
In Vitro Study of Licorice on IL-1β-Induced Chondrocytes and In Silico Approach for Osteoarthritis

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Chemical Characterization of WG by High-Performance Liquid Chromatograph (HPLC) System
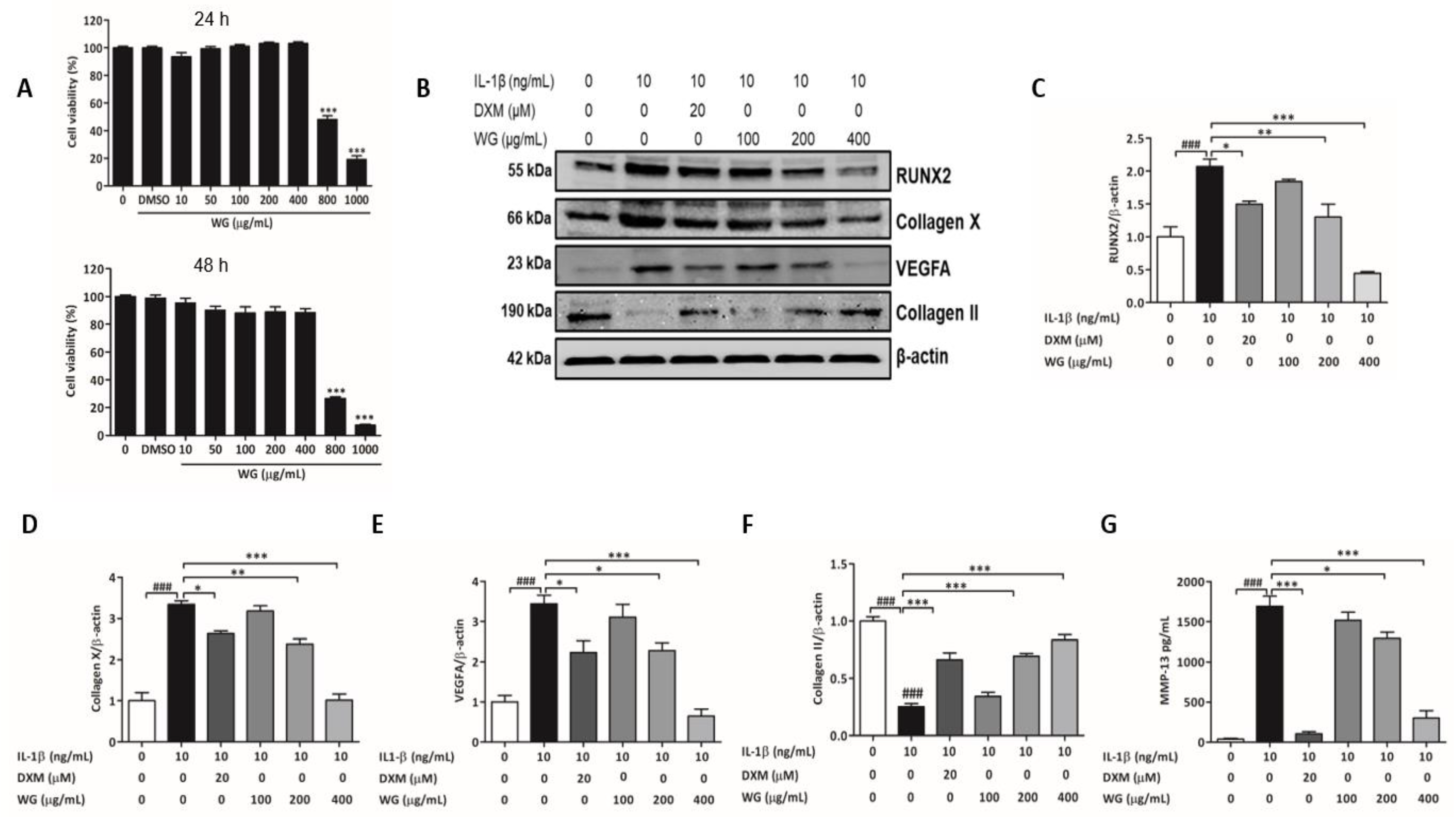
2.2. WG Ameliorated IL-1β Induced Chondrocyte Hypertrophy-like Changes
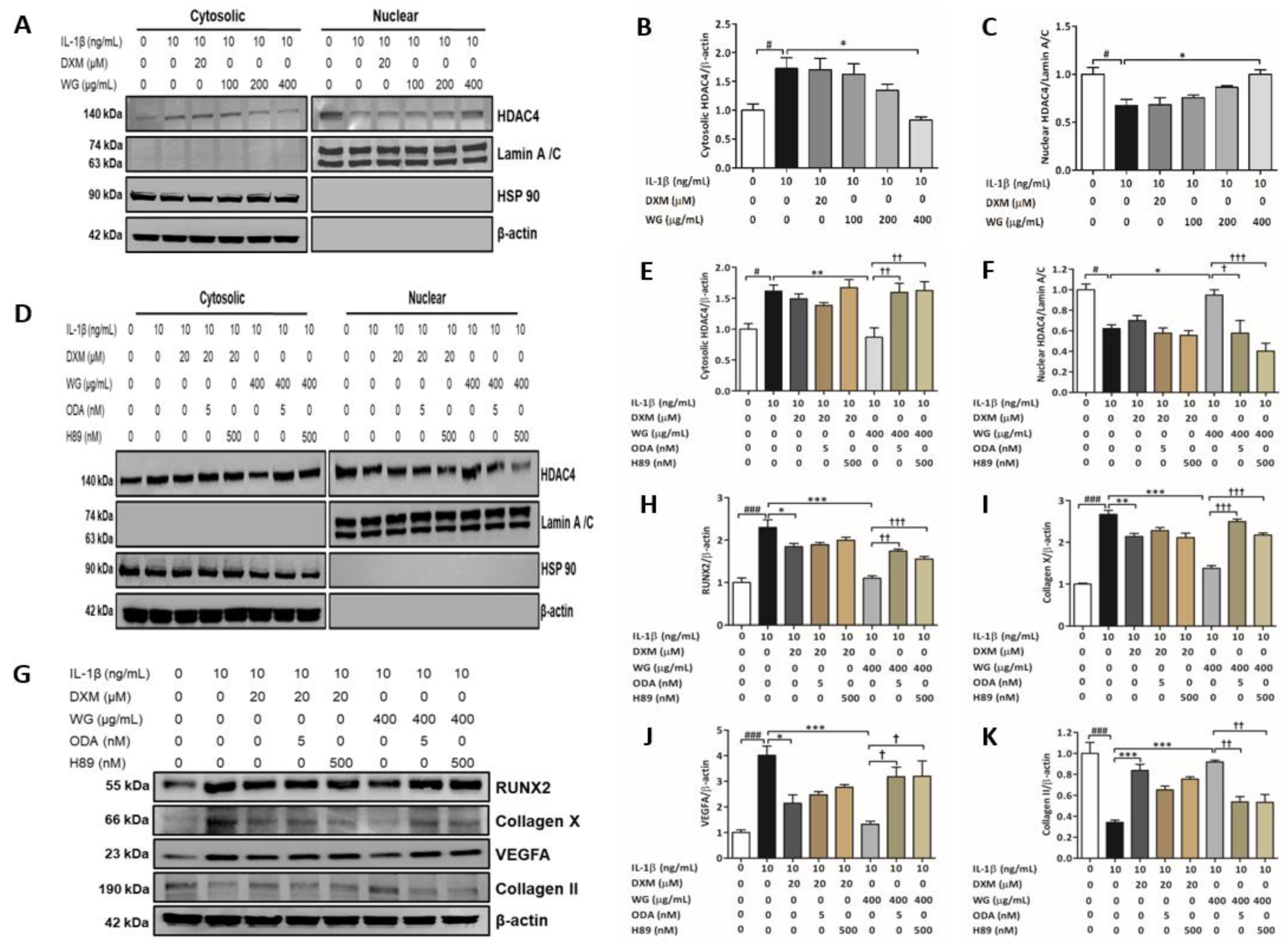
2.3. WG Stimulated HDAC4 Nuclear Translocation via PKA and PP2A
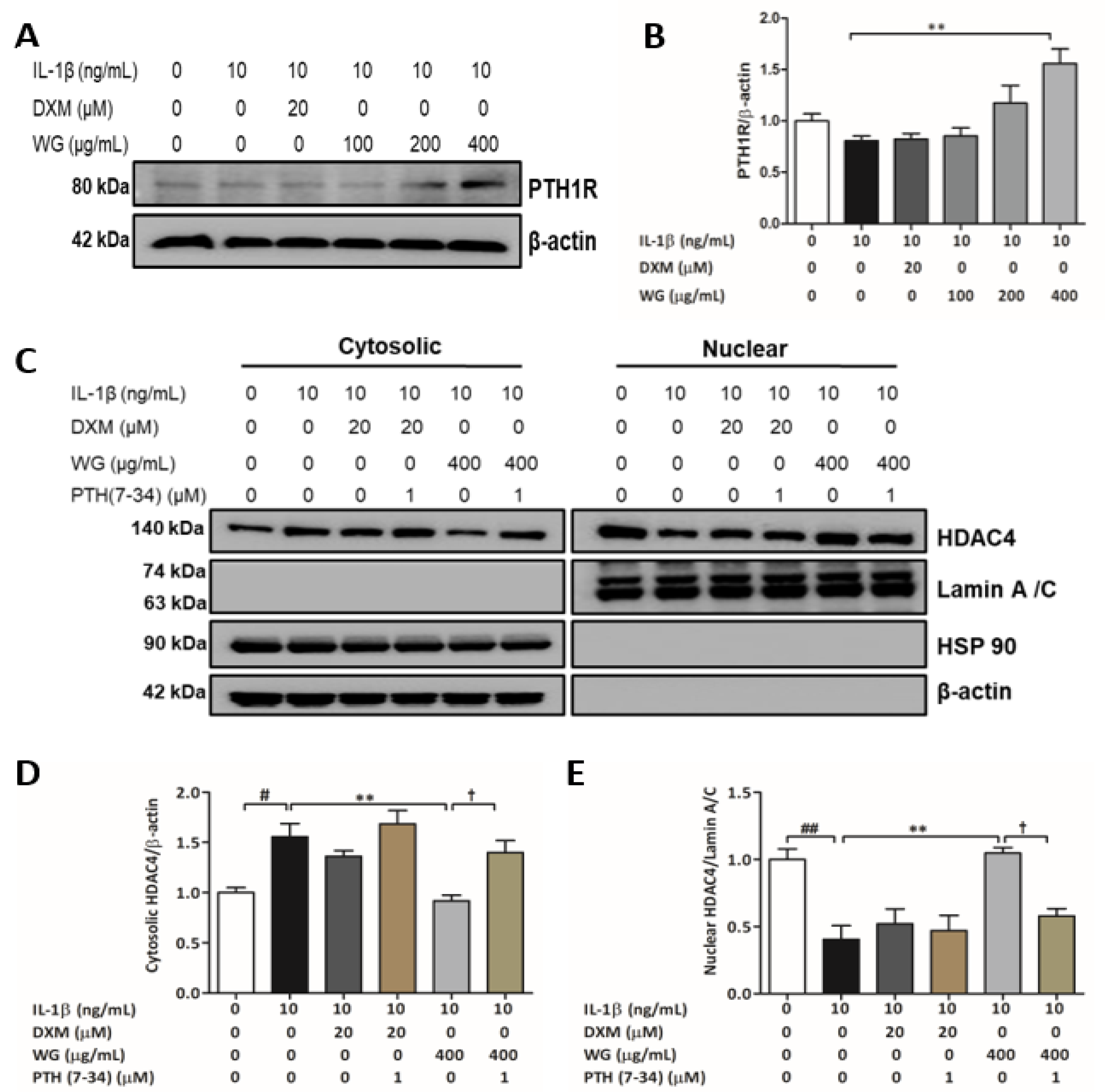
2.4. PTH1R Mediated HDAC4 Activation by WG
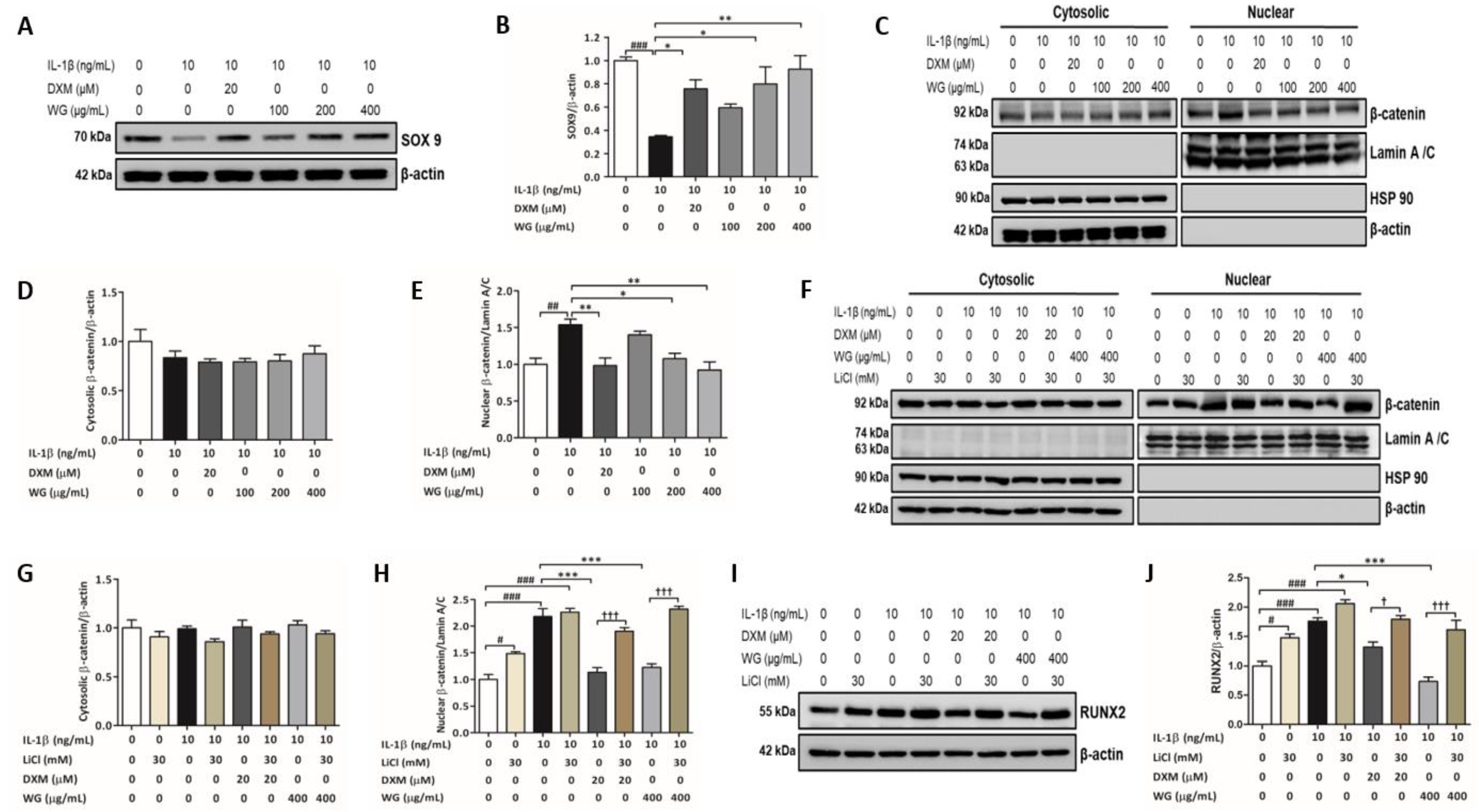
2.5. WG Increased SOX9 Expression and Decreased β-Catenin Activation
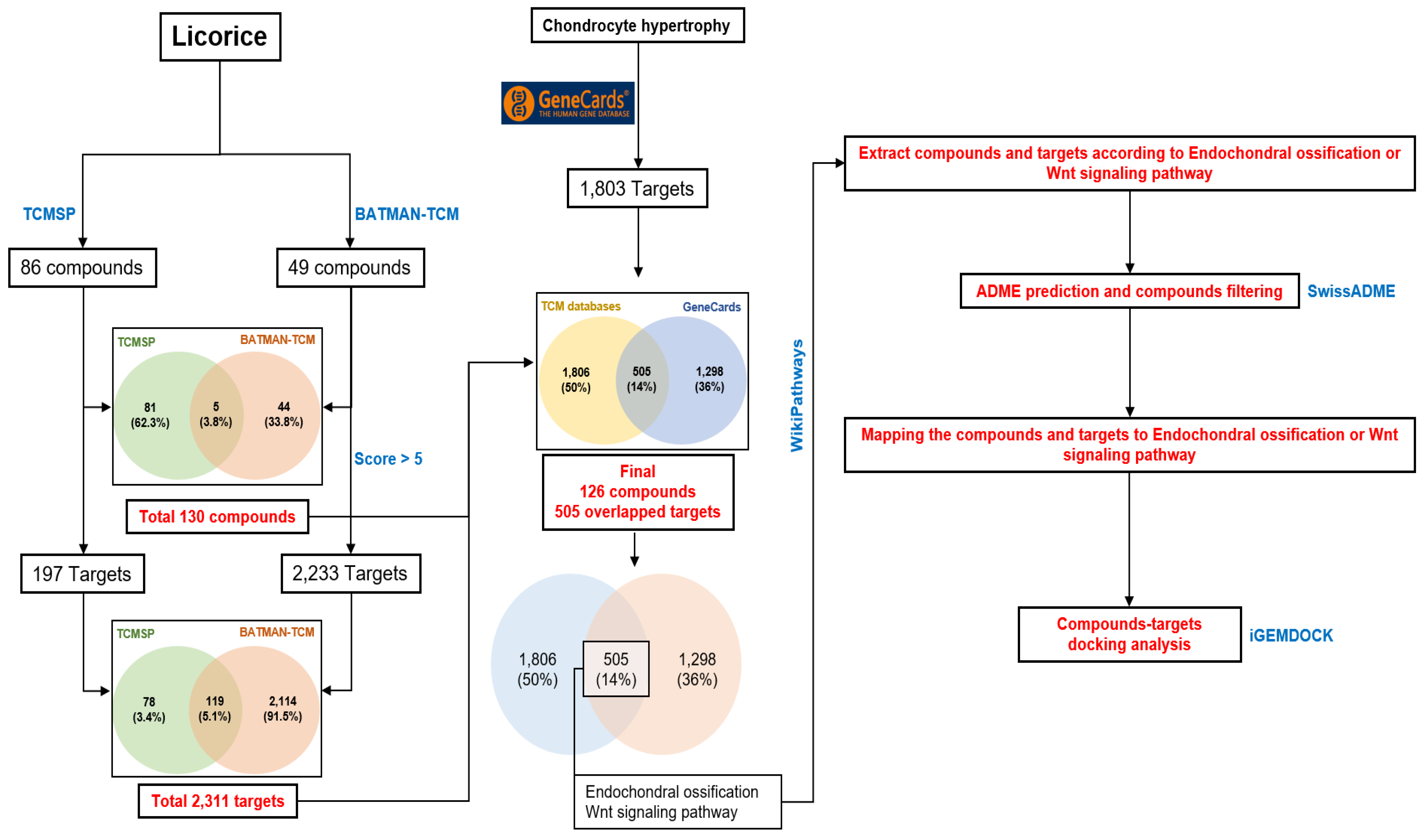
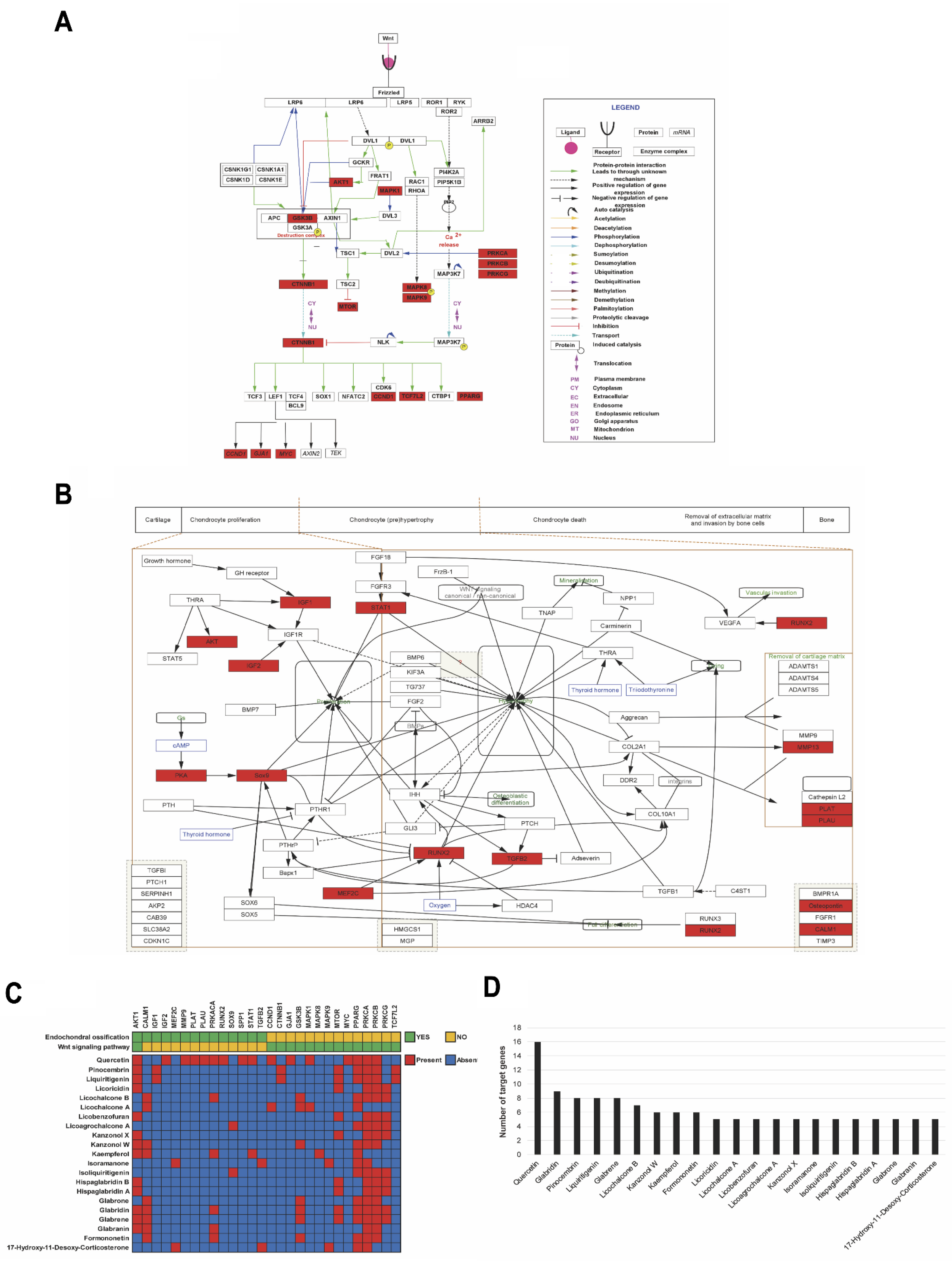
2.6. Computational Network Analysis to Identify Licorice Compounds Related to Chondrocyte Hypertrophy
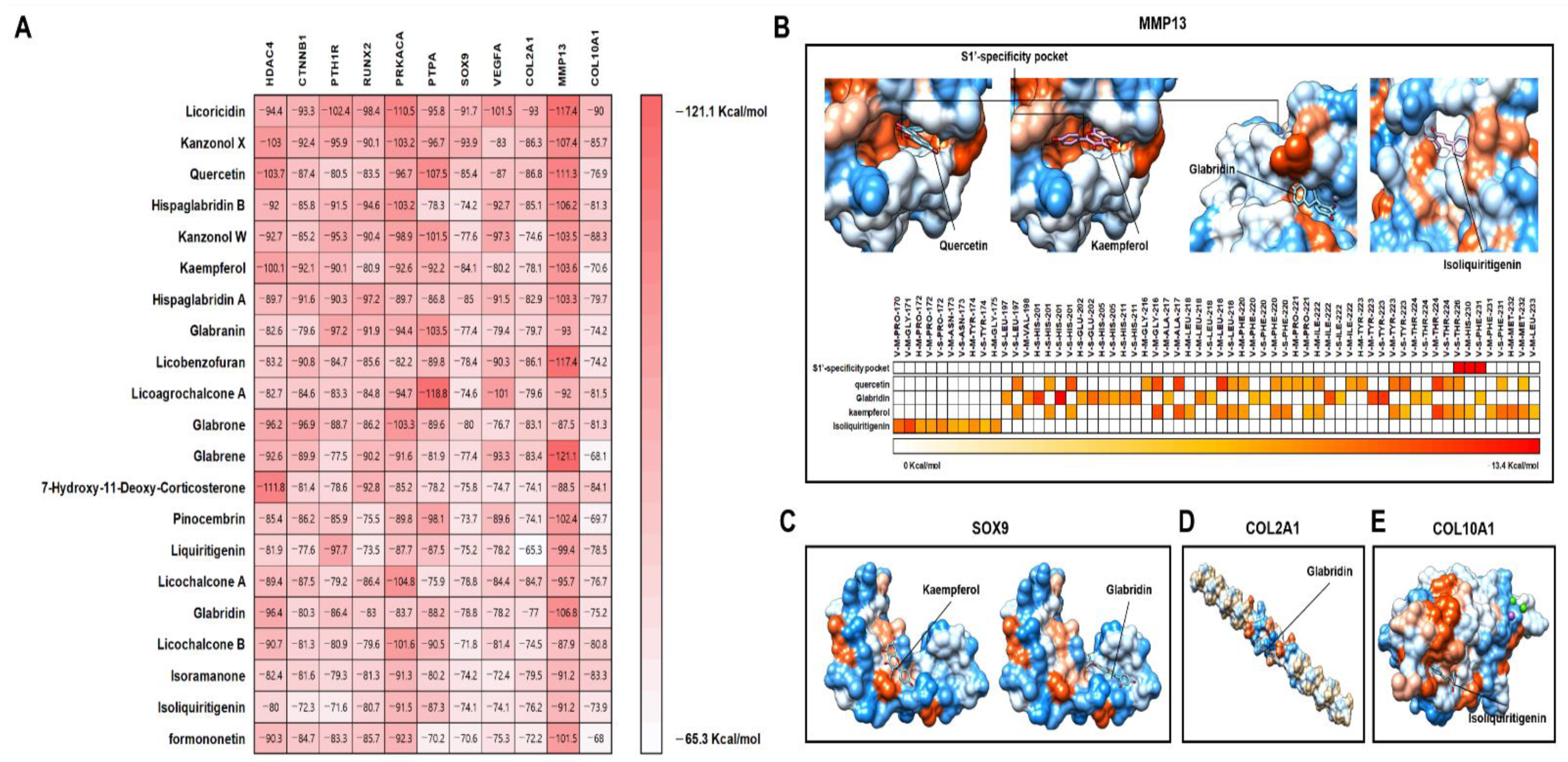
2.7. Validation of Major Active Compounds in Licorice Based on the Experimental OA Model
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of WG
4.3. HPLC and Preparation of Standard and Sample Solutions
4.4. Cell Culture and Cytotoxicity Assay
4.5. MMP-13 Assay
4.6. Western Blotting Assay
4.7. Data Collection
4.8. Gene Ontology Analysis
4.9. ADME Prediction
4.10. Protein-Compound Docking Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blagojevic, M.; Jinks, C.; Jeffery, A.; Jordan, K.P. Risk factors for onset of osteoarthritis of the knee in older adults: A systematic review and meta-analysis. Osteoarthr. Cartil. 2010, 18, 24–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breedveld, F. Osteoarthritis—The impact of a serious disease. Rheumatology 2004, 43, i4–i8. [Google Scholar] [CrossRef] [Green Version]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [Green Version]
- Belluzzi, E.; Macchi, V.; Fontanella, C.G.; Carniel, E.L.; Olivotto, E.; Filardo, G.; Sarasin, G.; Porzionato, A.; Granzotto, M.; Pozzuoli, A.; et al. Infrapatellar fat pad gene expression and protein production in patients with and without osteoarthritis. Int. J. Mol. Sci. 2020, 21, 6016. [Google Scholar] [CrossRef]
- Englund, M.; Guermazi, A.; Lohmander, S.L. The role of the meniscus in knee osteoarthritis: A cause or consequence? Radiol. Clin. N. Am. 2009, 47, 703–712. [Google Scholar] [CrossRef]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Tchetina, E.V.; Kobayashi, M.; Yasuda, T.; Meijers, T.; Pidoux, I.; Poole, A.R. Chondrocyte hypertrophy can be induced by a cryptic sequence of type ii collagen and is accompanied by the induction of mmp-13 and collagenase activity: Implications for development and arthritis. Matrix Biol. 2007, 26, 247–258. [Google Scholar] [CrossRef]
- Van der Kraan, P.M.; Van den Berg, W.B. Chondrocyte hypertrophy and osteoarthritis: Role in initiation and progression of cartilage degeneration? Osteoarthr. Cartil. 2012, 20, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Dong, S. The signaling pathways involved in chondrocyte differentiation and hypertrophic differentiation. Stem Cells Int. 2016, 2016, 2470351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastorino, G.; Cornara, L.; Soares, S.; Rodrigues, F.; Oliveira, M. Liquorice (Glycyrrhiza glabra): A phytochemical and pharmacological review. Phytother. Res. 2018, 32, 2323–2339. [Google Scholar] [CrossRef]
