
DAKS1, a Kunitz Scaffold Peptide from the Venom Gland of Deinagkistrodon acutus Prevents Carotid-Artery and Middle-Cerebral-Artery Thrombosis via Targeting Factor XIa

 , ,
, ,
Abstract
:1. Introduction
2. Results
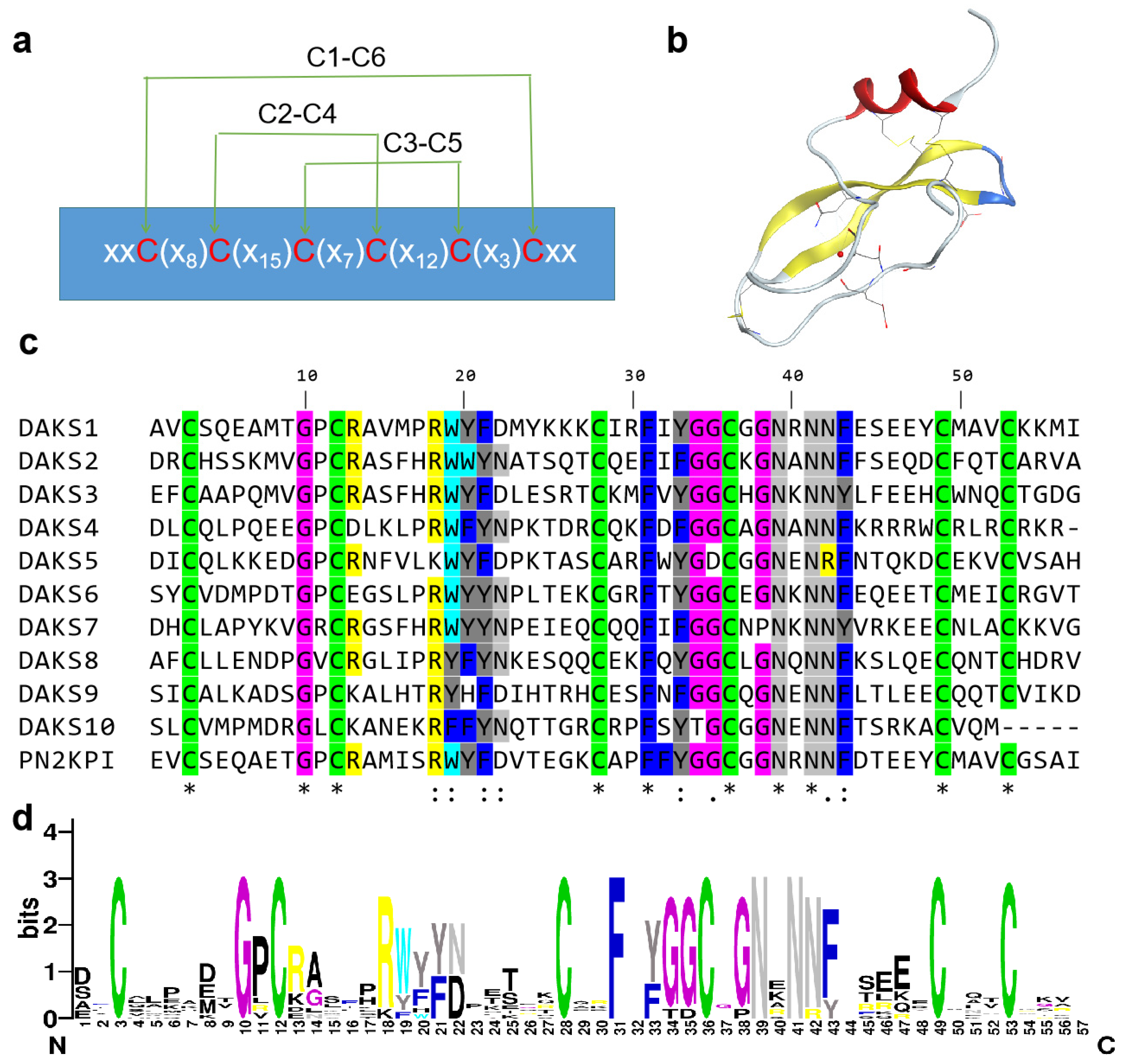
2.1. Ten Kunitz SBPs Discovered from Venom Gland of D. acutus
2.2. FXIa Inhibitory Activity of 10 Kunitz Peptides Tested by Substrate Chromogenic Method
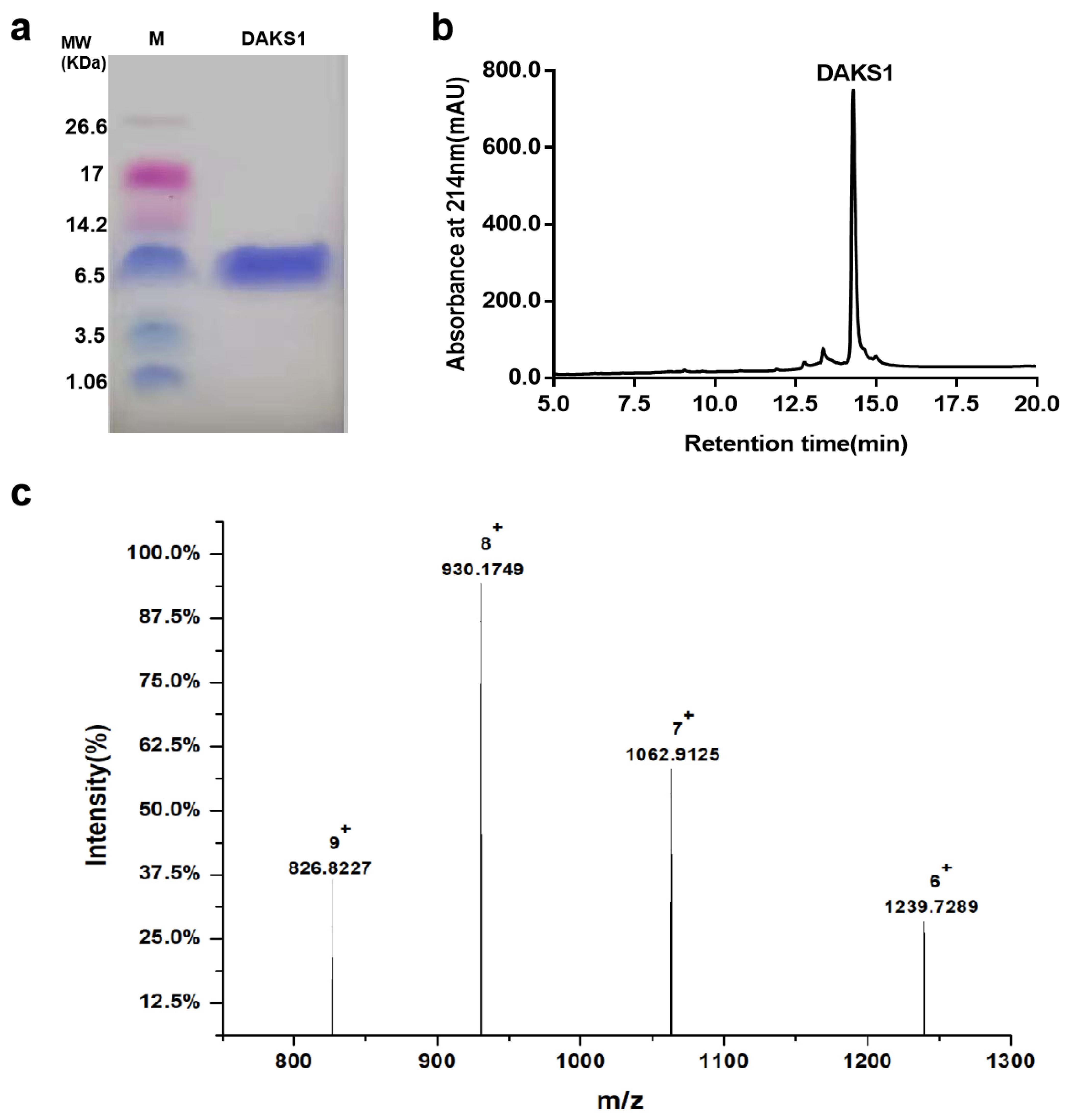
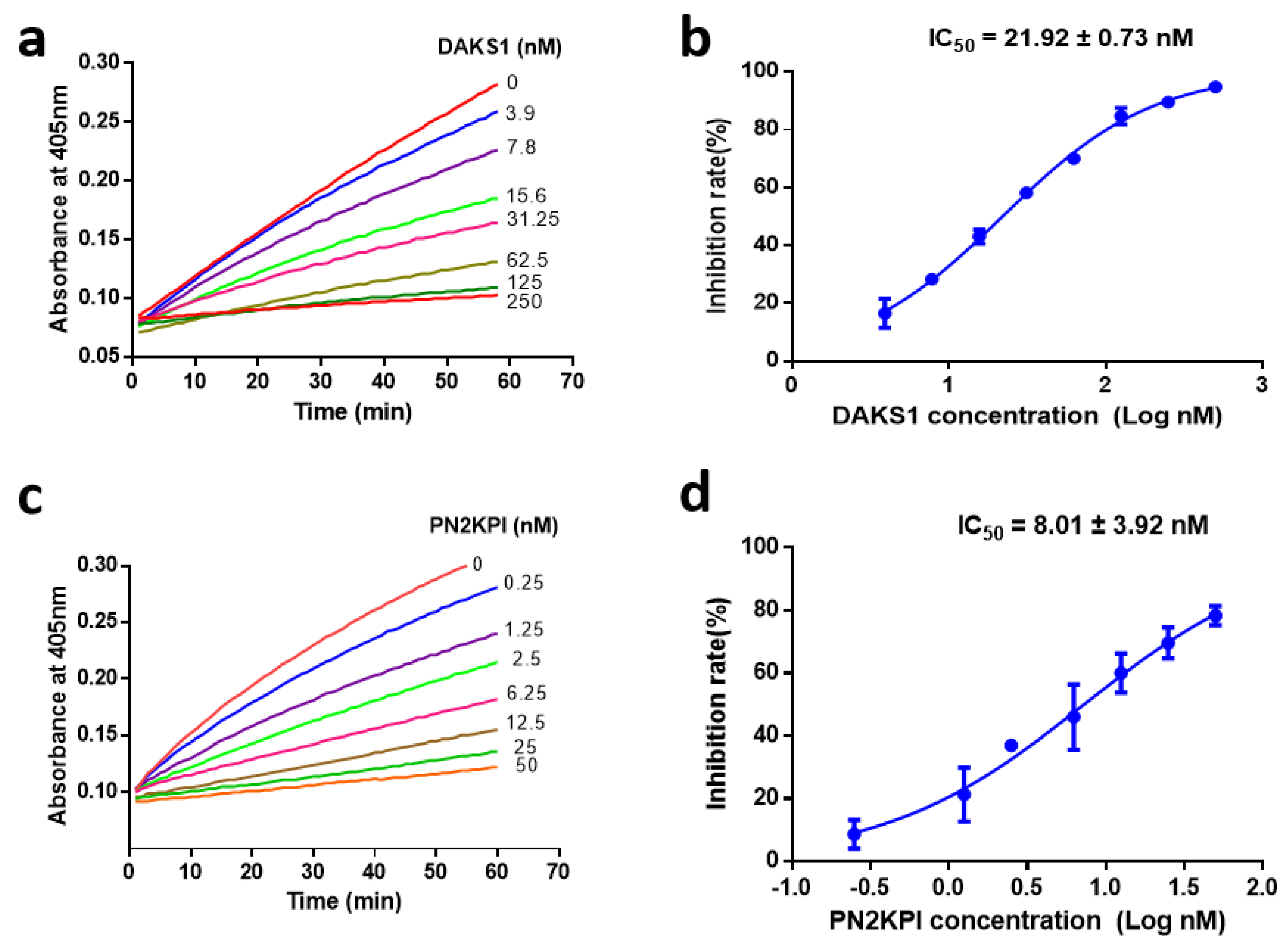
2.3. DAKS1 Is a Potent FXIa Inhibitor
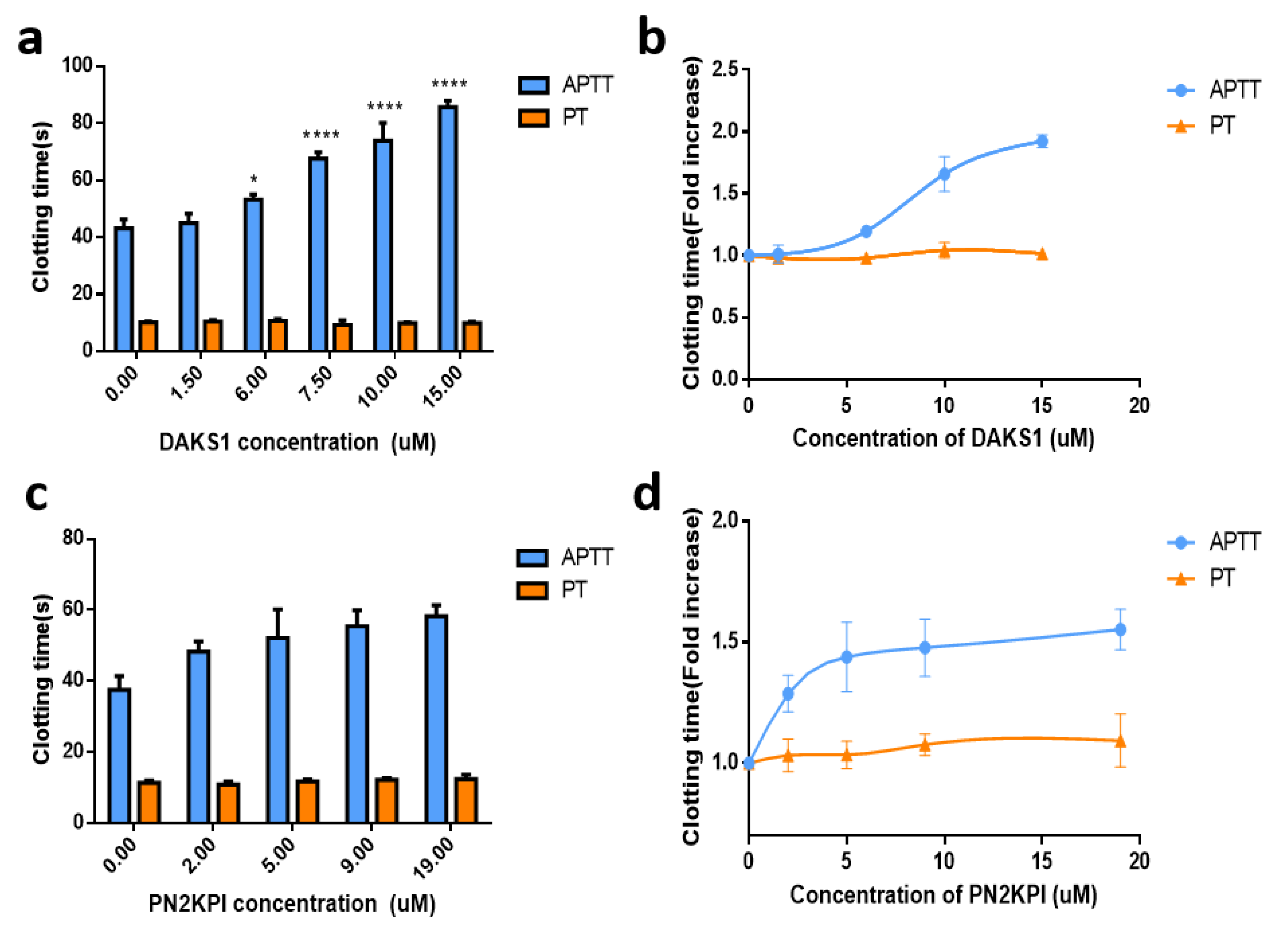
2.4. DAKS1 Prolonged APTT in Mouse Plasma
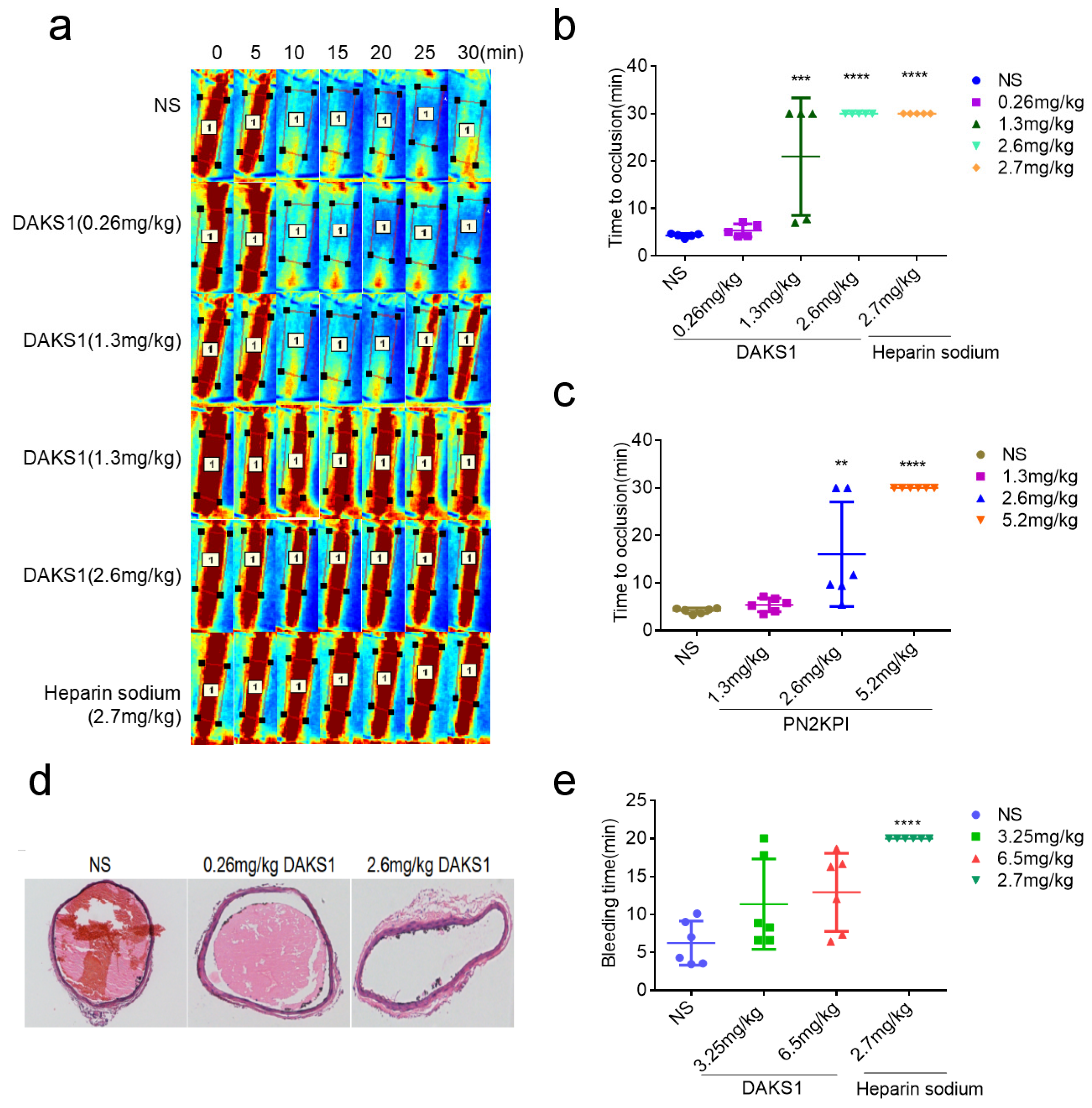
2.5. DAKS1 Inhibited Ferric Chloride-Induced Carotid-Artery Thrombosis in Mice
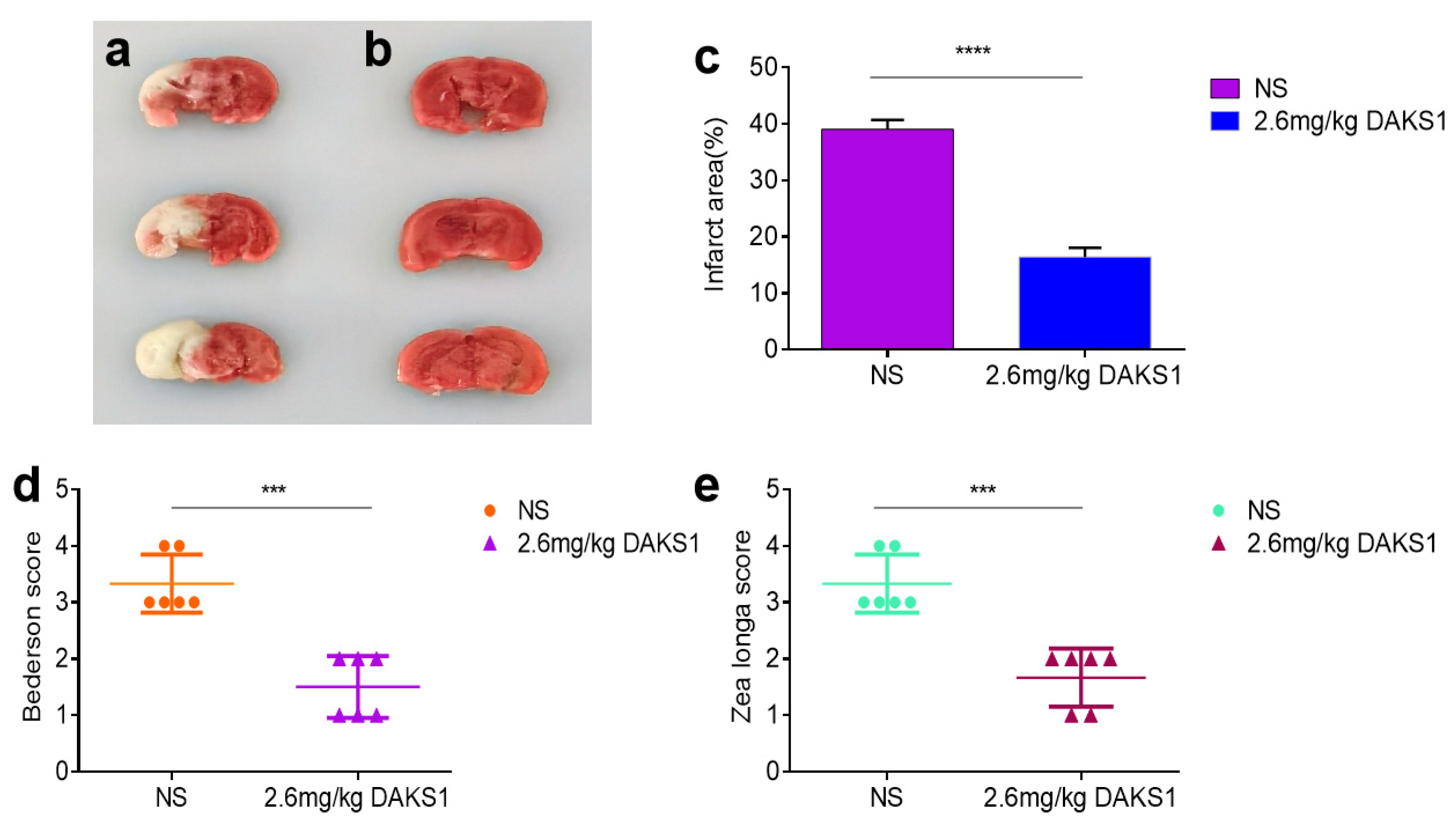
2.6. DAKS1 Inhibited Stroke in Mice
2.7. DAKS1 Did Not Show Bleeding Risk in Mice
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Animals Samples
4.3. Preparation of Venom Gland of D. acutus
4.4. Preparation of RNA Library and Sequencing
4.5. Bioinformatic Processing
4.6. Pfam Platform Recognizing
4.7. Expression and Purification of Kunitz Peptides
4.8. HPLC
4.9. LC–Q–TOF–MS Analysis
4.10. FXIa Inhibitory Activity Testing
4.11. Protease Selectivity Testing
4.12. APTT and PT
4.13. Antithrombotic Activity Assay
4.14. Transient Occlusion Model of Middle Cerebral Artery
4.15. Tail Bleeding-Time Assay
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gebauer, M.; Skerra, A. Engineered Protein Scaffolds as Next-Generation Therapeutics. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 391–415. [Google Scholar] [CrossRef] [PubMed]
- Simeon, R.; Chen, Z. In Vitro-Engineered Non-Antibody Protein Therapeutics. Protein Cell 2018, 9, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Lofblom, J.; Frejd, F.Y.; Stahl, S. Non-immunoglobulin Based Protein Scaffolds. Curr. Opin. Biotechnol. 2011, 22, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.S.; Deer, T.R. Safety and Efficacy of Intrathecal Ziconotide in the Management of Severe Chronic Pain. Ther. Clin. Risk Manag. 