
Acute MDPV Binge Paradigm on Mice Emotional Behavior and Glial Signature

 ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
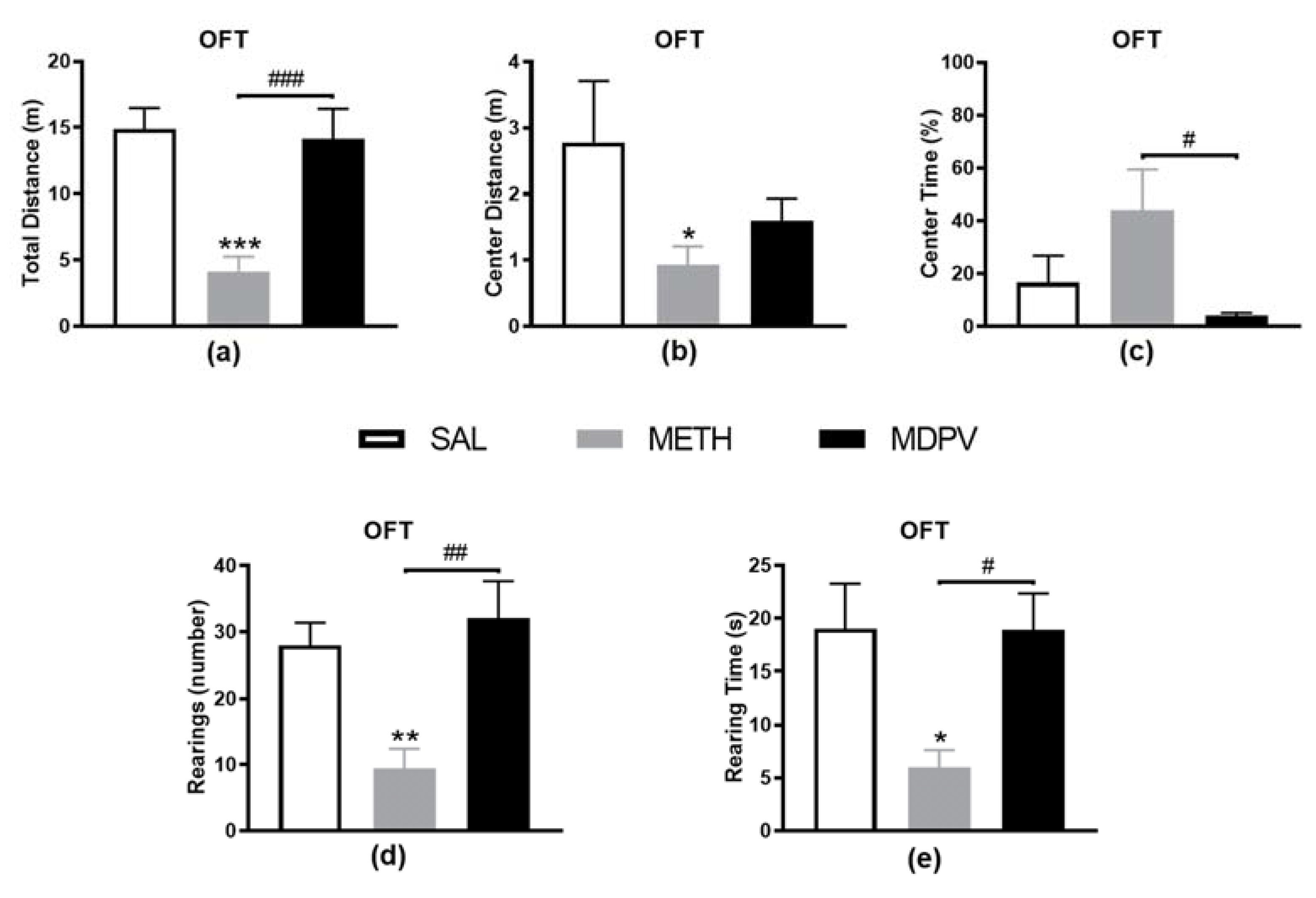
2.1. MPDV and METH on Locomotor and Exploratory Activity
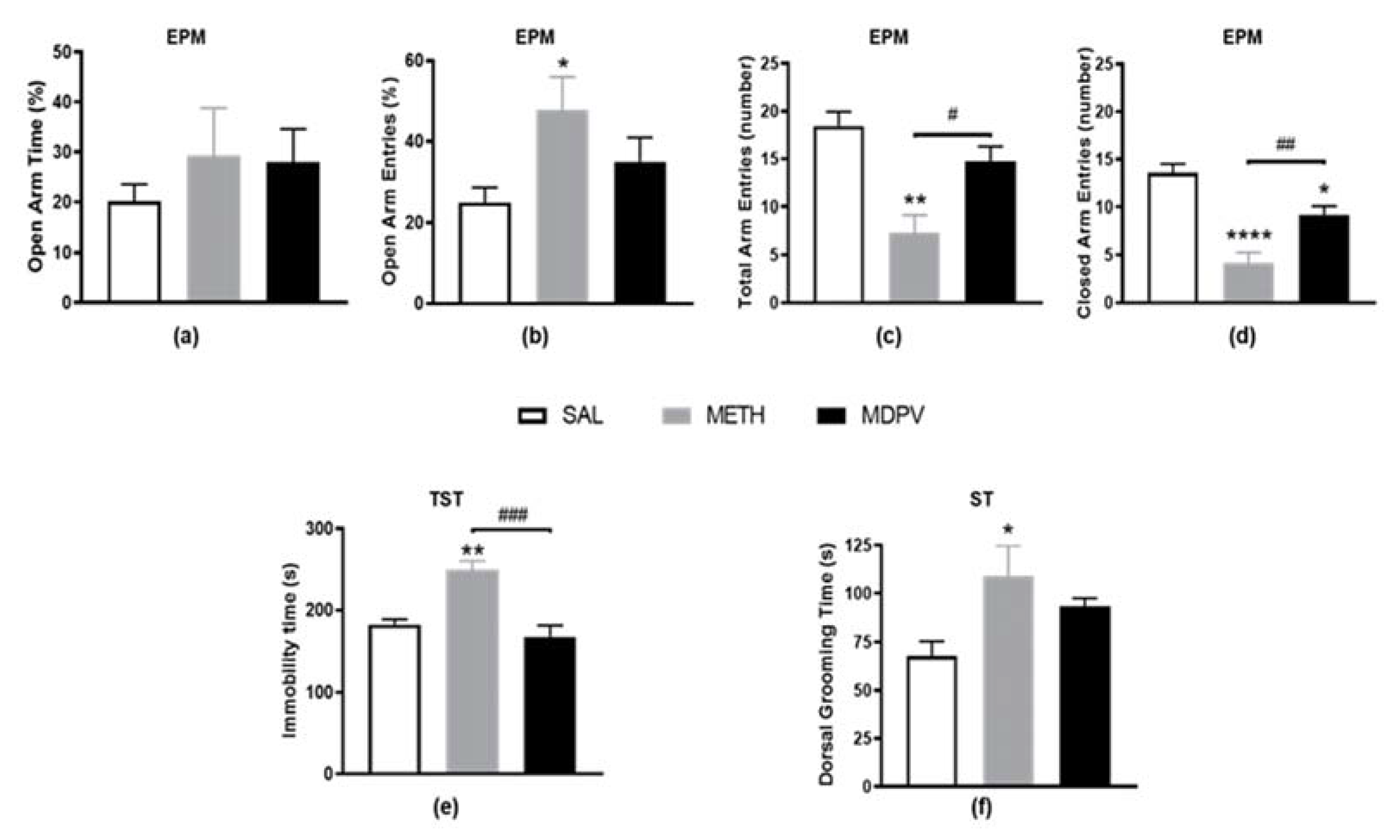
2.2. MPDV and METH on Emotional Activity
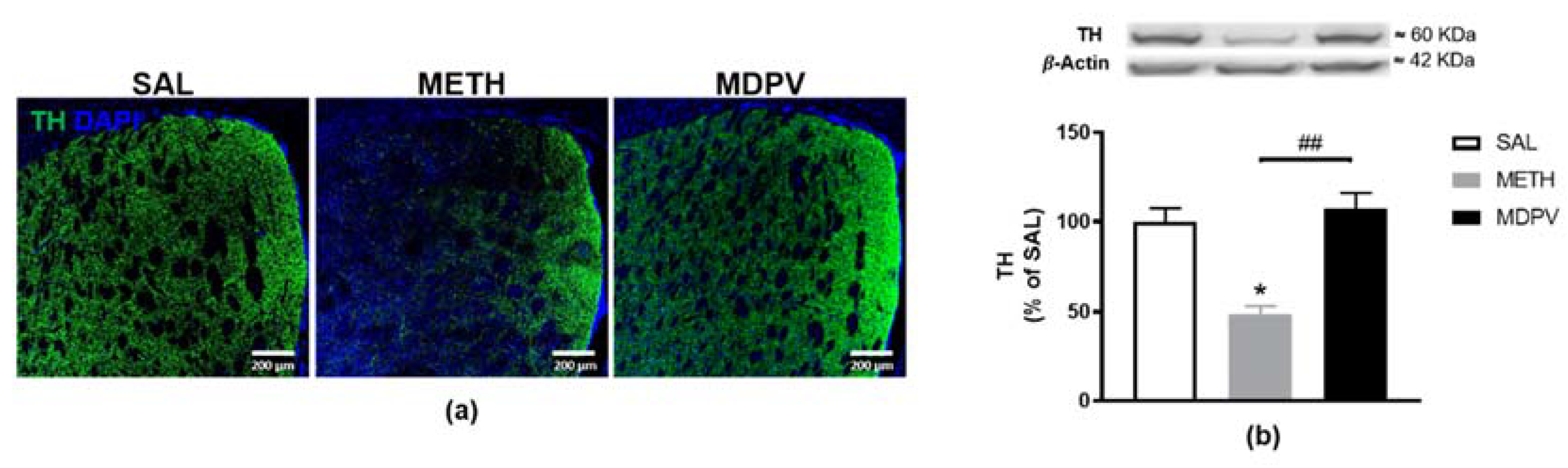
2.3. MPDV and METH on Striatal Dopaminergic Terminals
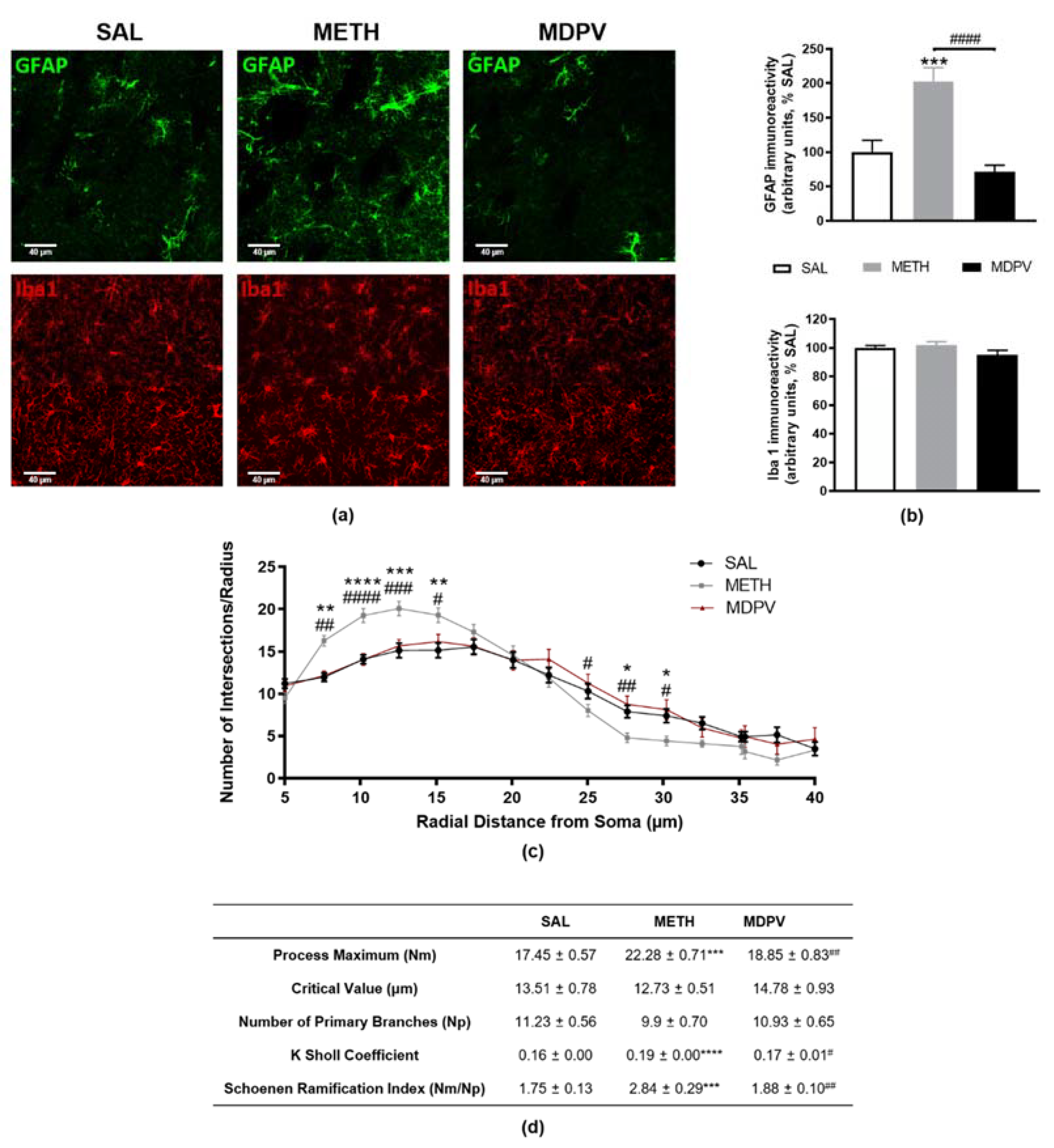
2.4. MPDV and METH on Striatal Glial Reactivity
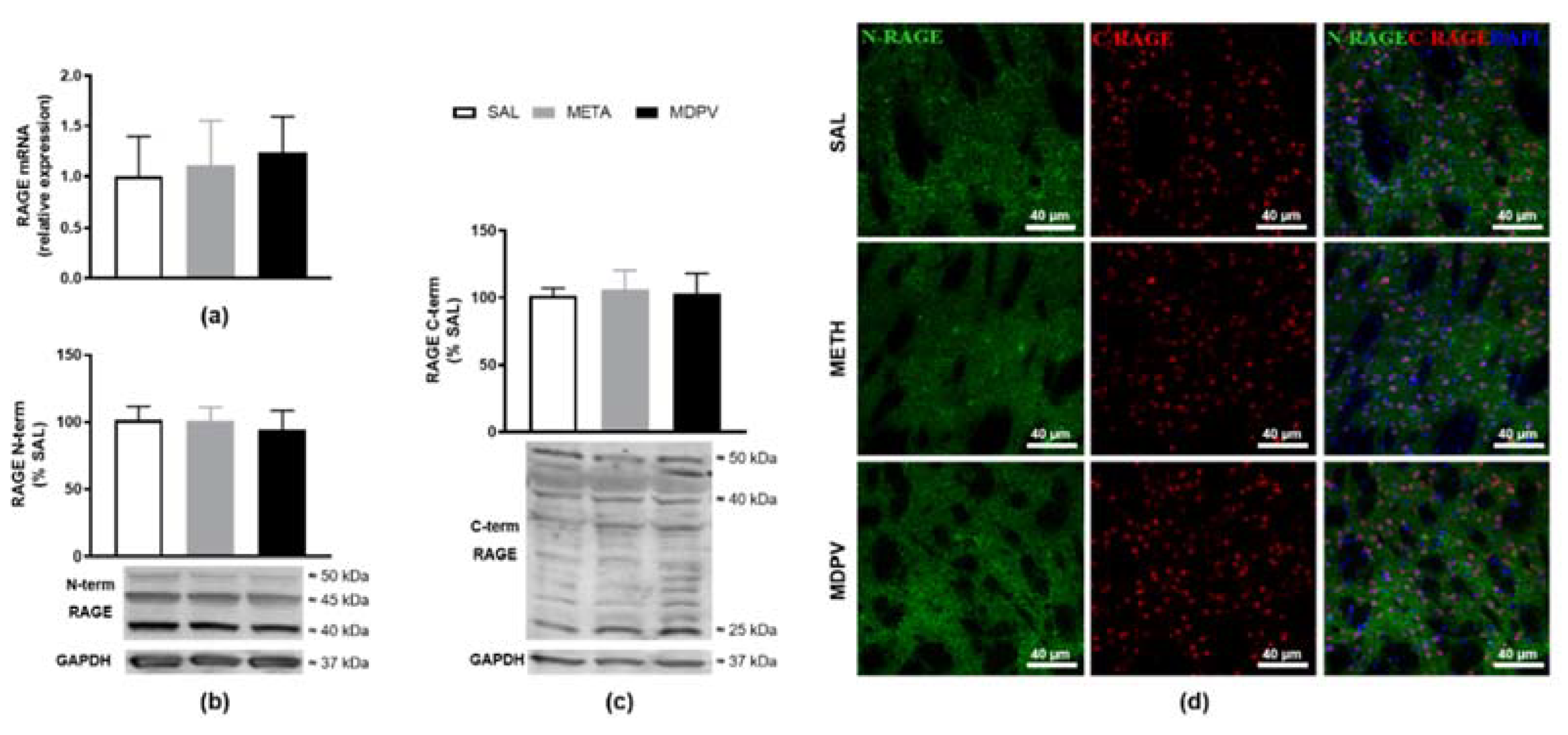
2.5. MPDV and METH on Immune Modulators RAGE and TLR
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Chemicals
4.3. Drug Administration
4.4. Behavioral Analysis
4.4.1. Elevated Plus-Maze
4.4.2. Open Field Test
4.4.3. Splash Test
4.4.4. Tail-Suspension Test
4.5. Tissue Collection and Processing
4.6. RT-qPCR Gene Expression
4.7. Western Blot
4.8. Immunohistochemistry
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Host | Dilution | Company | Catalog Number |
|---|---|---|---|---|
| Primary Antibodies | ||||
| Anti-GFAP | mouse | 1:100 | Merck Millipore | IF03L |
| Anti-TH | rabbit | 1:250 | Merck Millipore | AB152 |
| Anti-Iba-1 | rabbit | 1:250 | Wako | 019-19741 |
| Anti-C-RAGE | rabbit | 1:500 | Abcam | ab3611 |
| Anti-N-RAGE | goat | 1:1000 | Santa Cruz | Sc-8231 |
| Secondary Antibodies | ||||
| Alexa 488, anti-mouse | donkey | 1:1000 | ThermoFisherTM | A21202 |
| Alexa 488, anti-rabbit | goat | 1:1000 | Life Technologies | 1124089 |
| Alexa 594, anti-rabbit | donkey | 1:1000 | ThermoFisherTM | A21207 |
| Alexa 488, anti-goat | donkey | 1:1000 | Life Technologies | A11055 |
References
- European Monitoring Centre for Drugs and Drug Addiction. European Drug Report 2020: Trends and Developments; Publications Office of the European Union: Luxembourg, 2020. [Google Scholar]
- Goncalves, J.L.; Alves, V.L.; Aguiar, J.; Teixeira, H.M.; Camara, J.S. Synthetic cathinones: An evolving class of new psychoactive substances. Crit. Rev. Toxicol. 2019, 49, 549–566. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Mondola, R. 3,4-methylenedioxypyrovalerone (MDPV): Chemistry, pharmacology and toxicology of a new designer drug of abuse marketed online. Toxicol. Lett. 2012, 208, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.M.; Valente, M.J.; Carvalho, M.; da Silva, D.D.; Gaspar, H.; Carvalho, F.; de Lourdes Bastos, M.; de Pinho, P.G. Raising awareness of new psychoactive substances: Chemical analysis and in vitro toxicity screening of ‘legal high’ packages containing synthetic cathinones. Arch. Toxicol. 2015, 89, 757–771. [Google Scholar] [CrossRef]
- Hondebrink, L.; Zwartsen, A.; Westerink, R.H.S. Effect fingerprinting of new psychoactive substances (NPS): What can we learn from in vitro data? Pharmacol. Ther. 2018, 182, 193–224. [Google Scholar] [CrossRef] [PubMed]
- Glennon, R.A.; Young, R. Neurobiology of 3,4-methylenedioxypyrovalerone (MDPV) and alpha-pyrrolidinovalerophenone (alpha-PVP). Brain Res. Bull. 2016, 126, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Baumann, M.H.; Partilla, J.S.; Lehner, K.R. Psychoactive “bath salts”: Not so soothing. Eur. J. Pharmacol. 2013, 698, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, M.J.; de Pinho, P.G.; de Lourdes Bastos, M.; Carvalho, F.; Carvalho, M. Khat and synthetic cathinones: A review. Arch. Toxicol. 2014, 88, 15–45. [Google Scholar] [CrossRef]