- Li, F.; Liu, B.; Li, T.; Wu, Q.; Xu, Z.; Gu, Y.; Li, W.; Wang, P.; Ma, T.; Lei, H. Review of constituents and biological activities of triterpene saponins from Glycyrrhizae Radix et rhizoma and its solubilization characteristics. Molecules 2020, 25, 3904. [Google Scholar] [CrossRef]
- Mamedov, N.A.; Egamberdieva, D. Phytochemical constituents and pharmacological effects of licorice: A review. In Plant and Human Health, 1st ed.; Ozturk, M., Hakeem, K.R., Eds.; Springer: New York, NY, USA, 2019; Volume 3, pp. 1–21. [Google Scholar]
- Park, C.; Lee, J.; Kim, O.; Park, C.; Kim, G.; Ahn, Y.; Cha, S.; Lee, S.; Kim, M.; Heo, C. A new Glycyrrhiza variety “wongam” through interspecific cross between Glycyrrhiza glabra and Glycyrrhiza uralensis. Korean Soc. Med. Crops Sci. 2014, 22, 169–170. [Google Scholar]
- Lee, J.-H.; Oh, M.-W.; Lee, S.-H.; Park, C.-G.; Jeong, J.-T.; Han, J.-W.; Ma, K.-H.; Chang, J.-K. ‘Wongam’, a licorice interspecific hybrid cultivar with high yield. Korean J. Breed. Sci. 2020, 52, 454–459. [Google Scholar] [CrossRef]
- Cho, M.J.; Kim, J.H.; Park, C.H.; Lee, A.Y.; Shin, Y.S.; Lee, J.H.; Park, C.G.; Cho, E.J. Comparison of the effect of three licorice varieties on cognitive improvement via an amelioration of neuroinflammation in lipopolysaccharide-induced mice. Nutr. Res. Pract. 2018, 12, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-M.; Kim, W.; Jin, J.-S.; Lee, J.-H.; Chang, J.K.; Lee, J.; An, H.-J. The comparative study of immunomodulatory effect by Glycyrrhiza new varieties and official compendia. Korea J. Herbol. 2020, 35, 11–19. [Google Scholar]
- Kang, Y.-M.; Kim, W.; Jin, J.-S.; Lee, J.-H.; Chang, J.K.; Lee, J.; An, H.-J. The comparative study of anti-allergic effect by glycyrrhiza new varieties and official compendia. Korea J. Herbol. 2020, 35, 13–21. [Google Scholar]
- Kang, S.-H.; Song, Y.-J.; Jeon, Y.-D.; Kim, D.-K.; Park, J.-H.; Soh, J.-R.; Lee, J.-H.; Kitalong, C.; Kim, W.; An, H.-J.; et al. Comparative study of anti-inflammatory effect on dss-induced ulcerative colitis between novel Glycyrrhiza variety and official compendia. Appl. Sci. 2021, 11, 1545. [Google Scholar] [CrossRef]
- Kim, M.; Kang, M.; Lee, J.; Leem, K.-H.; An, H.-J.; Jin, J.-S.; Lee, J.-H.; Chang, J.; Seong, S.; Kim, W. Antioxidant activity and cytotoxicity against human cancer cells of Glycyrrhiza new varieties: A comparison with Glycyrrhiza official compendia. Korea J. Herbol. 2021, 36, 15–24. [Google Scholar]
- Huebner, K.D.; Shrive, N.G.; Frank, C.B. Dexamethasone inhibits inflammation and cartilage damage in a new model of post-traumatic osteoarthritis. J. Orthop. Res. 2014, 32, 566–572. [Google Scholar] [CrossRef]
- Black, R.; Grodzinsky, A.J. Dexamethasone: Chondroprotective corticosteroid or catabolic killer? Eur. Cell Mater. 2019, 38, 246–263. [Google Scholar] [CrossRef]
