
The Combination of AHCC and ETAS Decreases Migration of Colorectal Cancer Cells, and Reduces the Expression of LGR5 and Notch1 Genes in Cancer Stem Cells: A Novel Potential Approach for Integrative Medicine

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
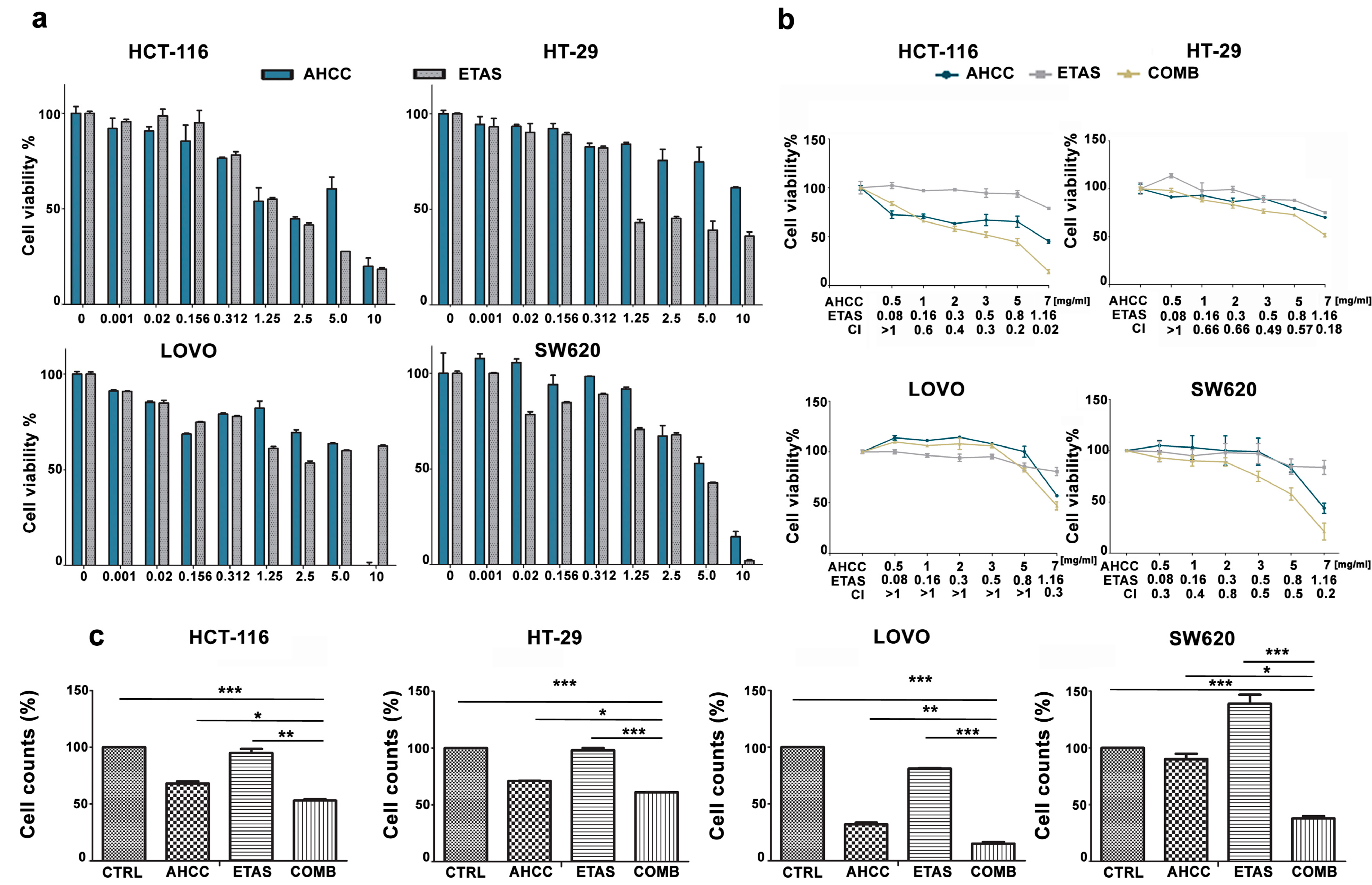
2.1. The Combination of AHCC and ETAS Is Synergistic and Inhibits Cell Growth in Colon Cancer Cells
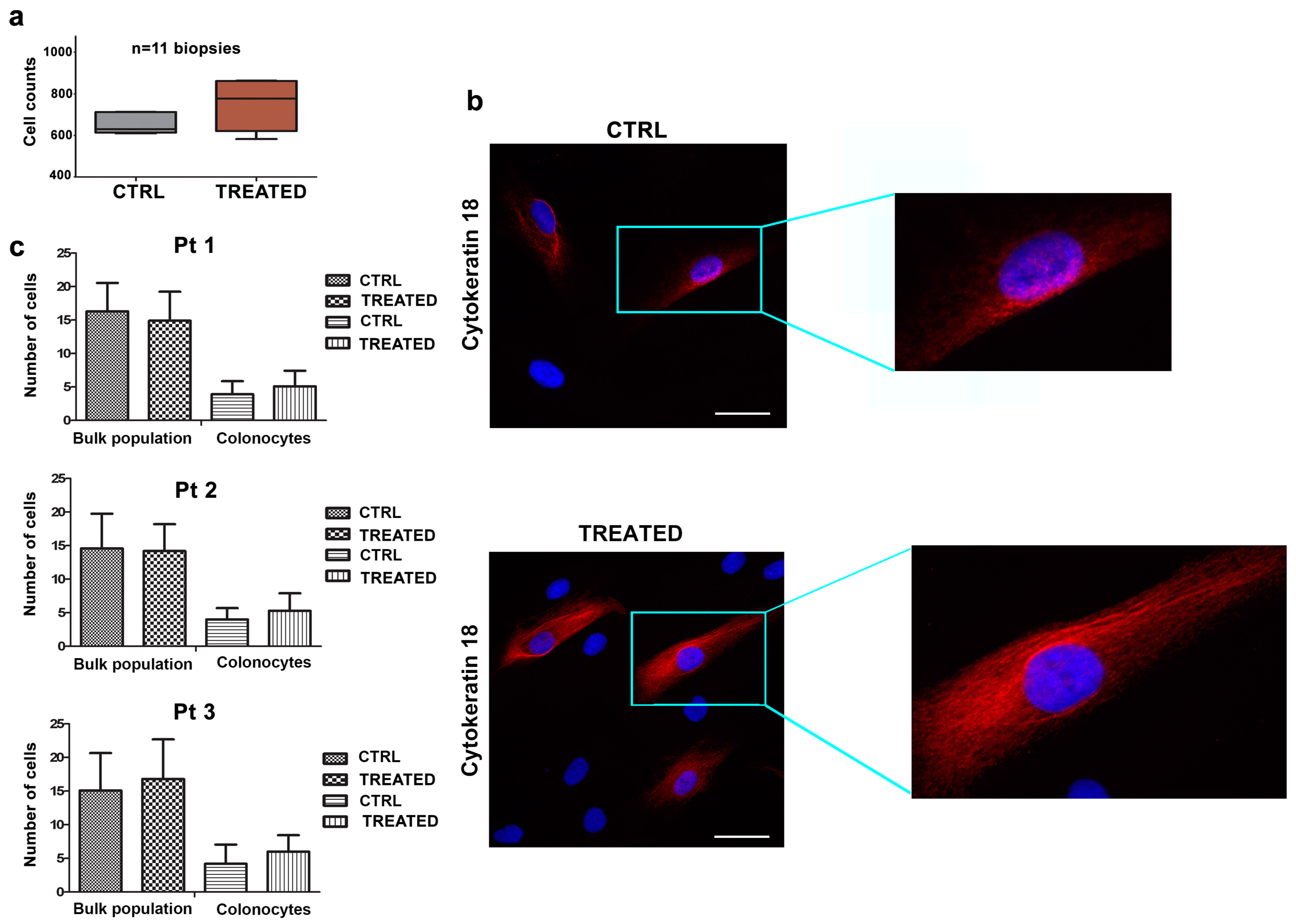
2.2. The Combination of AHCC and ETAS Does Not Affect Viability of Human Colonocytes from Healthy Donors
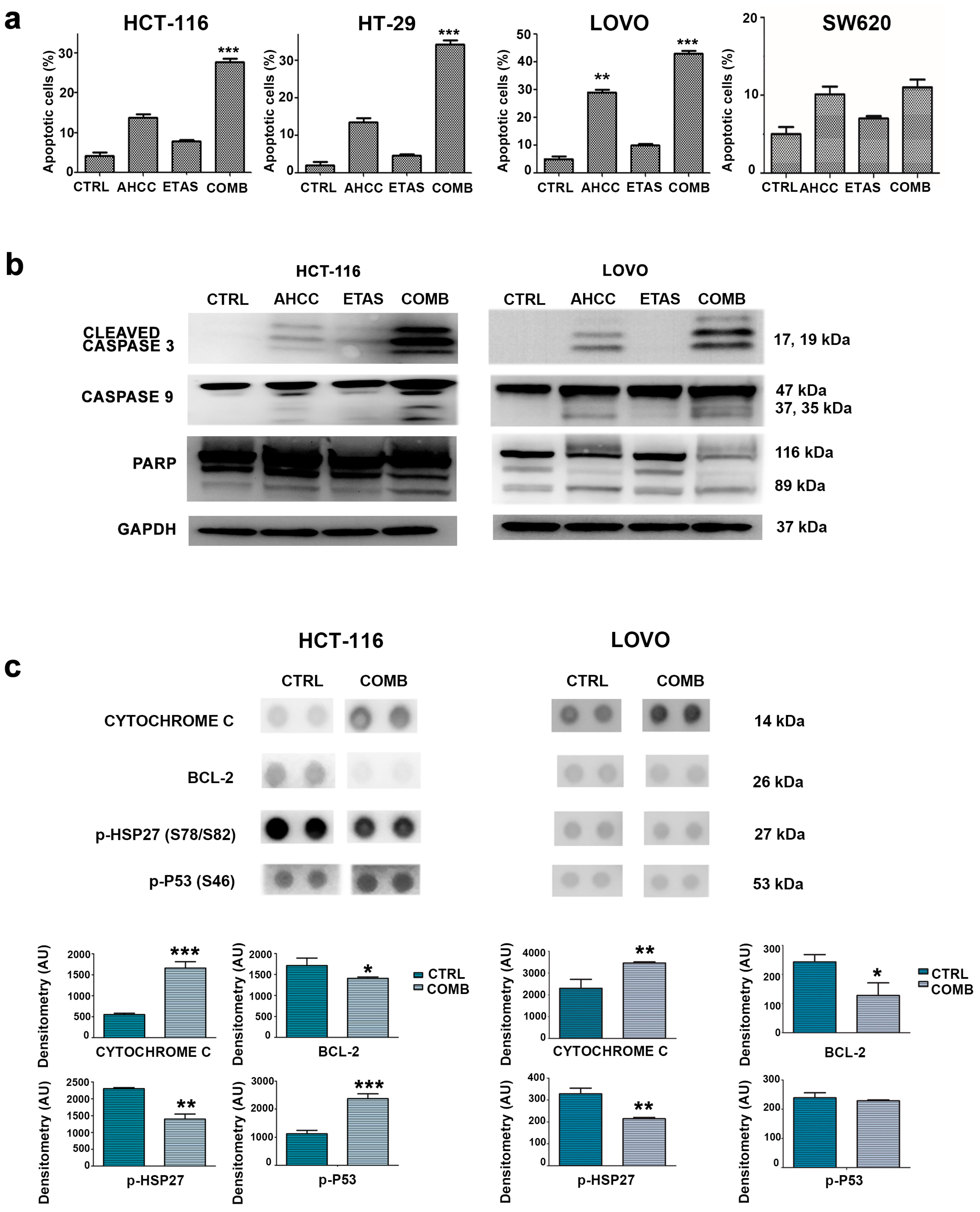
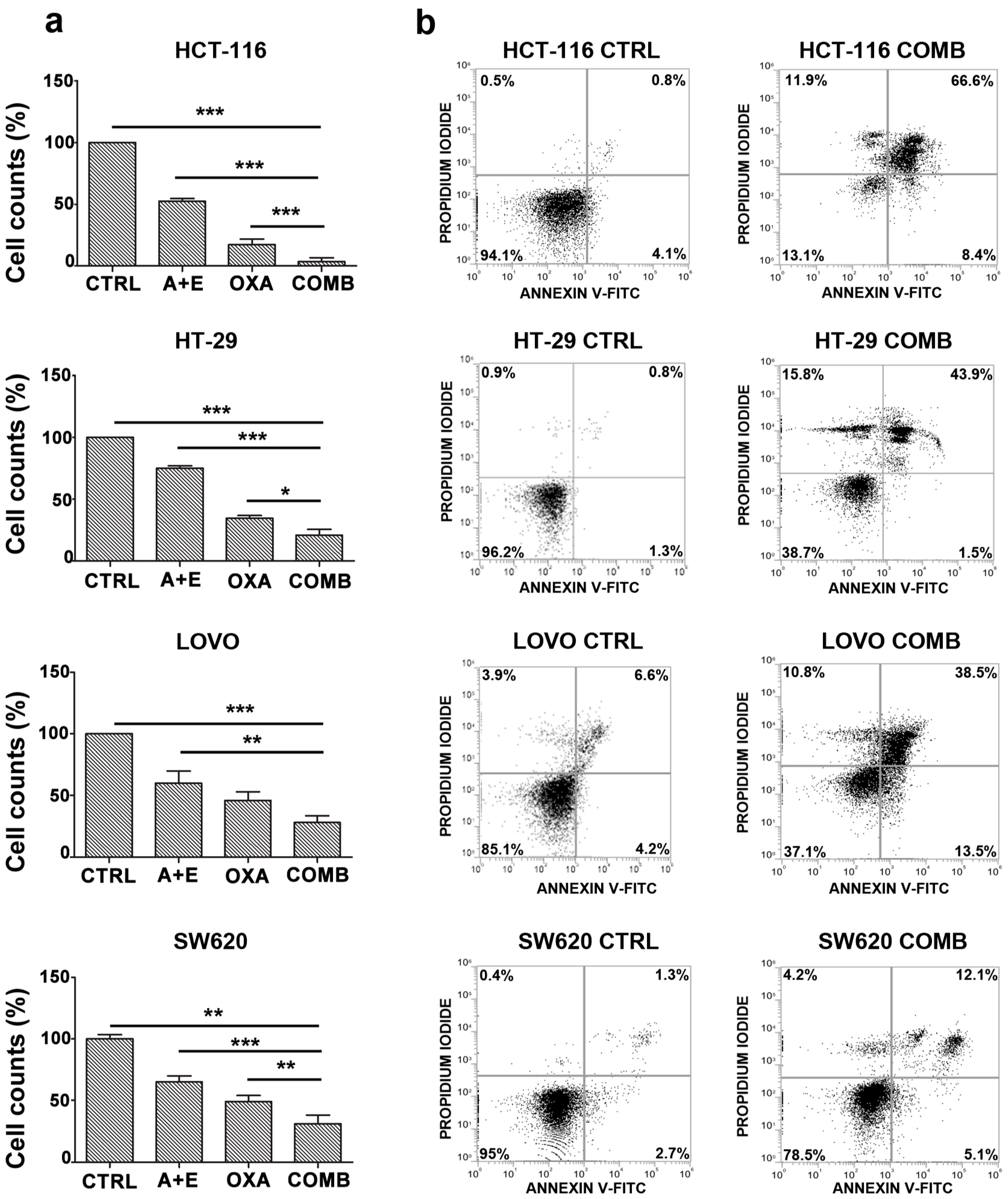
2.3. The Combination of AHCC and ETAS Promotes a Significant Increase in Cell Apoptosis in Colon Cancer Cells
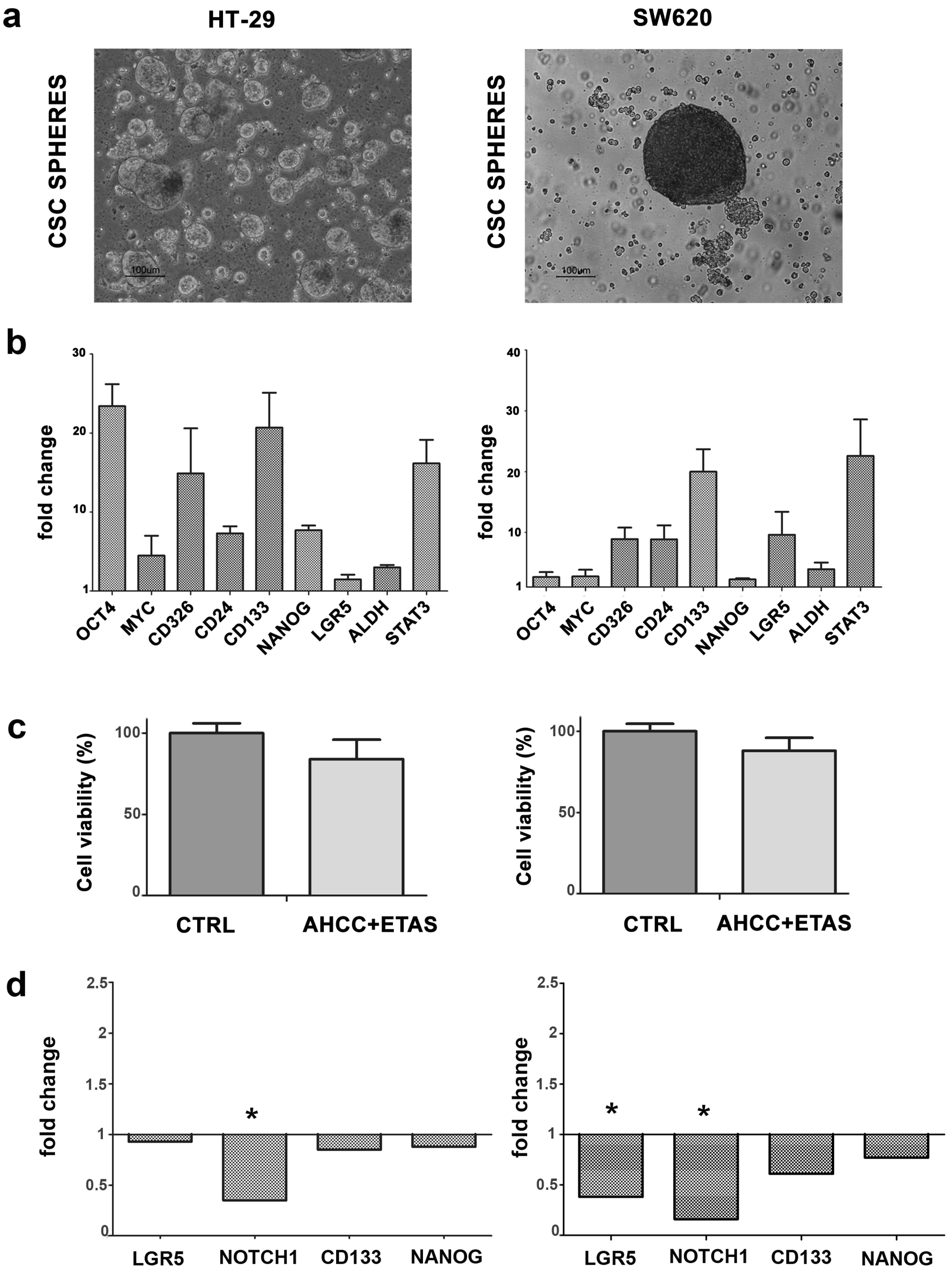
2.4. CSC Tumorspheres from CRC Cell Lines Are Affected by AHCC and ETAS Combination
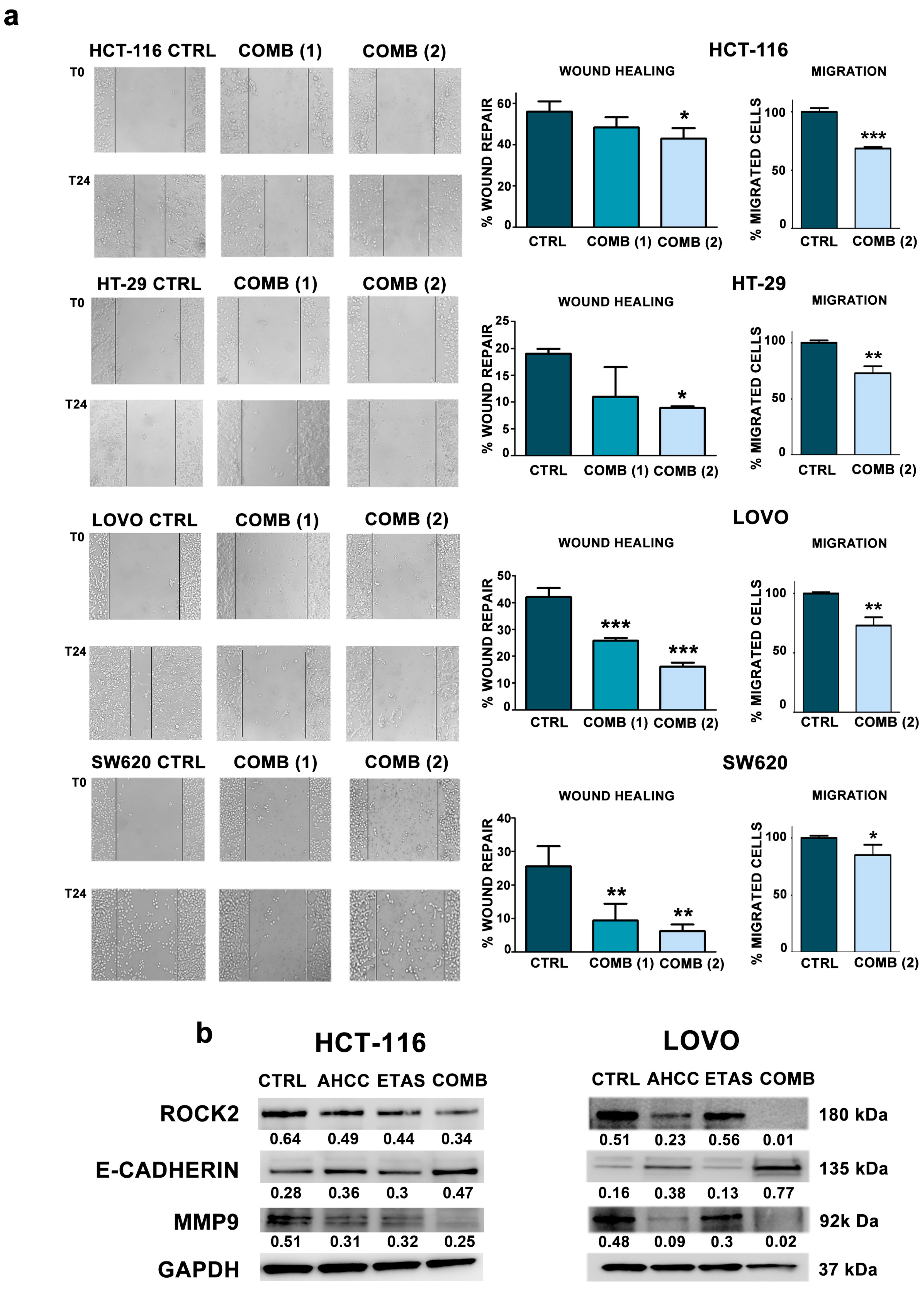
2.5. The Combination of AHCC and ETAS Reduces Cell Motility and Migration in Colon Cancer Cells
2.6. AHCC and ETAS Combination Enhances Oxaliplatinum Effects in Colon Cancer Cells
3. Discussion
4. Materials and Methods
4.1. AHCC and ETAS Preparation
4.2. Cell Line Cultures and Treatments
4.3. Human Tissue Collection and Primary Cell Culture
4.4. Cell Viability Analysis in Cell Lines
4.5. Treatments of Primary Cells from Biopsies
4.6. Isolation of Cancer Stem Cells from Tumor Cell Lines
4.7. Annexin V-FITC/PI Staining
4.8. Immunofluorescence Microscopy
4.9. Western Blotting Analyses
4.10. Wound Healing Assay
4.11. Transwell Migration Assay
4.12. Proteome Profiler
4.13. CSCs Treatment
4.14. RNA Extraction, Reverse Transcription, and Quantitative RT-PCR
4.15. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, R.A.; Asmis, T.R. Overview of systemic therapy for colorectal cancer. Clin. Colon. Rectal. Surg. 2009, 22, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Filip, S.; Vymetalkova, V.; Petera, J.; Vodickova, L.; Kubecek, O.; John, S.; Cecka, F.; Krupova, M.; Manethova, M.; Cervena, K.; et al. Distant Metastasis in Colorectal Cancer Patients-Do We Have New Predicting Clinicopathological and Molecular Biomarkers? A Comprehensive Review. Int. J. Mol. Sci. 2020, 21, 5255. [Google Scholar] [CrossRef]
- Rosenthal, D.S.; Dean-Clower, E. Integrative medicine in hematology/oncology: Benefits, ethical considerations, and controversies. Hematology 2005, 2005, 491–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelis, F.H.; Najdawi, M.; Ammar, M.B.; Nouri-Neuville, M.; Lombart, B.; Lotz, J.P.; Cadranel, J.; Barral, M. Integrative Medicine in Interventional Oncology: A Virtuous Alliance. Medicina 2020, 56, 35. [Google Scholar] [CrossRef] [Green Version]
