
The Beta-Lactam Resistome Expressed by Aerobic and Anaerobic Bacteria Isolated from Human Feces of Healthy Donors

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
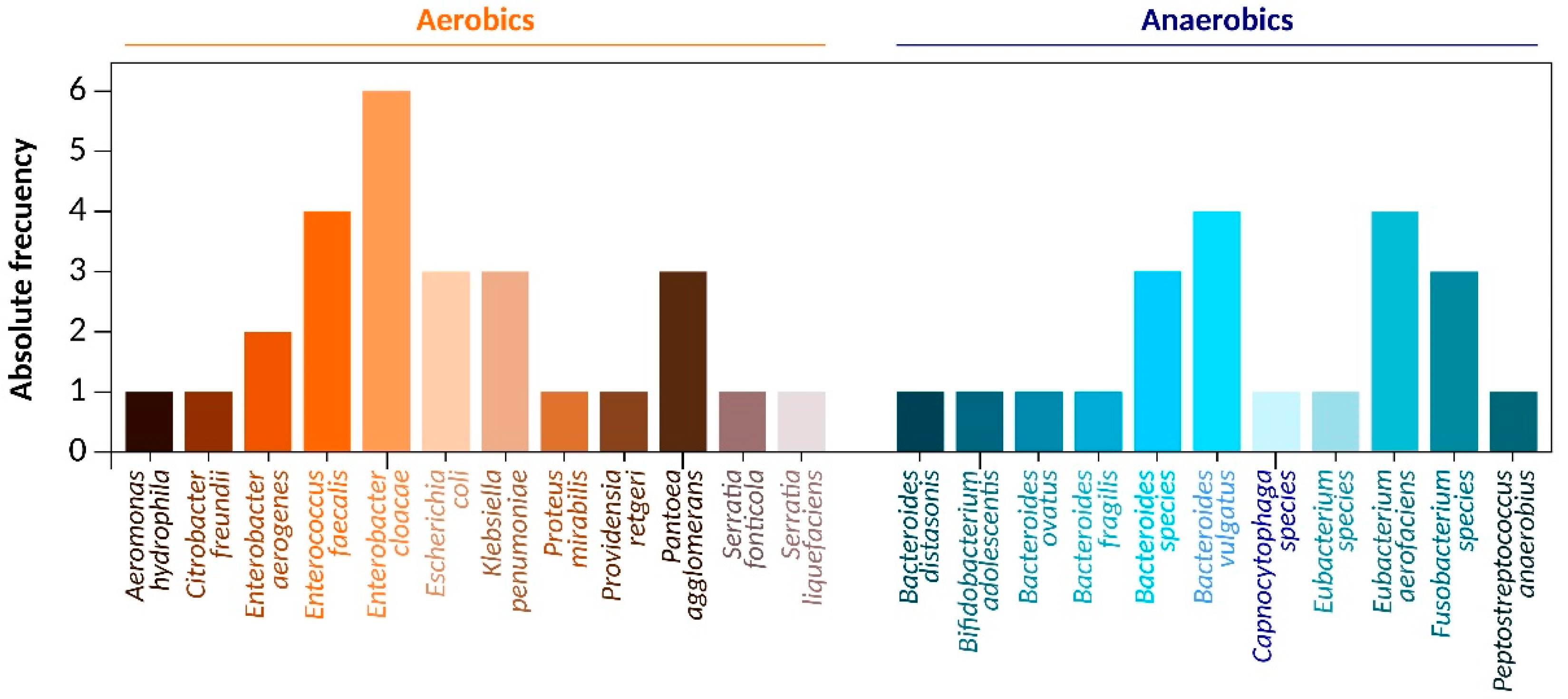
2.1. Bacteria Identification
2.2. Antimicrobial Susceptibility Testing
2.3. Beta-Lactamases Gene Family Identification
2.4. Beta-Lactamases Genotype
3. Discussion
3.1. Enterobacteriaceae Characterization
3.2. Beta-Lactam Gene Families
3.3. Beta-Lactam Phenotypes
4. Materials and Methods
4.1. Bioinformatic Analysis and Primer Design
4.2. Sampling
4.3. Bacterial Isolation
4.4. Biochemical Bacterial Identification
4.5. Antimicrobial Susceptibility Testing
4.6. DNA Extraction
4.6.1. Crude Extract
4.6.2. Genomic DNA Extraction
4.6.3. Plasmid DNA Extraction
4.7. Endpoint PCR
4.8. Whole and Plasmid DNA Sequencing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kardas, P.; Devine, S.; Golembesky, A.; Roberts, C. A systematic review and meta-analysis of misuse of antibiotic therapies in the community. Int. J. Antimicrob. Agents 2005, 26, 106–113. [Google Scholar] [CrossRef]
- Kadima, M.G. Concurrent Use of Traditional Medicine and Prescribed Antibiotics and/or Antiviral Medicines and Their Effects on Antimicrobial Resistance and Treatment Failure in Ilembe District, Kwazulu-Natal Province, South Africa. Master’s Thesis, University of KwaZulu-Natal, Scottsville, Pietermaritzburg, South Africa, January 2019. [Google Scholar]
- Huttner, A.; Harbarth, S.; Carlet, J.; Cosgrove, S.; Goossens, H.; Holmes, A.; Jarlier, V.; Voss, A.; Pittet, D. Antimicrobial resistance: A global view from the 2013 World Healthcare-Associated Infections Forum. Antimicrob. Resist. Infect. Control 2013, 2, 31. [Google Scholar] [CrossRef] [Green Version]
- Lucien, M.A.B.; Canarie, M.F.; Kilgore, P.E.; Jean-Denis, G.; Fénélon, N.; Pierre, M.; Cerpa, M.; Joseph, G.A.; Maki, G.; Zervos, M.J.; et al. Antibiotics and antimicrobial resistance in the COVID-19 era: Perspective from resource-limited settings. Int. J. Infect. Dis. 2021, 104, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Guo, J.; Jiang, Y.; Shao, Y. High concentration and high dose of disinfectants and antibiotics used during the COVID-19 pandemic threaten human health. Environ. Sci. Eur. 2021, 33, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. Epidemiology of β-lactamase-producing pathogens. Clin. Microbiol. Rev. 2020, 33, e00047-19. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Tapia, J.A. Antibiotic resistance, public health problem. An. Med. Asoc. Med. Hosp. ABC 2003, 48, 42–47. [Google Scholar]
- Kang, C.-I.; Kim, S.-H.; Park, W.B.; Lee, K.-D.; Kim, H.-B.; Kim, E.-C.; Oh, M.-D.; Choe, K.W. Bloodstream infections caused by antibiotic-resistant gram-negative bacilli: Risk factors for mortality and impact of inappropriate initial antimicrobial therapy on outcome. Antimicrob. Agents Chemother. 2005, 49, 760–766. [Google Scholar] [CrossRef] [Green Version]
- Zilberberg, M.D.; Shorr, A.F.; Micek, S.T.; Vazquez-Guillamet, C.; Kollef, M.H. Multi-drug resistance, inappropriate initial antibiotic therapy and mortality in Gram-negative severe sepsis and septic shock: A retrospective cohort study. Crit. Care 2014, 18, 596. [Google Scholar] [CrossRef] [PubMed]
- Fodor, A.; Abate, B.A.; Deák, P.; Fodor, L.; Gyenge, E.; Klein, M.G.; Koncz, Z.; Muvevi, J.; Ötvös, L.; Székely, G.; et al. Multidrug Resistance (MDR) and Collateral Sensitivity in Bacteria, with Special Attention to Genetic and Evolutionary Aspects and to the Perspectives of Antimicrobial Peptides—A Review. Pathogens 2020, 9, 522. [Google Scholar] [CrossRef]
- Kong, K.-F.; Schneper, L.; Mathee, K. Beta-lactam antibiotics: From antibiosis to resistance and bacteriology. APMIS 2010, 118, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Caruano, J.; Muccioli, G.G.; Robiette, R. Biologically active γ-lactams: Synthesis and natural sources. Org. Biomol. Chem. 2016, 14, 10134–10156. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Lim, D.Y.; Lee, K.A. Synthesis of A δ-Lactam Analogue of Penem Carboxylate. Bull. Korean Chem. Soc. 1990, 11, 361–362. [Google Scholar]
- Dutton, F.E.; Lee, B.H. Epsilon-lactam analogs of the anthelmintic cyclodepsipeptide PF1022A. Tetrahedron Lett. 1998, 39, 5313–5316. [Google Scholar] [CrossRef]
- Heesemann, J. Resistenzmechanismen gegen Betalaktamantibiotika. