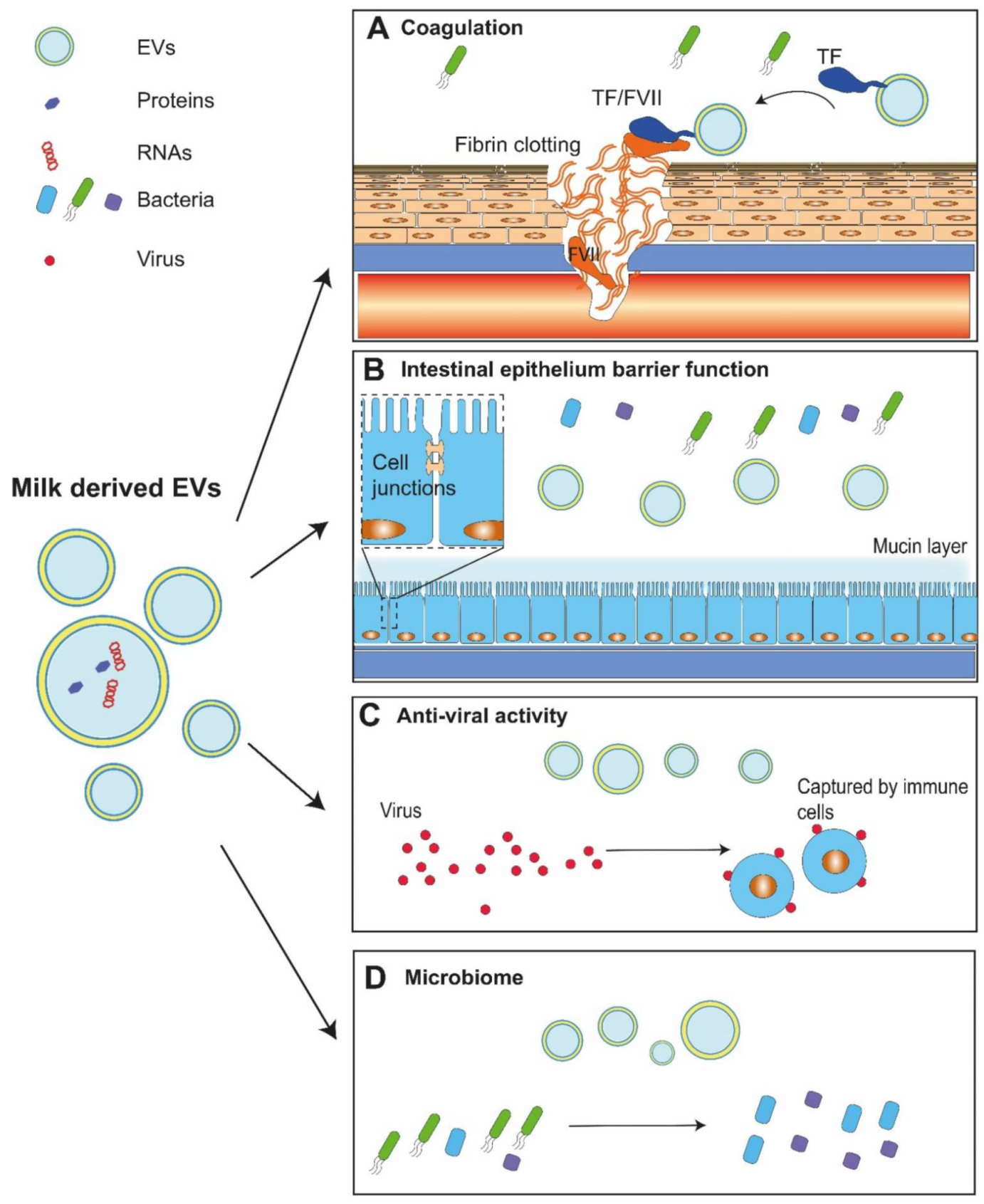{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hu, Y.; Thaler, J.; Nieuwland, R. Extracellular Vesicles in Human Milk. Pharmaceuticals 2021, 14, 1050. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101050
AMA Style
Hu Y, Thaler J, Nieuwland R. Extracellular Vesicles in Human Milk. Pharmaceuticals. 2021; 14(10):1050. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101050
Chicago/Turabian StyleHu, Yong, Johannes Thaler, and Rienk Nieuwland. 2021. "Extracellular Vesicles in Human Milk" Pharmaceuticals 14, no. 10: 1050. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101050
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.