- Greenwell, M.; Rahman, P.K. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar] [CrossRef]
- Balachandran, P.; Govindarajan, R. Cancer—An ayurvedic perspective. Pharmacol. Res. 2005, 51, 19–30. [Google Scholar] [CrossRef]
- Shin, M.S.; Park, H.J.; Maeda, T.; Nishioka, H.; Fujii, H.; Kang, I. The Effects of AHCC(R), a Standardized Extract of Cultured Lentinura edodes Mycelia, on Natural Killer and T Cells in Health and Disease: Reviews on Human and Animal Studies. J. Immunol. Res. 2019, 2019, 3758576. [Google Scholar] [CrossRef] [Green Version]
- Masuda, Y.; Nakayama, Y.; Tanaka, A.; Naito, K.; Konishi, M. Antitumor activity of orally administered maitake alpha-glucan by stimulating antitumor immune response in murine tumor. PLoS ONE 2017, 12, e0173621. [Google Scholar] [CrossRef]
- Hoshi, H.; Yagi, Y.; Iijima, H.; Matsunaga, K.; Ishihara, Y.; Yasuhara, T. Isolation and characterization of a novel immunomodulatory alpha-glucan-protein complex from the mycelium of Tricholoma matsutake in basidiomycetes. J. Agric. Food Chem. 2005, 53, 8948–8956. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, D.; Sun, B.; Fujii, H.; Kosuna, K.; Yin, Z. Active hexose correlated compound enhances tumor surveillance through regulating both innate and adaptive immune responses. Cancer Immunol. Immunother. 2006, 55, 1258–1266. [Google Scholar] [CrossRef]
- Choi, J.Y.; Lee, S.; Yun, S.M.; Suh, D.H.; Kim, K.; No, J.H.; Jeong, E.H.; Kim, Y.B. Active Hexose Correlated Compound (AHCC) Inhibits the Proliferation of Ovarian Cancer Cells by Suppressing Signal Transducer and Activator of Transcription 3 (STAT3) Activation. Nutr. Cancer 2018, 70, 109–115. [Google Scholar] [CrossRef]
- Takahashi, M.; Fujii, G.; Hamoya, T.; Kurokawa, Y.; Matsuzawa, Y.; Miki, K.; Komiya, M.; Narita, T.; Mutoh, M. Activation of NF-E2 p45-related factor-2 transcription and inhibition of intestinal tumor development by AHCC, a standardized extract of cultured Lentinula edodes mycelia. J. Clin. Biochem. Nutr. 2019, 65, 203–208. [Google Scholar] [CrossRef]
- Tokunaga, M.; Baron, B.; Kitagawa, T.; Tokuda, K.; Kuramitsu, Y. Active Hexose-correlated Compound Down-regulates Heat Shock Factor 1, a Transcription Factor for HSP27, in Gemcitabine-resistant Human Pancreatic Cancer Cells. Anticancer Res. 2015, 35, 6063–6067. [Google Scholar] [PubMed]
- Hangai, S.; Iwase, S.; Kawaguchi, T.; Kogure, Y.; Miyaji, T.; Matsunaga, T.; Nagumo, Y.; Yamaguchi, T. Effect of active hexose-correlated compound in women receiving adjuvant chemotherapy for breast cancer: A retrospective study. J. Altern. Complement. Med. 2013, 19, 905–910. [Google Scholar] [CrossRef] [Green Version]