- Mackie, E.J.; Ahmed, Y.A.; Tatarczuch, L.; Chen, K.S.; Mirams, M. Endochondral ossification: How cartilage is converted into bone in the developing skeleton. Int. J. Biochem. Cell Biol. 2008, 40, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Mackie, E.J.; Tatarczuch, L.; Mirams, M. The skeleton: A multi-functional complex organ: The growth plate chondrocyte and endochondral ossification. J. Endocrinol. 2011, 211, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, Z.; Guo, L.; Wei, X.; Zhang, Y.; Wang, X.; Wei, L. The role of histone deacetylase 4 during chondrocyte hypertrophy and endochondral bone development. Bone Jt. Res. 2020, 9, 82–89. [Google Scholar] [CrossRef]
- Kozhemyakina, E.; Cohen, T.; Yao, T.P.; Lassar, A.B. Parathyroid hormone-related peptide represses chondrocyte hypertrophy through a protein phosphatase 2a/histone deacetylase 4/mef2 pathway. Mol. Cell Biol. 2009, 29, 5751–5762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Chung, U.I.; Yang, D.; Karsenty, G.; Bringhurst, F.R.; Kronenberg, H.M. Pth/pthrp receptor delays chondrocyte hypertrophy via both runx2-dependent and -independent pathways. Dev. Biol. 2006, 292, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Nishimori, S.; Lai, F.; Shiraishi, M.; Kobayashi, T.; Kozhemyakina, E.; Yao, T.P.; Lassar, A.B.; Kronenberg, H.M. PTHrP targets HDAC4 and HDAC5 to repress chondrocyte hypertrophy. JCI Insight 2019, 4, e97903. [Google Scholar] [CrossRef]
- Zhou, G.; Zheng, Q.; Engin, F.; Munivez, E.; Chen, Y.; Sebald, E.; Krakow, D.; Lee, B. Dominance of sox9 function over runx2 during skeletogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19004–19009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topol, L.; Chen, W.; Song, H.; Day, T.F.; Yang, Y. Sox9 inhibits wnt signaling by promoting β-catenin phosphorylation in the nucleus. J. Biol. Chem. 2009, 284, 3323–3333. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Zhang, Y.; Tang, L.; Ji, Y.; Yan, C.; Zhang, X. Protective effects of quercetin against inflammation and oxidative stress in a rabbit model of knee osteoarthritis. Drug Dev. Res. 2019, 80, 360–367. [Google Scholar] [CrossRef]
- Dai, J.; Zhang, Y.; Chen, D.; Chen, D.; Li, X.; Wang, J.; Sun, Y. Glabridin inhibits osteoarthritis development by protecting chondrocytes against oxidative stress, apoptosis and promoting mtor mediated autophagy. Life Sci. 2021, 268, 118992. [Google Scholar] [CrossRef]
- Ji, B.; Zhang, Z.; Guo, W.; Ma, H.; Xu, B.; Mu, W.; Amat, A.; Cao, L. Isoliquiritigenin blunts osteoarthritis by inhibition of bone resorption and angiogenesis in subchondral bone. Sci. Rep. 2018, 8, 1721. [Google Scholar] [CrossRef]
- Estakhri, F.; Panjehshahin, M.R.; Tanideh, N.; Gheisari, R.; Mahmoodzadeh, A.; Azarpira, N.; Gholijani, N. The effect of kaempferol and apigenin on allogenic synovial membrane-derived stem cells therapy in knee osteoarthritic male rats. Knee 2020, 27, 817–832. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; March, L.; Chew, M. Osteoarthritis in 2020 and beyond: A lancet commission. Lancet 2020, 396, 1711–1712. [Google Scholar] [CrossRef]
- Vinatier, C.; Merceron, C.; Guicheux, J. Osteoarthritis: From pathogenic mechanisms and recent clinical developments to novel prospective therapeutic options. Drug Discov. Today 2016, 21, 1932–1937. [Google Scholar] [CrossRef] [PubMed]