2009, 5, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Layer, P.; Stanghellini, V. Review Article: Linaclotide for the Management of Irritable Bowel Syndrome with Constipation. Aliment. Pharmacol. Ther. 2014, 39, 371–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez-Lombardi, R.; Phan, T.G.; Zimmermann, C.; Lowe, D.; Jermutus, L.; Christ, D. Challenges and Opportunities for Non-Antibody Scaffold Drugs. Drug Discov. Today 2015, 20, 1271–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, M. Evolutionary Aspects of the Structural Convergence and Functional Diversification of Kunitz-Domain Inhibitors. J. Mol. Evol. 2020, 88, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.; Abbenante, G.; Fairlie, D.P. Protease Inhibitors: Current Status and Future Prospects. J. Med. Chem. 2000, 43, 305–341. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, M.; Kato, I. Protein Inhibitors of Proteinases. Annu. Rev. Biochem. 1980, 49, 593–626. [Google Scholar] [CrossRef] [PubMed]
- Hosse, R.J.; Rothe, A.; Power, B.E. A New Generation of Protein Display Scaffolds for Molecular Recognition. Protein Sci. 2006, 15, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Levy, R.J.; Lumry, W.R.; McNeil, D.L.; Li, H.H.; Campion, M.; Horn, P.T.; Pullman, W.E. EDEMA4: A Phase 3, Double-Blind Study of Subcutaneous Ecallantide Treatment for Acute Attacks of Hereditary Angioedema. Ann. Allergy Asthma Immunol. 2010, 104, 523–529. [Google Scholar] [CrossRef]
- Schneider, L.; Lumry, W.; Vegh, A.; Williams, A.H.; Schmalbach, T. Critical Role of Kallikrein in Hereditary Angioedema Pathogenesis: A Clinical Trial of Ecallantide, a Novel Kallikrein Inhibitor. J. Allergy Clin. Immun. 2007, 120, 416–422. [Google Scholar] [CrossRef]
- Martello, J.L.; Woytowish, M.R.; Chambers, H. Ecallantide for Treatment of Acute Attacks of Hereditary Angioedema. Am. J. Health Syst. Pharm. 2012, 69, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Roberts, B.L.; Markland, W.; Ley, A.C.; Kent, R.B.; White, D.W.; Guterman, S.K.; Ladner, R.C. Directed Evolution of a Protein: Selection of Potent Neutrophil Elastase Inhibitors Displayed on M13 Fusion Phage. Proc. Natl. Acad. Sci. USA 1992, 89, 2429–2433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attucci, S.; Gauthier, A.; Korkmaz, B.; Delà Pine, P.; Martino, M.L.F.; Saudubray, F.O.; Diot, P.; Gauthier, F. EPI-hNE4, a Proteolysis-Resistant Inhibitor of Human Neutrophil Elastase and Potential Anti-Inflammatory Drug for Treating Cystic Fibrosis. J. Pharmacol. Exp. Ther. 2006, 318, 803–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlevy, F.K.; Martin, S.L.; de Courcey, F.; Elborn, J.S.; Ennis, M. Anti-Inflammatory Effects of DX-890, a Human Neutrophil Elastase Inhibitor. J. Cyst. Fibros. 