- Hirose, A.; Sato, E.; Fujii, H.; Sun, B.; Nishioka, H.; Aruoma, O.I. The influence of active hexose correlated compound (AHCC) on cisplatin-evoked chemotherapeutic and side effects in tumor-bearing mice. Toxicol. Appl. Pharmacol. 2007, 222, 152–158. [Google Scholar] [CrossRef]
- Shigama, K.; Nakaya, A.; Wakame, K.; Nishioka, H.; Fujii, H. Alleviating effect of active hexose correlated compound (AHCC) for anticancer drug-induced side effects in non-tumor-bearing mice. J. Exp. Ther. Oncol. 2009, 8, 43–51. [Google Scholar]
- Sun, B.; Wakame, K.; Sato, E.; Nishioka, H.; Aruoma, O.I.; Fujii, H. The effect of active hexose correlated compound in modulating cytosine arabinoside-induced hair loss, and 6-mercaptopurine- and methotrexate-induced liver injury in rodents. Cancer Epidemiol. 2009, 33, 293–299. [Google Scholar] [CrossRef]
- Ye, S.F.; Ichimura, K.; Wakame, K.; Ohe, M. Suppressive effects of Active Hexose Correlated Compound on the increased activity of hepatic and renal ornithine decarboxylase induced by oxidative stress. Life Sci. 2003, 74, 593–602. [Google Scholar] [CrossRef]
- Aviles, H.; O’Donnell, P.; Orshal, J.; Fujii, H.; Sun, B.; Sonnenfeld, G. Active hexose correlated compound activates immune function to decrease bacterial load in a murine model of intramuscular infection. Am. J. Surg. 2008, 195, 537–545. [Google Scholar] [CrossRef]
- Roman, B.E.; Beli, E.; Duriancik, D.M.; Gardner, E.M. Short-term supplementation with active hexose correlated compound improves the antibody response to influenza B vaccine. Nutr. Res. 2013, 33, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Ono, T.; Sato, A.; Goto, K.; Miura, T.; Wakame, K.; Nishioka, H.; Maeda, T. Toxicological assessment of enzyme-treated asparagus extract in rat acute and subchronic oral toxicity studies and genotoxicity tests. Regul. Toxicol. Pharmacol. 2014, 68, 240–249. [Google Scholar] [CrossRef]
- Shimada, T.; Nanimoto, Y.; Baron, B.; Kitagawa, T.; Tokuda, K.; Kuramitsu, Y. Enzyme-treated Asparagus Extract Down-regulates Heat Shock Protein 27 of Pancreatic Cancer Cells. In Vivo 2018, 32, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.C.; Wu, C.S.; Wu, T.C.; Lin, Y.H.; Chang, S.J. A Standardized Extract of Asparagus officinalis Stem (ETAS((R))) Ameliorates Cognitive Impairment, Inhibits Amyloid beta Deposition via BACE-1 and Normalizes Circadian Rhythm Signaling via MT1 and MT2. Nutrients 2019, 11, 1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, T.; Ito, T.; Wakame, K.; Kitadate, K.; Arai, T.; Ogasawara, J.; Kizaki, T.; Sato, S.; Ishibashi, Y.; Fujiwara, T.; et al. Enzyme-treated Asparagus officinalis extract shows neuroprotective effects and attenuates cognitive impairment in senescence-accelerated mice. Nat. Prod. Commun. 