- Ding, C. Do NSAIDs affect the progression of osteoarthritis? Inflammation 2002, 26, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liang, J.; Zhang, J.; Wang, Y.; Chai, X. Natural chalcones in chinese materia medica: Licorice. Evid. Based Complement. Alternat. Med. 2020, 2020, 3821248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.H.; Xu, J.J.; Zhu, D.C.; Li, J.F.; Zhang, C.X.; Lin, N.; Gao, W.Y. Glycyrrhizin inhibits osteoarthritis development through suppressing the pi3k/akt/nf-kappab signaling pathway in vivo and in vitro. Food Funct. 2020, 11, 2126–2136. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Kim, D.J.; Shen, J.; Zou, Z.; O’Keefe, R.J. Runx2 plays a central role in osteoarthritis development. J. Orthop. Translat. 2020, 23, 132–139. [Google Scholar] [CrossRef]
- Liao, L.; Zhang, S.; Gu, J.; Takarada, T.; Yoneda, Y.; Huang, J.; Zhao, L.; Oh, C.D.; Li, J.; Wang, B.; et al. Deletion of runx2 in articular chondrocytes decelerates the progression of dmm-induced osteoarthritis in adult mice. Sci. Rep. 2017, 7, 2371. [Google Scholar] [CrossRef]
- Tang, Q.; Zheng, G.; Feng, Z.; Tong, M.; Xu, J.; Hu, Z.; Shang, P.; Chen, Y.; Wang, C.; Lou, Y.; et al. Wogonoside inhibits il-1beta induced catabolism and hypertrophy in mouse chondrocyte and ameliorates murine osteoarthritis. Oncotarget 2017, 8, 61440–61456. [Google Scholar] [CrossRef] [Green Version]
- Nurminskaya, M.; Linsenmayer, T.F. Identification and characterization of up-regulated genes during chondrocyte hypertrophy. Dev. Dyn. 1996, 206, 260–271. [Google Scholar] [CrossRef]
- Wang, M.; Sampson, E.R.; Jin, H.; Li, J.; Ke, Q.H.; Im, H.J.; Chen, D. MMP13 is a critical target gene during the progression of osteoarthritis. Arthritis Res. Ther. 2013, 15, R5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rim, Y.A.; Nam, Y.; Ju, J.H. The role of chondrocyte hypertrophy and senescence in osteoarthritis initiation and progression. Int. J. Mol. Sci. 2020, 21, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, R.B.; Matsuda, K.; Oh, J.; Barbosa, A.C.; Yang, X.; Meadows, E.; McAnally, J.; Pomajzl, C.; Shelton, J.M.; Richardson, J.A.; et al. Histone deacetylase 4 controls chondrocyte hypertrophy during skeletogenesis. Cell 2004, 119, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Guan, Y.; Tian, S.; Wang, Y.; Sun, K.; Chen, Q. Mechanical and il-1beta responsive mir-365 contributes to osteoarthritis development by targeting histone deacetylase 4. Int. J. Mol. Sci. 2016, 17, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, Q.; Gan, Y.H.; Shi, R.R.; Meng, J.H. Effects of hdac4 on il-1beta-induced matrix metalloproteinase expression regulated partially through the wnt3a/beta-catenin pathway. Chin. Med. J. 2021, 134, 963–970. [Google Scholar] [CrossRef]
- Paroni, G.; Cernotta, N.; Dello Russo, C.; Gallinari, P.; Pallaoro, M.; Foti, C.; Talamo, F.; Orsatti, L.; Steinkuhler, C.; Brancolini, C. Pp2a regulates hdac4 nuclear import. Mol. Biol. Cell 2008, 19, 655–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stove, J.; Schoniger, R.; Huch, K.; Brenner, R.; Gunther, K.P.; Puhl, W.; Scharf, H.P. Effects of dexamethasone on proteoglycan content and gene expression of il-1beta-stimulated osteoarthrotic chondrocytes in vitro. Acta Orthop. Scand. 2002, 73, 562–567. [Google Scholar] [CrossRef]