2012, 11, 300–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, F.; Gailani, D.; Renné, T. Factor XI and XII As Antithrombotic Targets. Curr. Opin. Hematol. 2011, 18, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Salomon, O.; Steinberg, D.M.; Zucker, M.; Varon, D.; Zivelin, A.; Seligsohn, U. Patients with Severe Factor XI Deficiency Have a Reduced Incidence of Deep-Vein Thrombosis. Thromb. Haemost. 2011, 105, 269. [Google Scholar] [CrossRef]
- Salomon, O.; Steinberg, D.M.; Koren-Morag, N.; Tanne, D.; Seligsohn, U. Reduced Incidence of Ischemic Stroke in Patients with Severe Factor XI Deficiency. Blood 2008, 111, 4113–4117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, E.D.; Gailani, D.; Castellino, F.J. FXI Is Essential for Thrombus Formation Following FeCl3-Induced Injury of the Carotid Artery in the Mouse. Thromb. Haemost. 2002, 87, 774–776. [Google Scholar]
- Wang, X.; Cheng, Q.; Xu, L.; Feuerstein, G.Z.; Hsu, M.Y.; Smith, P.L.; Seiffert, D.A.; Schumacher, W.A.; Ogletree, M.L.; Gailani, D. Effects of Factor IX or Factor XI Deficiency on Ferric Chloride-Induced Carotid Artery Occlusion in Mice. J. Thromb. Haemost. 2005, 3, 695–702. [Google Scholar] [CrossRef]
- Wang, X.; Smith, P.L.; Hsu, M.Y.; Gailani, D.; Schumacher, W.A.; Ogletree, M.L.; Seiffert, D.A. Effects of Factor XI Deficiency on Ferric Chloride-Induced Vena Cava Thrombosis in Mice. J. Thromb. Haemost. 2006, 4, 1982–1988. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, H.; Navaneetham, D.; Reichenbach, Z.W.; Tuma, R.F.; Walsh, P.N. The Kunitz Protease Inhibitor Domain of Protease Nexin-2 Inhibits Factor XIa and Murine Carotid Artery and Middle Cerebral Artery Thrombosis. Blood 2012, 120, 671–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oltersdorf, T.; Fritz, L.C.; Schenk, D.B.; Lieberburg, I.; Johnson-Wood, K.L.; Beattie, E.C.; Ward, P.J.; Blacher, R.W.; Dovey, H.F.; Sinha, S. The Secreted Form of the Alzheimer’s Amyloid Precursor Protein with the Kunitz Domain Is Protease Nexin-II. Nature 1989, 341, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Carvalho, L.P.D.; Chan, M.Y.; Kini, R.M.; Kang, T.S. Fasxiator, a Novel Factor XIa Inhibitor from Snake Venom, and Its Site-Specific Mutagenesis to Improve Potency and Selectivity. J. Thromb. Haemost. 2015, 13, 248–261. [Google Scholar] [CrossRef]