2014, 9, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirato, K.; Takanari, J.; Sakurai, T.; Ogasawara, J.; Imaizumi, K.; Ohno, H. Enzyme-Treated Asparagus Extract Prevents’Hydrogen Peroxide-Induced Pro-Inflammatory Responses by Suppressing p65 Nuclear Translocation in Skin L929 Fibroblasts. Nat. Prod. Commun. 2016, 11, 1883–1888. [Google Scholar] [PubMed]
- Shirato, K.; Takanari, J.; Ogasawara, J.; Sakurai, T.; Imaizumi, K.; Ohno, H.; Kizaki, T. Enzyme-Treated Asparagus Extract Attenuates Hydrogen Peroxide-Induced Matrix Metalloproteinase-9 Expression in Murine Skin Fibroblast L929 Cells. Nat. Prod. Commun. 2016, 11, 677–680. [Google Scholar] [CrossRef] [Green Version]
- Oda, K.; Arakawa, H.; Tanaka, T.; Matsuda, K.; Tanikawa, C.; Mori, T.; Nishimori, H.; Tamai, K.; Tokino, T.; Nakamura, Y.; et al. p53AIP1, a potential mediator of p53-dependent apoptosis, and its regulation by Ser-46-phosphorylated p53. Cell 2000, 102, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.M.; Ranganathan, V.; Farnsworth, M.L.; Kavallaris, M.; Lock, R.B. Bcl-2 inhibits Bax translocation from cytosol to mitochondria during drug-induced apoptosis of human tumor cells. Cell Death Differ. 2000, 7, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Rouse, J.; Cohen, P.; Trigon, S.; Morange, M.; Alonso-Llamazares, A.; Zamanillo, D.; Hunt, T.; Nebreda, A.R. A novel kinase cascade triggered by stress and heat shock that stimulates MAPKAP kinase-2 and phosphorylation of the small heat shock proteins. Cell 1994, 78, 1027–1037. [Google Scholar] [CrossRef]
- Concannon, C.G.; Gorman, A.M.; Samali, A. On the role of Hsp27 in regulating apoptosis. Apoptosis 2003, 8, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Kang, K.W.; Kim, K.H.; Kwon, B.; Kim, S.K.; Lee, H.Y.; Kong, S.Y.; Lee, E.S.; Jang, S.G.; Yoo, B.C. Upregulated HSP27 in human breast cancer cells reduces Herceptin susceptibility by increasing Her2 protein stability. BMC Cancer 2008, 8, 286. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.S.; Lin, J.H.; Huang, W.C.; Hsu, T.W.; Su, K.; Chiou, S.H.; Tsai, Y.T.; Hung, S.C. Chemoresistance of lung cancer stemlike cells depends on activation of Hsp27. Cancer 2011, 117, 1516–1528. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Bao, Y.; Yang, G.K.; Wan, J.; Du, L.J.; Ma, Z.H. MiR-214 sensitizes human colon cancer cells to 5-FU by targeting Hsp27. Cell Mol. Biol. Lett. 2019, 24, 22. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, S.; Kuramitsu, Y.; Kaino, S.; Maehara, S.; Maehara, Y.; Sakaida, I.; Nakamura, K. Active hexose-correlated compound down-regulates HSP27 of pancreatic cancer cells, and helps the cytotoxic effect of gemcitabine. Anticancer Res. 2014, 34, 141–146. [Google Scholar]