- Grodzinsky, A.J.; Wang, Y.; Kakar, S.; Vrahas, M.S.; Evans, C.H. Intra-articular dexamethasone to inhibit the development of post-traumatic osteoarthritis. J. Orthop. Res. 2017, 35, 406–411. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.C.; Evans, C.H.; Grodzinsky, A.J. Effects of short-term glucocorticoid treatment on changes in cartilage matrix degradation and chondrocyte gene expression induced by mechanical injury and inflammatory cytokines. Arthritis Res. Ther. 2011, 13, R142. [Google Scholar] [CrossRef] [Green Version]
- Almalki, S.G.; Agrawal, D.K. Key transcription factors in the differentiation of mesenchymal stem cells. Differentiation 2016, 92, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Weissenberger, M.; Weissenberger, M.H.; Gilbert, F.; Groll, J.; Evans, C.H.; Steinert, A.F. Reduced hypertrophy in vitro after chondrogenic differentiation of adult human mesenchymal stem cells following adenoviral SOX9 gene delivery. BMC Musculoskelet. Disord. 2020, 21, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Huang, H.; Pan, X.; Li, S.; Xie, Z.; Ma, Y.; Hu, B.; Wang, J.; Chen, Z.; Shi, P. EGR1 promotes the cartilage degeneration and hypertrophy by activating the Krüppel-like factor 5 and beta-catenin signaling. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2490–2503. [Google Scholar] [CrossRef]
- Chen, D.H.; Zheng, G.; Zhong, X.Y.; Lin, Z.H.; Yang, S.W.; Liu, H.X.; Shang, P. Oroxylin A attenuates osteoarthritis progression by dual inhibition of cell inflammation and hypertrophy. Food Funct. 2021, 12, 328–339. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.F.; Soung, D.Y.; Schwarz, E.M.; O’Keefe, R.J.; Drissi, H. Wnt induction of chondrocyte hypertrophy through the Runx2 transcription factor. J. Cell Physiol. 2006, 208, 77–86. [Google Scholar] [CrossRef]
- Golovchenko, S.; Hattori, T.; Hartmann, C.; Gebhardt, M.; Gebhard, S.; Hess, A.; Pausch, F.; Schlund, B.; Von der Mark, K. Deletion of beta catenin in hypertrophic growth plate chondrocytes impairs trabecular bone formation. Bone 2013, 55, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Kamekura, S.; Hoshi, K.; Shimoaka, T.; Chung, U.; Chikuda, H.; Yamada, T.; Uchida, M.; Ogata, N.; Seichi, A.; Nakamura, K.; et al. Osteoarthritis development in novel experimental mouse models induced by knee joint instability. Osteoarthr. Cartil. 2005, 13, 632–641. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Qi, X.; Duan, J.; Zheng, Y.; Xu, H.; Chen, X.; Chen, L.; Xiang, X. Characterization of pathological and biochemical changes in rat destabilization of medial meniscus models of osteoarthritis. Int. J. Clin. Exp. Med. 2018, 11, 9572–9577. [Google Scholar]
- Kamekura, S.; Kawasaki, Y.; Hoshi, K.; Shimoaka, T.; Chikuda, H.; Maruyama, Z.; Komori, T.; Sato, S.; Takeda, S.; Karsenty, G.; et al. Contribution of runt-related transcription factor 2 to the pathogenesis of osteoarthritis in mice after induction of knee joint instability. Arthritis Rheum. 2006, 54, 2462–2470. [Google Scholar] [CrossRef]