- Decrem, Y.; Rath, G.; Blasioli, V.; Cauchie, P.; Robert, S.; Beaufays, J.; Frere, J.M.; Feron, O.; Dogne, J.M.; Dessy, C.; et al. Ir-CPI, a Coagulation Contact Phase Inhibitor from the Tick Ixodes ricinus, Inhibits Thrombus Formation without Impairing Hemostasis. J. Exp. Med. 2009, 206, 2381–2395. [Google Scholar] [CrossRef] [Green Version]
- Pireaux, V.; Tassignon, J.; Demoulin, S.; Derochette, S.; Borenstein, N.; Ente, A.; Fiette, L.; Douxfils, J.; Lancellotti, P.; Guyaux, M.; et al. Anticoagulation with an Inhibitor of Factors XIa and XIIa During Cardiopulmonary Bypass. J. Am. Coll. Cardiol. 2019, 74, 2178–2189. [Google Scholar] [CrossRef]
- De Oliveira, C.F.R.; Oliveira, C.T.; Taveira, G.B.; de Oliveira Mello, E.; Gomes, V.M.; Macedo, M.L.R. Characterization of a Kunitz Trypsin Inhibitor from Enterolobium timbouva with Activity against Candida Species. Int. J. Biol. Macromol. 2018, 119, 645–653. [Google Scholar] [CrossRef]
- Liao, Q.; Li, S.; Siu, S.W.I.; Yang, B.; Huang, C.; Chan, J.Y.; Morlighem, J.R.L.; Wong, C.T.T.; Rádis-Baptista, G.; Lee, S.M. Novel Kunitz-like Peptides Discovered in the Zoanthid Palythoa caribaeorum through Transcriptome Sequencing. J. Proteome Res. 2018, 17, 891–902. [Google Scholar] [CrossRef]
- Emsley, J.; McEwan, P.A.; Gailani, D. Structure and Function of Factor XI. Blood 2010, 115, 2569–2577. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Mizurini, D.M.; Assumpção, T.C.F.; Li, Y.; Qi, Y.; Kotsyfakis, M.; Ribeiro, J.M.C.; Monteiro, R.Q.; Francischetti, I.M.B. Desmolaris, a Novel Factor XIa Anticoagulant from the Salivary Gland of the Vampire Bat (Desmodus rotundus) Inhibits In-Flammation and Thrombosis In Vivo. Blood 2013, 122, 4094–4106. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.F.; Li, K.K.; Zhu, E.F.; Opel, C.F.; Kauke, M.J.; Kim, H.; Atolia, E.; Wittrup, K.D. Artificial Anti-Tumor Opsonizing Proteins with Fibronectin Scaffolds Engineered for Specificity to Each of the Murine FcgammaR Types. J. Mol. Biol. 2018, 430, 1786–1798. [Google Scholar] [CrossRef]