- European Monitoring Centre for Drugs and Drug Addiction. EMCDDA–Europol Joint Report on a New Psychoactive Substance: MDPV (3,4-methylenedioxypyrovalerone); Joint Reports; Publications Office of the European Union: Luxembourg, 2014. [Google Scholar]
- Simmler, L.D.; Buser, T.A.; Donzelli, M.; Schramm, Y.; Dieu, L.H.; Huwyler, J.; Chaboz, S.; Hoener, M.C.; Liechti, M.E. Pharmacological characterization of designer cathinones in vitro. Br. J. Pharmacol. 2013, 168, 458–470. [Google Scholar] [CrossRef] [Green Version]
- Baumann, M.H.; Bukhari, M.O.; Lehner, K.R.; Anizan, S.; Rice, K.C.; Concheiro, M.; Huestis, M.A. Neuropharmacology of 3,4-Methylenedioxypyrovalerone (MDPV), Its Metabolites, and Related Analogs. Curr. Top. Behav. Neurosci. 2017, 32, 93–117. [Google Scholar] [CrossRef]
- Marusich, J.A.; Grant, K.R.; Blough, B.E.; Wiley, J.L. Effects of synthetic cathinones contained in “bath salts” on motor behavior and a functional observational battery in mice. Neurotoxicology 2012, 33, 1305–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, J.D.; Aarde, S.M.; Cole, M.; Vandewater, S.A.; Grant, Y.; Taffe, M.A. Locomotor Stimulant and Rewarding Effects of Inhaling Methamphetamine, MDPV, and Mephedrone via Electronic Cigarette-Type Technology. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2016, 41, 2759–2771. [Google Scholar] [CrossRef]
- Aarde, S.M.; Huang, P.K.; Creehan, K.M.; Dickerson, T.J.; Taffe, M.A. The novel recreational drug 3,4-methylenedioxypyrovalerone (MDPV) is a potent psychomotor stimulant: Self-administration and locomotor activity in rats. Neuropharmacology 2013, 71, 130–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, M.H.; Partilla, J.S.; Lehner, K.R.; Thorndike, E.B.; Hoffman, A.F.; Holy, M.; Rothman, R.B.; Goldberg, S.R.; Lupica, C.R.; Sitte, H.H.; et al. Powerful cocaine-like actions of 3,4-methylenedioxypyrovalerone (MDPV), a principal constituent of psychoactive ‘bath salts’ products. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2013, 38, 552–562. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.K.; Aarde, S.M.; Angrish, D.; Houseknecht, K.L.; Dickerson, T.J.; Taffe, M.A. Contrasting effects of d-methamphetamine, 3,4-methylenedioxymethamphetamine, 3,4-methylenedioxypyrovalerone, and 4-methylmethcathinone on wheel activity in rats. Drug Alcohol Depend. 2012, 126, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Fantegrossi, W.E.; Gannon, B.M.; Zimmerman, S.M.; Rice, K.C. In vivo effects of abused ‘bath salt’ constituent 3,4-methylenedioxypyrovalerone (MDPV) in mice: Drug discrimination, thermoregulation, and locomotor activity. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2013, 38, 563–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novellas, J.; Lopez-Arnau, R.; Carbo, M.L.; Pubill, D.; Camarasa, J.; Escubedo, E. Concentrations of MDPV in rat striatum correlate with the psychostimulant effect. J. Psychopharmacol. 2015, 29, 1209–1218. [Google Scholar] [CrossRef]