- Vijayababu, M.R.; Kanagaraj, P.; Arunkumar, A.; Ilangovan, R.; Dharmarajan, A.; Arunakaran, J. Quercetin induces p53-independent apoptosis in human prostate cancer cells by modulating Bcl-2-related proteins: A possible mediation by IGFBP-3. Oncol. Res. 2006, 16, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, N.; Saito, K.; Maeda, A.; Kitagawa, Y.; Kiso, Y.; Watanabe, K.; Tomonaga, A.; Nagaoka, I.; Yamaguchi, H. Effect of a dietary supplement containing glucosamine hydrochloride, chondroitin sulfate and quercetin glycosides on symptomatic knee osteoarthritis: A randomized, double-blind, placebo-controlled study. J. Sci. Food Agric. 2012, 92, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Lakroun, Z.; Kebieche, M.; Lahouel, A.; Zama, D.; Desor, F.; Soulimani, R. Oxidative stress and brain mitochondria swelling induced by endosulfan and protective role of quercetin in rat. Environ. Sci. Pollut. Res. Int. 2015, 22, 7776–7781. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Sun, C.; Zhou, B.; Xing, H.; Ma, D.; Chen, G.; Weng, D. Low concentration of quercetin antagonizes the cytotoxic effects of anti-neoplastic drugs in ovarian cancer. PLoS ONE 2014, 9, e100314. [Google Scholar]
- Simmler, C.; Pauli, G.F.; Chen, S.N. Phytochemistry and biological properties of glabridin. Fitoterapia 2013, 90, 160–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, F.; Du, Q.; Peng, C.; Wang, N.; Tang, H.; Xie, X.; Shen, J.; Chen, J. A review: The pharmacology of isoliquiritigenin. Phytother. Res. 2015, 29, 969–977. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Tavakol, S.; Ahmadi, Z.; Roomiani, S.; Mohammadinejad, R.; Samarghandian, S. Therapeutic effects of kaempferol affecting autophagy and endoplasmic reticulum stress. Phytother. Res. 2020, 34, 911–923. [Google Scholar] [CrossRef]
- Johnson, A.R.; Pavlovsky, A.G.; Ortwine, D.F.; Prior, F.; Man, C.F.; Bornemeier, D.A.; Banotai, C.A.; Mueller, W.T.; McConnell, P.; Yan, C.; et al. Discovery and characterization of a novel inhibitor of matrix metalloprotease-13 that reduces cartilage damage in vivo without joint fibroplasia side effects. J. Biol. Chem. 2007, 282, 27781–27791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.; Peng, L.; Kang, A.; Xie, T.; Xu, J.; Shen, C.; Ji, J.; Di, L.; Wu, H.; Shan, J. Influence of Jiegeng on pharmacokinetic properties of flavonoids and saponins in Gancao. Molecules 2017, 22, 1587. [Google Scholar] [CrossRef] [Green Version]
- Braune, A.; Gutschow, M.; Blaut, M. An NADH-dependent reductase from eubacterium ramulus catalyzes the stereospecific heteroring cleavage of flavanones and flavanonols. Appl. Environ. Microbiol. 2019, 85, e01233-19. [Google Scholar] [CrossRef] [Green Version]
- Riva, A.; Kolimar, D.; Spittler, A.; Wisgrill, L.; Herbold, C.W.; Abranko, L.; Berry, D. Conversion of rutin, a prevalent dietary flavonol, by the human gut microbiota. Front. Microbiol. 2020, 11, 585428. [Google Scholar] [CrossRef]
- Shimojo, Y.; Ozawa, Y.; Toda, T.; Igami, K.; Shimizu, T. Probiotic Lactobacillus paracasei a221 improves the functionality and bioavailability of kaempferol-glucoside in kale by its glucosidase activity. Sci. Rep. 2018, 8, 9239. [Google Scholar] [CrossRef]