- Bloom, L.; Calabro, V. FN3: A New Protein Scaffold Reaches the Clinic. Drug Discov. Today 2009, 14, 949–955. [Google Scholar] [CrossRef]
- Wurch, T.; Pierre, A.; Depil, S. Novel Protein Scaffolds as Emerging Therapeutic Proteins: From Discovery to Clinical Proof-of-Concept. Trends Biotechnol. 2012, 30, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Owens, B. Faster, Deeper, Smaller–The Rise of Antibody-Like Scaffolds. Nat. Biotechnol. 2017, 35, 602–603. [Google Scholar] [CrossRef]
- Nuttall, S.D.; Walsh, R.B. Display Scaffolds: Protein Engineering for Novel Therapeutics. Curr. Opin. Pharmacol. 2008, 8, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Koide, A.; Bailey, C.W.; Huang, X.; Koide, S. The Fibronectin Type III Domain as a Scaffold for Novel Binding Proteins. J. Mol. Biol. 1998, 284, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Schiefner, A.; Skerra, A. The Menagerie of Human Lipocalins: A Natural Protein Scaffold for Molecular Recognition of Physiological Compounds. Acc. Chem. Res. 2015, 48, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Schlehuber, S.; Skerra, A. Lipocalins in Drug Discovery: From Natural Ligand-Binding Proteins to “Anticalins”. Drug Discov. Today 2005, 10, 23–33. [Google Scholar] [CrossRef]
- Morgenstern, D.; King, G.F. The Venom Optimization Hypothesis Revisited. Toxicon 2013, 63, 120–128. [Google Scholar] [CrossRef]
- Chen, M.; Ye, X.; Ming, X.; Chen, Y.; Wang, Y.; Su, X.; Su, W.; Kong, Y. A Novel Direct Factor Xa Inhibitory Peptide with Anti-Platelet Aggregation Activity from Agkistrodon acutus Venom Hydrolysates. Sci. Rep. 2015, 5, 10846. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Sun, Q.; Zhao, Q.; Zhang, Y. Purification and Characterization of a Novel Antiplatelet Peptide from Deinagkistrodon acutus Venom. Toxins 2018, 10, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Župunski, V.; Kordiš, D.; Gubenšek, F. Adaptive Evolution in the Snake Venom Kunitz/BPTI Protein Family. FEBS Lett. 2003, 547, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Rowan, E.G.; Harvey, A.L. Snake Toxins from Mamba Venoms: Unique Tools for the Physiologist. Acta Chim. Slov. 2011, 58, 689–692. [Google Scholar]
- Lehmann, A. Ecallantide (DX-88), a Plasma Kallikrein Inhibitor for the Treatment of Hereditary Angioedema and the Prevention of Blood Loss in On-Pump Cardiothoracic Surgery. Expert Opin. Biol. Ther. 2008, 8, 1187–1199. [Google Scholar] [CrossRef]