- Kanwar, S.S.; Yu, Y.; Nautiyal, J.; Patel, B.B.; Majumdar, A.P. The Wnt/beta-catenin pathway regulates growth and maintenance of colonospheres. Mol. Cancer 2010, 9, 212. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Vishnubhotla, R.; Sun, S.; Huq, J.; Bulic, M.; Ramesh, A.; Guzman, G.; Cho, M.; Glover, S.C. ROCK-II mediates colon cancer invasion via regulation of MMP-2 and MMP-13 at the site of invadopodia as revealed by multiphoton imaging. Lab Investig. 2007, 87, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Lane, J.; Martin, T.A.; Watkins, G.; Mansel, R.E.; Jiang, W.G. The expression and prognostic value of ROCK I and ROCK II and their role in human breast cancer. Int. J. Oncol. 2008, 33, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Kamai, T.; Tsujii, T.; Arai, K.; Takagi, K.; Asami, H.; Ito, Y.; Oshima, H. Significant association of Rho/ROCK pathway with invasion and metastasis of bladder cancer. Clin. Cancer Res. 2003, 9, 2632–2641. [Google Scholar]
- Said, A.H.; Raufman, J.P.; Xie, G. The role of matrix metalloproteinases in colorectal cancer. Cancers 2014, 6, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Dragutinovic, V.V.; Radonjic, N.V.; Petronijevic, N.D.; Tatic, S.B.; Dimitrijevic, I.B.; Radovanovic, N.S.; Krivokapic, Z.V. Matrix metalloproteinase-2 (MMP-2) and -9 (MMP-9) in preoperative serum as independent prognostic markers in patients with colorectal cancer. Mol. Cell Biochem. 2011, 355, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Hazan, R.B.; Qiao, R.; Keren, R.; Badano, I.; Suyama, K. Cadherin switch in tumor progression. Ann. N. Y. Acad. Sci. 2004, 1014, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Bu, X.; Chen, H.; Wang, Q.; Sha, W. Bmi-1 promotes the invasion and migration of colon cancer stem cells through the downregulation of E-cadherin. Int. J. Mol. Med. 2016, 38, 1199–1207. [Google Scholar] [CrossRef] [Green Version]
- Cronin, K.A.; Lake, A.J.; Scott, S.; Sherman, R.L.; Noone, A.M.; Howlader, N.; Henley, S.J.; Anderson, R.N.; Firth, A.U.; Ma, J.; et al. Annual Report to the Nation on the Status of Cancer, part I: National cancer statistics. Cancer 2018, 124, 2785–2800. [Google Scholar] [CrossRef] [Green Version]
- Van der Jeught, K.; Xu, H.C.; Li, Y.J.; Lu, X.B.; Ji, G. Drug resistance and new therapies in colorectal cancer. World J. Gastroenterol. 2018, 24, 3834–3848. [Google Scholar] [CrossRef]
- Spierings, E.L.; Fujii, H.; Sun, B.; Walshe, T. A Phase I study of the safety of the nutritional supplement, active hexose correlated compound, AHCC, in healthy volunteers. J. Nutr. Sci. Vitaminol. 2007, 53, 536–539. [Google Scholar] [CrossRef] [Green Version]