- Gannon, B.M.; Williamson, A.; Rice, K.C.; Fantegrossi, W.E. Role of monoaminergic systems and ambient temperature in bath salts constituent 3,4-methylenedioxypyrovalerone (MDPV)-elicited hyperthermia and locomotor stimulation in mice. Neuropharmacology 2018, 134, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Atehortua-Martinez, L.A.; Masniere, C.; Campolongo, P.; Chasseigneaux, S.; Callebert, J.; Zwergel, C.; Mai, A.; Laplanche, J.L.; Chen, H.; Etheve-Quelquejeu, M.; et al. Acute and chronic neurobehavioral effects of the designer drug and bath salt constituent 3,4-methylenedioxypyrovalerone in the rat. J. Psychopharmacol. 2019, 33, 392–405. [Google Scholar] [CrossRef] [PubMed]
- Aarde, S.M.; Huang, P.K.; Dickerson, T.J.; Taffe, M.A. Binge-like acquisition of 3,4-methylenedioxypyrovalerone (MDPV) self-administration and wheel activity in rats. Psychopharmacology 2015, 232, 1867–1877. [Google Scholar] [CrossRef] [Green Version]
- Magee, C.P.; German, C.L.; Siripathane, Y.H.; Curtis, P.S.; Anderson, D.J.; Wilkins, D.G.; Hanson, G.R.; Fleckenstein, A.E. 3,4-Methylenedioxypyrovalerone: Neuropharmacological Impact of a Designer Stimulant of Abuse on Monoamine Transporters. J. Pharmacol. Exp. Ther. 2020, 374, 273–282. [Google Scholar] [CrossRef]
- Duart-Castells, L.; Blanco-Gandia, M.C.; Ferrer-Perez, C.; Puster, B.; Pubill, D.; Minarro, J.; Escubedo, E.; Rodriguez-Arias, M. Cross-reinstatement between 3,4-methylenedioxypyrovalerone (MDPV) and cocaine using conditioned place preference. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 100, 109876. [Google Scholar] [CrossRef]
- Gatch, M.B.; Taylor, C.M.; Forster, M.J. Locomotor stimulant and discriminative stimulus effects of ‘bath salt’ cathinones. Behav. Pharmacol. 2013, 24, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Angoa-Perez, M.; Anneken, J.H.; Kuhn, D.M. Neurotoxicology of Synthetic Cathinone Analogs. Curr. Top. Behav. Neurosci. 2017, 32, 209–230. [Google Scholar] [CrossRef]
- Anneken, J.H.; Angoa-Perez, M.; Kuhn, D.M. 3,4-Methylenedioxypyrovalerone prevents while methylone enhances methamphetamine-induced damage to dopamine nerve endings: Beta-ketoamphetamine modulation of neurotoxicity by the dopamine transporter. J. Neurochem. 2015, 133, 211–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miner, N.B.; O’Callaghan, J.P.; Phillips, T.J.; Janowsky, A. The combined effects of 3,4-methylenedioxymethamphetamine (MDMA) and selected substituted methcathinones on measures of neurotoxicity. Neurotoxicol. Teratol. 2017, 61, 74–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwartsen, A.; Olijhoek, M.E.; Westerink, R.H.S.; Hondebrink, L. Hazard Characterization of Synthetic Cathinones Using Viability, Monoamine Reuptake, and Neuronal Activity Assays. Front. Neurosci. 2020, 14, 9. [Google Scholar] [CrossRef]
- Zwartsen, A.; Hondebrink, L.; Westerink, R.H. Neurotoxicity screening of new psychoactive substances (NPS): Effects on neuronal activity in rat cortical cultures using microelectrode arrays (MEA). Neurotoxicology 2018, 66, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Leong, H.S.; Philp, M.; Simone, M.; Witting, P.K.; Fu, S. Synthetic Cathinones Induce Cell Death in Dopaminergic SH-SY5Y Cells via Stimulating Mitochondrial Dysfunction. Int. J. Mol. Sci. 2020, 21, 1370. [Google Scholar] [CrossRef] [Green Version]