- Kanchongkittiphon, W.; Kabil, N.; Bacharier, L.B.; Kitcharoensakkul, M. Ecallantide: An Alternative Treatment of Refractory Angioedema in Adolescents with Systemic Lupus Erythematosus. J. Allergy Clin. Immunol. Pract. 2020, 8, 1115–1116. [Google Scholar] [CrossRef]
- Navaneetham, D.; Jin, L.; Pandey, P.; Strickler, J.E.; Babine, R.E.; Abdel-Meguid, S.S.; Walsh, P.N. Structural and Mutational Analyses of the Molecular Interactions between the Catalytic Domain of Factor XIa and the Kunitz Protease Inhibitor Domain of Protease Nexin 2. J. Biol. Chem. 2005, 280, 36165–36175. [Google Scholar] [CrossRef] [Green Version]
- Schaar, K.L.; Brenneman, M.M.; Savitz, S.I. Functional Assessments in the Rodent Stroke Model. Exp. Transl. Stroke Med. 2010, 2, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.N.; Zhang, G.F.; Chen, H.L.; Sun, X.P.; Qin, W.W.; Shi, F.; Sun, L.X.; Xu, X.N.; Wang, M.S. Selective Brain Hypothermia-Induced Neuroprotection against Focal Cerebral Ischemia/Reperfusion Injury Is Associated with Fis1 Inhibition. Neural Regen. Res. 2020, 15, 903–911. [Google Scholar]
- Tomaselli, G.F.; Mahaffey, K.W.; Cuker, A.; Dobesh, P.P.; Doherty, J.U.; Eikelboom, J.W.; Florido, R.; Hucker, W.; Mehran, R.; Messe, S.R.; et al. 2017 ACC Expert Consensus Decision Pathway on Management of Bleeding in Patients on Oral Anticoagulants: A Report of the American College of Cardiology Task Force on Expert Consensus Decision Pathways. J. Am. Coll. Cardiol. 2017, 70, 3042–3067. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.C.; Go, A.S.; Chang, Y.; Borowsky, L.H.; Pomernacki, N.K.; Udaltsova, N.; Singer, D.E. A New Risk Scheme to Predict Warfarin-Associated Hemorrhage: The ATRIA (Anticoagulation and Risk Factors in Atrial Fibrillation) Study. J. Am. Coll. Cardiol. 2011, 58, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paladin, L.; Tosatto, S.C. Comparison of Protein Repeat Classifications Based on Structure and Sequence Families. Biochem. Soc. Trans. 2015, 43, 832–837. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam Protein Families Database: Towards a More Sustainable Future. Nucleic Acids Res. 2016, 44, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Morowski, M.; Vögtle, T.; Kraft, P.; Kleinschnitz, C.; Stoll, G.; Nieswandt, B. Only Severe Thrombocytopenia Results in Bleeding and Defective Thrombus Formation in Mice. Blood 2013, 121, 4938–4947. [Google Scholar] [CrossRef] [Green Version]