- Tulukcu, E.; Cebi, N.; Sagdic, O. Chemical fingerprinting of seeds of some salvia species in turkey by using GC-MS and FTIR. Foods 2019, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.F.; Cheng, X.L.; Lin, Q.H.; Li, S.S.; Jia, Z.; Han, T.; Lin, R.C.; Wang, D.; Wei, F.; Li, X.R. Identification of mountain-cultivated ginseng and cultivated ginseng using UPLC/oa-TOF MSE with a multivariate statistical sample-profiling strategy. J. Ginseng Res. 2016, 40, 344–350. [Google Scholar] [CrossRef]
- Papaioanou, M.; Chronopoulou, E.G.; Ciobotari, G.; Efrose, R.C.; Sfichi-Duke, L.; Chatzikonstantinou, M.; Pappa, E.; Ganopoulos, I.; Madesis, P.; Nianiou-Obeidat, I.; et al. Evaluation of the nutraceutical and cosmeceutical potential of two cultivars of Rubus fruticosus L. under different cultivation conditions. Curr. Pharm. Biotechnol. 2017, 18, 890–899. [Google Scholar] [CrossRef]
- Fuji, Y.; Uchida, A.; Fukahori, K.; Chino, M.; Ohtsuki, T.; Matsufuji, H. Chemical characterization and biological activity in young sesame leaves (Sesamum indicum L.) and changes in iridoid and polyphenol content at different growth stages. PLoS ONE 2018, 13, e0194449. [Google Scholar] [CrossRef]
- Ayeka, P.A.; Bian, Y.; Githaiga, P.M.; Zhao, Y. The immunomodulatory activities of licorice polysaccharides (Glycyrrhiza uralensis Fisch.) in ct 26 tumor-bearing mice. BMC Complement. Altern. Med. 2017, 17, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Zeng, G.; Sun, B.; Cai, X.; Bi, L.; Tang, G.; Yang, Y. A polysaccharide from Glycyrrhiza inflata licorice inhibits proliferation of human oral cancer cells by inducing apoptosis via mitochondrial pathway. Tumour Biol. 2015, 36, 4825–4831. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.; Wang, X.; Ma, X.; Jin, Y.; Fan, W.; Laba, C.; Wujin, C.; Wang, Y.; Liang, J. Preparation of complex microcapsules of soluble polysaccharide from Glycyrrhiza uralensis and its application in wound repair and scar inhibition. Int. J. Biol. Macromol. 2020, 156, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Bello, A.; Arai, Y.; Ahn, J.; Kim, D.; Cha, K.Y.; Baek, I.; Park, H.; Lee, S.H. Functional duality of chondrocyte hypertrophy and biomedical application trends in osteoarthritis. Pharmaceutics 2021, 13, 1139. [Google Scholar] [CrossRef] [PubMed]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Guo, F.; Wang, Y.; Li, C.; Zhang, X.; Li, H.; Diao, L.; Gu, J.; Wang, W.; Li, D.; et al. BATMAN-TCM: A bioinformatics analysis tool for molecular mechanism of traditional chinese medicine. Sci. Rep. 2016, 6, 21146. [Google Scholar] [CrossRef]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. Uniprot: The universal protein knowledgebase. Nucleic Acids Res. 2004, 32 (Suppl. 1), D115–D119. [Google Scholar] [CrossRef] [PubMed]
- Safran, M.; Dalah, I.; Alexander, J.; Rosen, N.; Iny Stein, T.; Shmoish, M.; Nativ, N.; Bahir, I.; Doniger, T.; Krug, H.; et al. Genecards version 3: The human gene integrator. Database 2010, 2010, baq020. [Google Scholar] [CrossRef] [PubMed]