- Rosas-Hernandez, H.; Cuevas, E.; Lantz, S.M.; Rice, K.C.; Gannon, B.M.; Fantegrossi, W.E.; Gonzalez, C.; Paule, M.G.; Ali, S.F. Methamphetamine, 3,4-methylenedioxymethamphetamine (MDMA) and 3,4-methylenedioxypyrovalerone (MDPV) induce differential cytotoxic effects in bovine brain microvessel endothelial cells. Neurosci. Lett. 2016, 629, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Valente, M.J.; Bastos, M.L.; Fernandes, E.; Carvalho, F.; de Pinho, P.G.; Carvalho, M. Neurotoxicity of beta-Keto Amphetamines: Deathly Mechanisms Elicited by Methylone and MDPV in Human Dopaminergic SH-SY5Y Cells. ACS Chem. Neurosci. 2017, 8, 850–859. [Google Scholar] [CrossRef]
- Valente, M.J.; Amaral, C.; Correia-da-Silva, G.; Duarte, J.A.; Bastos, M.L.; Carvalho, F.; de Pinho, P.G.; Carvalho, M. Methylone and MDPV activate autophagy in human dopaminergic SH-SY5Y cells: A new insight into the context of beta-keto amphetamines-related neurotoxicity. Arch. Toxicol. 2017, 91, 3663–3676. [Google Scholar] [CrossRef]
- Coccini, T.; Vecchio, S.; Crevani, M.; De Simone, U. Cytotoxic Effects of 3,4-Catechol-PV (One Major MDPV Metabolite) on Human Dopaminergic SH-SY5Y Cells. Neurotox. Res. 2019, 35, 49–62. [Google Scholar] [CrossRef]
- Thomas, D.M.; Walker, P.D.; Benjamins, J.A.; Geddes, T.J.; Kuhn, D.M. Methamphetamine neurotoxicity in dopamine nerve endings of the striatum is associated with microglial activation. J. Pharmacol. Exp. Ther. 2004, 311, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Northcutt, A.L.; Cochran, T.A.; Zhang, X.; Fabisiak, T.J.; Haas, M.E.; Amat, J.; Li, H.; Rice, K.C.; Maier, S.F.; et al. Methamphetamine Activates Toll-Like Receptor 4 to Induce Central Immune Signaling within the Ventral Tegmental Area and Contributes to Extracellular Dopamine Increase in the Nucleus Accumbens Shell. ACS Chem. Neurosci. 2019, 10, 3622–3634. [Google Scholar] [CrossRef]
- Yang, T.; Zang, S.; Wang, Y.; Zhu, Y.; Jiang, L.; Chen, X.; Zhang, X.; Cheng, J.; Gao, R.; Xiao, H.; et al. Methamphetamine induced neuroinflammation in mouse brain and microglial cell line BV2: Roles of the TLR4/TRIF/Peli1 signaling axis. Toxicol. Lett. 2020, 333, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.M.; Rivera-Rodriguez, D.E.; Martinez, L.R. Methamphetamine alters the TLR4 signaling pathway, NF-kappaB activation, and pro-inflammatory cytokine production in LPS-challenged NR-9460 microglia-like cells. Mol. Immunol. 2020, 121, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.J.; Zhi, X.; Zhang, K.K.; Wang, L.B.; Li, J.H.; Liu, J.L.; Xu, L.L.; Yoshida, J.S.; Xie, X.L.; Wang, Q. Escalating dose-multiple binge methamphetamine treatment elicits neurotoxicity, altering gut microbiota and fecal metabolites in mice. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2021, 148, 111946. [Google Scholar] [CrossRef] [PubMed]
- Loftis, J.M.; Janowsky, A. Neuroimmune basis of methamphetamine toxicity. Int. Rev. Neurobiol. 2014, 118, 165–197. [Google Scholar] [CrossRef] [Green Version]