- Krupka, J.; May, F.; Weimer, T.; Pragst, I.; Kleinschnitz, C.; Stoll, G.; Panousis, C.; Dickneite, G.; Nolte, M.W. The Coagulation Factor XIIa Inhibitor rHA-Infestin-4 Improves Outcome after Cerebral Ischemia/Reperfusion Injury in Rats. PLoS ONE 2016, 11, e0146783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bederson, J.B.; Pitts, L.H.; Tsuji, M.; Nishimura, M.C.; Davis, R.L.; Bartkowski, H. Rat Middle Cerebral Artery Occlusion: Evaluation of the Model and Development of a Neurologic Examination. Stroke 1986, 17, 472–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.W.; Yang, W.T.; Chen, S.; Xu, Q.Q.; Shan, C.S.; Zheng, G.Q.; Ruan, J.C. Neuroprotection of Catalpol for Experimental Acute Focal Ischemic Stroke: Preclinical Evidence and Possible Mechanisms of Antioxidation, Anti-Inflammation, and Antiapoptosis. Oxid. Med. Cell Longev. 2017, 2017, 5058609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, S.; Liu, Z.; Cao, Y.; Hua, Y.; Chen, C.; Guo, W.; Kong, Y. A Novel Protease-Activated Receptor 1 Inhibitor from the Leech Whitmania pigra. Chin. J. Nat. Med. 2019, 17, 591–599. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Name | Unigene_id | Sequences |
|---|---|---|
| DAKS1 | Unigene7405_D_acutus | AVCSQEAMTGPCRAVMPRWYFDMYKKKCIRFIYGGCGGNRNNFESEEYCMAVCKKMI |
| DAKS2 | Unigene2097_D_acutus_1 | DRCHSSKMVGPCRASFHRWWYNATSQTCQEFIFGGCKGNANNFFSEQDCFQTCARVA |
| DAKS3 | Unigene2097_D_acutus_2 | EFCAAPQMVGPCRASFHRWYFDLESRTCKMFVYGGCHGNKNNYLFEEHCWNQCTGDG |
| DAKS4 | Unigene24735_D_acutus_2 | DLCQLPQEEGPCDLKLPRWFYNPKTDRCQKFDFGGCAGNANNFKRRRWCRLRCRKR |
| DAKS5 | Unigene5437_D_acutus | DICQLKKEDGPCRNFVLKWYFDPKTASCARFWYGDCGGNENRFNTQKDCEKVCVSAH |
| DAKS6 | Unigene2946_D_acutus_2 | SYCVDMPDTGPCEGSLPRWYYNPLTEKCGRFTYGGCEGNKNNFEQEETCMEICRGVT |
| DAKS7 | Unigene2946_D_acutus_1 | DHCLAPYKVGRCRGSFHRWYYNPEIEQCQQFIFGGCNPNKNNYVRKEECNLACKKVG |
| DAKS8 | Unigene20491_D_acutus_2 | AFCLLENDPGVCRGLIPRYFYNKESQQCEKFQYGGCLGNQNNFKSLQECQNTCHDRV |
| DAKS9 | Unigene20491_D_acutus_1 | SICALKADSGPCKALHTRYHFDIHTRHCESFNFGGCQGNENNFLTLEECQQTCVIKD |