- Slenter, D.N.; Kutmon, M.; Hanspers, K.; Riutta, A.; Windsor, J.; Nunes, N.; Melius, J.; Cirillo, E.; Coort, S.L.; Digles, D.; et al. Wikipathways: A multifaceted pathway database bridging metabolomics to other omics research. Nucleic Acids Res. 2018, 46, D661–D667. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. Cluego: A cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. Swissadme: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliman, K.; Grimm, F.; Wurm, C.A.; Egner, A. Predicting the membrane permeability of organic fluorescent probes by the deep neural network based lipophilicity descriptor DeepFl-LogP. Sci. Rep. 2021, 11, 6991. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. Pubchem substance and compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The protein data bank. Acta Crystallogr. D Biol. Crystallogr. 2002, 58, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, K.C.; Chen, Y.F.; Lin, S.R.; Yang, J.M. iGEMDOCK: A graphical environment of enhancing gemdock using pharmacological interactions and post-screening analysis. BMC Bioinform. 2011, 12, S33. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound ** | MW * | Consensus Log P | GI Absorption * | Bioavailability Score |
|---|---|---|---|---|
| Quercetin | 302.24 | 1.23 | High | 0.55 |
| Pinocembrin | 256.25 | 2.26 | High | 0.55 |
| Liquiritigenin | 256.25 | 2.07 | High | 0.55 |
| Licoricidin | 424.53 | 5.05 | High | 0.55 |
| Licochalcone B | 286.28 | 2.14 | High | 0.55 |
| licochalcone A | 338.4 | 3.98 | High | 0.55 |
| Licobenzofuran | 354.4 | 4.15 | High | 0.55 |
| Licoagrochalcone A | 324.37 | 3.86 | High | 0.55 |
| L-Aspartic Acid | 133.1 | −2.76 | Low | 0.56 |
| Kanzonol X | 394.5 | 5.06 | High | 0.55 |
| kanzonol W | 336.34 | 3.36 | High | 0.55 |
| kaempferol | 286.24 | 1.58 | High | 0.55 |
| Isoramanone | 348.48 | 2.25 | High | 0.55 |
| Isoliquiritigenin | 256.25 | 2.37 | High | 0.55 |
| Hispaglabridin B | 390.47 | 4.69 | High | 0.55 |
| Hispaglabridin A | 392.49 | 4.93 | High | 0.55 |
| Glabrone | 336.34 | 3.13 | High | 0.55 |
| Glabridin | 324.37 | 3.45 | High | 0.55 |
| Glabrene | 322.35 | 3.38 | High | 0.55 |
| Glabranin | 324.37 | 3.65 | High | 0.55 |
| Formononetin | 268.26 | 2.66 | High | 0.55 |
| 5-Hydroxyisophthalic Acid | 182.13 | 0.54 | High | 0.56 |
| 17-Hydroxy-11-Deoxy-Corticosterone | 360.49 | 3.02 | High | 0.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.; Park, Y.; Lee, J.; An, H.-J.; Jin, J.-S.; Lee, J.-H.; Chang, J.; Kim, D.-K.; Goo, B.; Park, Y.C.; et al. In Vitro Study of Licorice on IL-1β-Induced Chondrocytes and In Silico Approach for Osteoarthritis. Pharmaceuticals 2021, 14, 1337. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121337
Ali A, Park Y, Lee J, An H-J, Jin J-S, Lee J-H, Chang J, Kim D-K, Goo B, Park YC, et al. In Vitro Study of Licorice on IL-1β-Induced Chondrocytes and In Silico Approach for Osteoarthritis. Pharmaceuticals. 2021; 14(12):1337. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121337
Chicago/Turabian StyleAli, Akhtar, YoungJoon Park, Jeonghoon Lee, Hyo-Jin An, Jong-Sik Jin, Jong-Hyun Lee, Jaeki Chang, Dong-Keun Kim, Bonhyuk Goo, Yeon Cheol Park, and et al. 2021. "In Vitro Study of Licorice on IL-1β-Induced Chondrocytes and In Silico Approach for Osteoarthritis" Pharmaceuticals 14, no. 12: 1337. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121337