- Shaerzadeh, F.; Streit, W.J.; Heysieattalab, S.; Khoshbouei, H. Methamphetamine neurotoxicity, microglia, and neuroinflammation. J. Neuroinflamm. 2018, 15, 341. [Google Scholar] [CrossRef]
- Gadani, S.P.; Walsh, J.T.; Lukens, J.R.; Kipnis, J. Dealing with Danger in the CNS: The Response of the Immune System to Injury. Neuron 2015, 87, 47–62. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Li, J.X. Toll-Like Receptor 4 Signaling and Drug Addiction. Front. Pharmacol. 2020, 11, 603445. [Google Scholar] [CrossRef]
- Lwin, T.; Yang, J.L.; Ngampramuan, S.; Viwatpinyo, K.; Chancharoen, P.; Veschsanit, N.; Pinyomahakul, J.; Govitrapong, P.; Mukda, S. Melatonin ameliorates methamphetamine-induced cognitive impairments by inhibiting neuroinflammation via suppression of the TLR4/MyD88/NFkappaB signaling pathway in the mouse hippocampus. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 110109. [Google Scholar] [CrossRef]
- Ding, Q.; Keller, J.N. Evaluation of rage isoforms, ligands, and signaling in the brain. Biochim. Et Biophys. Acta 2005, 1746, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Viana, S.D.; Valero, J.; Rodrigues-Santos, P.; Couceiro, P.; Silva, A.M.; Carvalho, F.; Ali, S.F.; Fontes-Ribeiro, C.A.; Pereira, F.C. Regulation of striatal astrocytic receptor for advanced glycation end-products variants in an early stage of experimental Parkinson’s disease. J. Neurochem. 2016, 138, 598–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, F.C.; Cunha-Oliveira, T.; Viana, S.D.; Travassos, A.S.; Nunes, S.; Silva, C.; Prediger, R.D.; Rego, A.C.; Ali, S.F.; Ribeiro, C.A. Disruption of striatal glutamatergic/GABAergic homeostasis following acute methamphetamine in mice. Neurotoxicol. Teratol. 2012, 34, 522–529. [Google Scholar] [CrossRef]
- Edwards, S.; Koob, G.F. Experimental Psychiatric Illness and Drug Abuse Models: From Human to Animal, an Overview. In Psychiatric Disorders; Kobeissy, F.H., Ed.; Humana Press: Totowa, NJ, USA, 2012. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.M.; Kalasinsky, K.S.; Levey, A.I.; Bergeron, C.; Reiber, G.; Anthony, R.M.; Schmunk, G.A.; Shannak, K.; Haycock, J.W.; Kish, S.J. Striatal dopamine nerve terminal markers in human, chronic methamphetamine users. Nat. Med. 1996, 2, 699–703. [Google Scholar] [CrossRef]
- Silva, C.D.; Neves, A.F.; Dias, A.I.; Freitas, H.J.; Mendes, S.M.; Pita, I.; Viana, S.D.; de Oliveira, P.A.; Cunha, R.A.; Fontes Ribeiro, C.A.; et al. A single neurotoxic dose of methamphetamine induces a long-lasting depressive-like behaviour in mice. Neurotox. Res. 2014, 25, 295–304. [Google Scholar] [CrossRef]
- Davis, B.M.; Salinas-Navarro, M.; Cordeiro, M.F.; Moons, L.; De Groef, L. Characterizing microglia activation: A spatial statistics approach to maximize information extraction. Sci. Rep. 2017, 7, 1576. [Google Scholar] [CrossRef] [Green Version]
- Leitao, R.A.; Sereno, J.; Castelhano, J.M.; Goncalves, S.I.; Coelho-Santos, V.; Fontes-Ribeiro, C.; Castelo-Branco, M.; Silva, A.P. Aquaporin-4 as a New Target against Methamphetamine-Induced Brain Alterations: Focus on the Neurogliovascular Unit and Motivational Behavior. Mol. Neurobiol. 2018, 55, 2056–2069. [Google Scholar] [CrossRef] [PubMed]