| DAKS10 | Unigene20491_D_acutus_3 | SLCVMPMDRGLCKANEKRFFYNQTTGRCRPFSYTGCGGNENNFTSRKACVQM |
| Peptide Name | Concentration (uM) | Inhibition Rate (%) |
|---|---|---|
| DAKS1 | 7.5 | 98.13 ± 0.31 |
| 0.15 | 91.20 ± 0.31 | |
| 0.05 | 63.51 ± 0.96 | |
| DAKS2 | 7.5 | 7.54 ± 0.34 |
| DAKS3 | 7.5 | 14.56 ± 0.45 |
| DAKS4 | 7.5 | 65.26 ± 1.10 |
| 0.15 | 9.82 ± 0.32 | |
| 0.05 | 4.54 ± 1.18 | |
| DAKS5 | 7.5 | −14.24 ± 1.08 |
| DAKS6 | 7.5 | −8.840 ± 0.75 |
| DAKS7 | 7.5 | 6.237 ± 1.92 |
| DAKS8 | 7.5 | −8.820 ± 2.12 |
| DAKS9 | 7.5 | 69.07 ± 2.08 |
| 0.15 | 27.16 ± 0.98 | |
| 0.05 | 16.17 ± 1.28 | |
| DAKS10 | 7.5 | −10.03 ± 1.24 |
| Inhibitor | Molar Ratio | Inhibition, % |
|---|---|---|
| DAKS1 | 1:125, FXIa:inhibitor | >70 |
| PN2KPI | 1:13.3, FXIa:inhibitor | >90 |
| DAKS1 | 1:6000, FXa:inhibitor | 27.45 |
| PN2KPI | 1:504, FXa:inhibitor | 55.84 |
| DAKS1 | 1:6000, thrombin:inhibtor | 1.54 |
| PN2KPI | 1:2066, thrombin:inhibtor | −39.51 |
| DAKS1 | 1:3000, kallikrein:inhibitor | 62.28 |
| PN2KPI | 1:206, kallikrein:inhibitor | 16.65 |
| DAKS1 | 1:500, trypsin:inhibitor | 68.46 |
| PN2KPI | 1:52, trypsin:inhibitor | 87.41 |
| DAKS1 | 1:6000, FXIIa:inhibitor | 0 |
| PN2KPI | 1:1033, FXIIa:inhibitor | 19.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Z.; Liu, Y.; Ji, X.; Zheng, Y.; Li, Z.; Jiang, S.; Li, H.; Kong, Y. DAKS1, a Kunitz Scaffold Peptide from the Venom Gland of Deinagkistrodon acutus Prevents Carotid-Artery and Middle-Cerebral-Artery Thrombosis via Targeting Factor XIa. Pharmaceuticals 2021, 14, 966. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14100966
Jia Z, Liu Y, Ji X, Zheng Y, Li Z, Jiang S, Li H, Kong Y. DAKS1, a Kunitz Scaffold Peptide from the Venom Gland of Deinagkistrodon acutus Prevents Carotid-Artery and Middle-Cerebral-Artery Thrombosis via Targeting Factor XIa. Pharmaceuticals. 2021; 14(10):966. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14100966
Chicago/Turabian StyleJia, Zhiping, Yunyang Liu, Xiaoru Ji, Yizheng Zheng, Zhengyang Li, Shuai Jiang, Hongjin Li, and Yi Kong. 2021. "DAKS1, a Kunitz Scaffold Peptide from the Venom Gland of Deinagkistrodon acutus Prevents Carotid-Artery and Middle-Cerebral-Artery Thrombosis via Targeting Factor XIa" Pharmaceuticals 14, no. 10: 966. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14100966