- Terakawa, N.; Matsui, Y.; Satoi, S.; Yanagimoto, H.; Takahashi, K.; Yamamoto, T.; Yamao, J.; Takai, S.; Kwon, A.H.; Kamiyama, Y. Immunological effect of active hexose correlated compound (AHCC) in healthy volunteers: A double-blind, placebo-controlled trial. Nutr. Cancer 2008, 60, 643–651. [Google Scholar] [CrossRef]
- Matsui, Y.; Uhara, J.; Satoi, S.; Kaibori, M.; Yamada, H.; Kitade, H.; Imamura, A.; Takai, S.; Kawaguchi, Y.; Kwon, A.H.; et al. Improved prognosis of postoperative hepatocellular carcinoma patients when treated with functional foods: A prospective cohort study. J. Hepatol. 2002, 37, 78–86. [Google Scholar] [CrossRef]
- Matsushita, K.; Kuramitsu, Y.; Ohiro, Y.; Obara, M.; Kobayashi, M.; Li, Y.Q.; Hosokawa, M. Combination therapy of active hexose correlated compound plus UFT significantly reduces the metastasis of rat mammary adenocarcinoma. Anticancer Drugs 1998, 9, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, E.; Sankari, L.S.; Malathi, L.; Krupaa, J.R. Naturally occurring products in cancer therapy. J. Pharm. Bioallied. Sci. 2015, 7, S181–S183. [Google Scholar] [CrossRef] [PubMed]
- Amano, M.; Nakayama, M.; Kaibuchi, K. Rho-kinase/ROCK: A key regulator of the cytoskeleton and cell polarity. Cytoskeleton 2010, 67, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinca, R.S.; Manara, M.C.; Chiadini, V.; Picci, P.; Zucchini, C.; Scotlandi, K. Targeting ROCK2 rather than ROCK1 inhibits Ewing sarcoma malignancy. Oncol. Rep. 2017, 37, 1387–1393. [Google Scholar] [CrossRef] [Green Version]
- Zucchini, C.; Manara, M.C.; Cristalli, C.; Carrabotta, M.; Greco, S.; Pinca, R.S.; Ferrari, C.; Landuzzi, L.; Pasello, M.; Lollini, P.L.; et al. ROCK2 deprivation leads to the inhibition of tumor growth and metastatic potential in osteosarcoma cells through the modulation of YAP activity. J. Exp. Clin. Cancer Res. 2019, 38, 503. [Google Scholar] [CrossRef] [Green Version]
- Dalerba, P.; Dylla, S.J.; Park, I.K.; Liu, R.; Wang, X.; Cho, R.W.; Hoey, T.; Gurney, A.; Huang, E.H.; Simeone, D.M.; et al. Phenotypic characterization of human colorectal cancer stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 10158–10163. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Keckesova, Z.; Donaher, J.L.; Shibue, T.; Tischler, V.; Reinhardt, F.; Itzkovitz, S.; Noske, A.; Zurrer-Hardi, U.; Bell, G.; et al. Slug and Sox9 cooperatively determine the mammary stem cell state. Cell 2012, 148, 1015–1028. [Google Scholar] [CrossRef] [Green Version]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Tam, W.L.; Shibue, T.; Kaygusuz, Y.; Reinhardt, F.; Ng Eaton, E.; Weinberg, R.A. Distinct EMT programs control normal mammary stem cells and tumour-initiating cells. Nature 2015, 525, 256–260. [Google Scholar] [CrossRef] [Green Version]