- Coelho-Santos, V.; Leitao, R.A.; Cardoso, F.L.; Palmela, I.; Rito, M.; Barbosa, M.; Brito, M.A.; Fontes-Ribeiro, C.A.; Silva, A.P. The TNF-alpha/NF-kappaB signaling pathway has a key role in methamphetamine-induced blood-brain barrier dysfunction. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2015, 35, 1260–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grace, C.E.; Schaefer, T.L.; Herring, N.R.; Graham, D.L.; Skelton, M.R.; Gudelsky, G.A.; Williams, M.T.; Vorhees, C.V. Effect of a neurotoxic dose regimen of (+)-methamphetamine on behavior, plasma corticosterone, and brain monoamines in adult C57BL/6 mice. Neurotoxicol. Teratol. 2010, 32, 346–355. [Google Scholar] [CrossRef] [Green Version]
- Giannotti, G.; Canazza, I.; Caffino, L.; Bilel, S.; Ossato, A.; Fumagalli, F.; Marti, M. The Cathinones MDPV and alpha-PVP Elicit Different Behavioral and Molecular Effects Following Acute Exposure. Neurotox. Res. 2017, 32, 594–602. [Google Scholar] [CrossRef]
- Fonseca, R.; Carvalho, R.A.; Lemos, C.; Sequeira, A.C.; Pita, I.R.; Carvalho, F.; Silva, C.D.; Prediger, R.D.; Jarak, I.; Cunha, R.A.; et al. Methamphetamine Induces Anhedonic-Like Behavior and Impairs Frontal Cortical Energetics in Mice. CNS Neurosci. Ther. 2017, 23, 119–126. [Google Scholar] [CrossRef]
- Boger, H.A.; Middaugh, L.D.; Patrick, K.S.; Ramamoorthy, S.; Denehy, E.D.; Zhu, H.; Pacchioni, A.M.; Granholm, A.C.; McGinty, J.F. Long-term consequences of methamphetamine exposure in young adults are exacerbated in glial cell line-derived neurotrophic factor heterozygous mice. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 8816–8825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joca, L.; Zuloaga, D.G.; Raber, J.; Siegel, J.A. Long-term effects of early adolescent methamphetamine exposure on depression-like behavior and the hypothalamic vasopressin system in mice. Dev. Neurosci. 2014, 36, 108–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, C.G.; Whitfield, T.; Schulteis, G.; Koob, G.F.; Wee, S. A dysphoric-like state during early withdrawal from extended access to methamphetamine self-administration in rats. Psychopharmacology 2013, 225, 753–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolinsky, A.N.; Bergner, C.L.; LaPorte, J.L.; Kalueff, A.V. Analysis of Grooming Behavior and Its Utility in Studying Animal Stress, Anxiety, and Depression. In Mood and Anxiety Related Phenotypes in Mice; Gould, T., Ed.; Humana Press: Totowa, NJ, USA, 2009; Volume 42. [Google Scholar] [CrossRef]
- Pomrenze, M.B.; Tovar-Diaz, J.; Blasio, A.; Maiya, R.; Giovanetti, S.M.; Lei, K.; Morikawa, H.; Hopf, F.W.; Messing, R.O. A Corticotropin Releasing Factor Network in the Extended Amygdala for Anxiety. J. Neurosci. Off. J. Soc. Neurosci. 2019, 39, 1030–1043. [Google Scholar] [CrossRef] [PubMed]
- Duart-Castells, L.; Lopez-Arnau, R.; Buenrostro-Jauregui, M.; Munoz-Villegas, P.; Valverde, O.; Camarasa, J.; Pubill, D.; Escubedo, E. Neuroadaptive changes and behavioral effects after a sensitization regime of MDPV. Neuropharmacology 2019, 144, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philogene-Khalid, H.L.; Hicks, C.; Reitz, A.B.; Liu-Chen, L.Y.; Rawls, S.M. Synthetic cathinones and stereochemistry: S enantiomer of mephedrone reduces anxiety- and depressant-like effects in cocaine- or MDPV-abstinent rats. Drug Alcohol Depend. 2017, 178, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Zhu, J.P.; Xu, W.; Angulo, J.A. Disparity in the temporal appearance of methamphetamine-induced apoptosis and depletion of dopamine terminal markers in the striatum of mice. Brain Res. 2005, 1049, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Krasnova, I.N.; Justinova, Z.; Ladenheim, B.; Jayanthi, S.; McCoy, M.T.; Barnes, C.; Warner, J.E.; Goldberg, S.R.; Cadet, J.L. Methamphetamine self-administration is associated with persistent biochemical alterations in striatal and cortical dopaminergic terminals in the rat. PLoS ONE 2010, 5, e8790. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, J.P.; Miller, D.B. Neurotoxicity profiles of substituted amphetamines in the C57BL/6J mouse. J. Pharmacol. Exp. Ther. 1994, 270, 741–751. [Google Scholar] [PubMed]
- Granado, N.; Ares-Santos, S.; Tizabi, Y.; Moratalla, R. Striatal Reinnervation Process after Acute Methamphetamine-Induced Dopaminergic Degeneration in Mice. Neurotox. Res. 2018, 34, 627–639. [Google Scholar] [CrossRef]
- Friend, D.M.; Keefe, K.A. Glial reactivity in resistance to methamphetamine-induced neurotoxicity. J. Neurochem. 2013, 125, 566–574. [Google Scholar] [CrossRef] [Green Version]
- LaVoie, M.J.; Card, J.P.; Hastings, T.G. Microglial activation precedes dopamine terminal pathology in methamphetamine-induced neurotoxicity. Exp. Neurol. 2004, 187, 47–57. [Google Scholar] [CrossRef]
- Colon-Perez, L.M.; Pino, J.A.; Saha, K.; Pompilus, M.; Kaplitz, S.; Choudhury, N.; Jagnarine, D.A.; Geste, J.R.; Levin, B.A.; Wilks, I.; et al. Functional connectivity, behavioral and dopaminergic alterations 24 h following acute exposure to synthetic bath salt drug methylenedioxypyrovalerone. Neuropharmacology 2018, 137, 178–193. [Google Scholar] [CrossRef]
- Lopez-Arnau, R.; Duart-Castells, L.; Aster, B.; Camarasa, J.; Escubedo, E.; Pubill, D. Effects of MDPV on dopamine transporter regulation in male rats. Comparison with cocaine. Psychopharmacology 2019, 236, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Cameron, K.N.; Kolanos, R.; Solis, E., Jr.; Glennon, R.A.; De Felice, L.J. Bath salts components mephedrone and methylenedioxypyrovalerone (MDPV) act synergistically at the human dopamine transporter. Br. J. Pharmacol. 2013, 168, 1750–1757. [Google Scholar] [CrossRef] [Green Version]
- Ross, E.A.; Reisfield, G.M.; Watson, M.C.; Chronister, C.W.; Goldberger, B.A. Psychoactive “bath salts” intoxication with methylenedioxypyrovalerone. Am. J. Med. 2012, 125, 854–858. [Google Scholar] [CrossRef]
- Cruickshank, C.C.; Dyer, K.R. A review of the clinical pharmacology of methamphetamine. Addiction 2009, 104, 1085–1099. [Google Scholar] [CrossRef]
- Araujo, A.M.; Carvalho, M.; Costa, V.M.; Duarte, J.A.; Dinis-Oliveira, R.J.; Bastos, M.L.; Guedes de Pinho, P.; Carvalho, F. In vivo toxicometabolomics reveals multi-organ and urine metabolic changes in mice upon acute exposure to human-relevant doses of 3,4-methylenedioxypyrovalerone (MDPV). Arch. Toxicol. 2021, 95, 509–527. [Google Scholar] [CrossRef] [PubMed]
- Lacagnina, M.J.; Rivera, P.D.; Bilbo, S.D. Glial and Neuroimmune Mechanisms as Critical Modulators of Drug Use and Abuse. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2017, 42, 156–177. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Foster Olive, M.; Wang, K.K.; Febo, M.; Borsten, J.; Giordano, J.; Hauser, M.; Gold, M.S. Hypothesizing that designer drugs containing cathinones (“bath salts”) have profound neuro-inflammatory effects and dangerous neurotoxic response following human consumption. Med. Hypotheses 2013, 81, 450–455. [Google Scholar] [CrossRef]
- Fang, F.; Lue, L.F.; Yan, S.; Xu, H.; Luddy, J.S.; Chen, D.; Walker, D.G.; Stern, D.M.; Yan, S.; Schmidt, A.M.; et al. RAGE-dependent signaling in microglia contributes to neuroinflammation, Abeta accumulation, and impaired learning/memory in a mouse model of Alzheimer’s disease. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Lister, R.G. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology 1987, 92, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. JoVE 2015. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, T.A.; Blackman, A.V.; Oyrer, J.; Jayabal, S.; Chung, A.J.; Watt, A.J.; Sjostrom, P.J.; van Meyel, D.J. Neuronal morphometry directly from bitmap images. Nat. Methods 2014, 11, 982–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGill, B.E.; Barve, R.A.; Maloney, S.E.; Strickland, A.; Rensing, N.; Wang, P.L.; Wong, M.; Head, R.; Wozniak, D.F.; Milbrandt, J. Abnormal Microglia and Enhanced Inflammation-Related Gene Transcription in Mice with Conditional Deletion of Ctcf in Camk2a-Cre-Expressing Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2018, 38, 200–219. [Google Scholar] [CrossRef] [Green Version]






| PRR 1 | SAL | METH | MDPV |
|---|---|---|---|
| TLR2 | 100.0 ± 3.2 | 95.3 ± 7.0 | 100.6 ± 3.0 |
| TLR4 | 100.0 ± 11.3 | 101.6 ± 23.1 | 99.1 ± 15.7 |
| TLR7 | 100.0 ± 20.9 | 127.0 ± 32.8 | 90.7 ± 28.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campeão, M.; Fernandes, L.; Pita, I.R.; Lemos, C.; Ali, S.F.; Carvalho, F.; Rodrigues-Santos, P.; Fontes-Ribeiro, C.A.; Soares, E.; Viana, S.D.; et al. Acute MDPV Binge Paradigm on Mice Emotional Behavior and Glial Signature. Pharmaceuticals 2021, 14, 271. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030271
Campeão M, Fernandes L, Pita IR, Lemos C, Ali SF, Carvalho F, Rodrigues-Santos P, Fontes-Ribeiro CA, Soares E, Viana SD, et al. Acute MDPV Binge Paradigm on Mice Emotional Behavior and Glial Signature. Pharmaceuticals. 2021; 14(3):271. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030271
Chicago/Turabian StyleCampeão, Mafalda, Luciana Fernandes, Inês R. Pita, Cristina Lemos, Syed F. Ali, Félix Carvalho, Paulo Rodrigues-Santos, Carlos A. Fontes-Ribeiro, Edna Soares, Sofia D. Viana, and et al. 2021. "Acute MDPV Binge Paradigm on Mice Emotional Behavior and Glial Signature" Pharmaceuticals 14, no. 3: 271. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030271