- de Sousa e Melo, F.; Kurtova, A.V.; Harnoss, J.M.; Kljavin, N.; Hoeck, J.D.; Hung, J.; Anderson, J.E.; Storm, E.E.; Modrusan, Z.; Koeppen, H.; et al. A distinct role for Lgr5(+) stem cells in primary and metastatic colon cancer. Nature 2017, 543, 676–680. [Google Scholar] [CrossRef]
- Merlos-Suarez, A.; Barriga, F.M.; Jung, P.; Iglesias, M.; Cespedes, M.V.; Rossell, D.; Sevillano, M.; Hernando-Momblona, X.; da Silva-Diz, V.; Munoz, P.; et al. The intestinal stem cell signature identifies colorectal cancer stem cells and predicts disease relapse. Cell Stem. Cell 2011, 8, 511–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, N.; van Es, J.H.; Kuipers, J.; Kujala, P.; van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, A.; Oost, K.C.; Kester, L.; Morgner, J.; Bornes, L.; Bruens, L.; Spaargaren, L.; Azkanaz, M.; Schelfhorst, T.; Beerling, E.; et al. Plasticity of Lgr5-Negative Cancer Cells Drives Metastasis in Colorectal Cancer. Cell Stem. Cell 2020, 26, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Ke, J.; Sun, L.; He, Z.; Zou, Y.; He, X.; Chen, Y.; Wu, X.; Cai, Z.; Wang, L.; et al. HES1 promotes metastasis and predicts poor survival in patients with colorectal cancer. Clin. Exp. Metastasis 2015, 32, 169–179. [Google Scholar] [CrossRef]
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzyme Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Evangelisti, C.; Paganelli, F.; Giuntini, G.; Mattioli, E.; Cappellini, A.; Ramazzotti, G.; Faenza, I.; Maltarello, M.C.; Martelli, A.M.; Scotlandi, K.; et al. Lamin A and Prelamin A Counteract Migration of Osteosarcoma Cells. Cells 2020, 9, 774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Primer Sequences 5′–3′ |
|---|---|---|
| OCT4 | POU class 5 homeobox 1 | For-CCCGAAAGAGAAAGCGAACC |
| Rev-CTCTCGTTGTGCATAGTCGC | ||
| cMyc | Myc proto-oncogene | For-ATTCTCTGCTCTCCTCGACG |
| Rev-TGCGTAGTTGTGCTGATGTG | ||
| CD326 | Epithelial Cell Adhesion Molecule | For-TCTGTGAAAACTACAAGCTGGC |
| Rev-GGTTTTGCTCTTCTCCCAAGTT | ||
| CD24 | CD24 molecule | For-CATGGGCAGAGCAATGGTG |
| Rev-TGGTGGTGGCATTAGTTGGA | ||
| CD133 | Prominin 1 | For-CGACAATGTAACTCAGCGTCTT |
| Rev-CACACAGTAAGCCCAGGTAGTA | ||
| NANOG | Nanog homeobox | For-TGAGTGTGGATCCAGCTTGT |
| Rev-TCTCTGCAGAAGTGGGTTGT | ||
| LGR5 | Leucine rich repeat containing G protein-coupled receptor 5 | For-AAATGCCTTATGCTTACCAG |
| Rev-ATCTTGAGCCTGAAACATTC | ||
| ALDH | Aldehyde dehydrogenase | For-GACAATGGAGTCAATGAATGG |
| Rev-ATCAATTGGTATTGTACGGC | ||
| STAT3 | Signal transducer and activator of transcription 3 | For-CCTTTGACATGGAGTTGACC |
| Rev-TAAAAGTGCCCAGATTGCTC |
| Gene Symbol | Gene Name | Primer Sequences 5′–3′ |
|---|---|---|
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase | For-ACAGTTGCCATGTAGACC |
| Rev-TTTTTGGTTGAGCACAGG | ||
| ACTB | Actin beta | For-GGACTTCGAGCAAGAGATGG |
| Rev-AGCACTGTGTTGGCGTACAG | ||
| TFRC | Transferrin Receptor | For-CGCTGGTCAGTTCGTGATTA |
| Rev-GCATTCCCGAAATCTGTTGT |
| Gene Symbol | Gene Name | Primer Sequences 5′–3′ |
|---|---|---|
| ROCK2 | Rho associated coiled-coil containing protein kinase 2 | For-ACTCCATTTTATGCGGATTC |
| Rev-CTCCCTATCTGTTAAGAAAGC | ||
| TIMP1 | TIMP metallopeptidase inhibitor 1 | For-CACCTTATACCAGCTTATG |
| Rev-TTTCCAGCAATGAGAAACTC | ||
| CDH1 | Cadherin 1 | For-CTGGGCAGAGTGAATTTTG |
| Rev-GACTGTAATCACACCATCTG | ||
| SNAI2 | Snail family transcriptional repressor 2 | For-CAGTGATTATTTCCCCGTATC |
| Rev-CCCCAAAGATGAGGAGTATC | ||
| ITGA5 | Integrin subunit alpha 5 | For-AAGCTTGGATTCTTCAAACG |
| Rev-TCCTTTTCAGTAGAATGAGGG | ||
| LGR5 | Leucine rich repeat containing G protein-coupled receptor 5 | For-AAATGCCTTATGCTTACCAG |
| Rev-ATCTTGAGCCTGAAACATTC | ||
| NOTCH1 | Notch 1 | For-AAGATATGCAGAACAACAGG |
| Rev-TCCATATGATCCGTGATGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paganelli, F.; Chiarini, F.; Palmieri, A.; Martinelli, M.; Sena, P.; Bertacchini, J.; Roncucci, L.; Cappellini, A.; Martelli, A.M.; Bonucci, M.; et al. The Combination of AHCC and ETAS Decreases Migration of Colorectal Cancer Cells, and Reduces the Expression of LGR5 and Notch1 Genes in Cancer Stem Cells: A Novel Potential Approach for Integrative Medicine. Pharmaceuticals 2021, 14, 1325. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121325
Paganelli F, Chiarini F, Palmieri A, Martinelli M, Sena P, Bertacchini J, Roncucci L, Cappellini A, Martelli AM, Bonucci M, et al. The Combination of AHCC and ETAS Decreases Migration of Colorectal Cancer Cells, and Reduces the Expression of LGR5 and Notch1 Genes in Cancer Stem Cells: A Novel Potential Approach for Integrative Medicine. Pharmaceuticals. 2021; 14(12):1325. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121325
Chicago/Turabian StylePaganelli, Francesca, Francesca Chiarini, Annalisa Palmieri, Marcella Martinelli, Paola Sena, Jessika Bertacchini, Luca Roncucci, Alessandra Cappellini, Alberto M. Martelli, Massimo Bonucci, and et al. 2021. "The Combination of AHCC and ETAS Decreases Migration of Colorectal Cancer Cells, and Reduces the Expression of LGR5 and Notch1 Genes in Cancer Stem Cells: A Novel Potential Approach for Integrative Medicine" Pharmaceuticals 14, no. 12: 1325. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121325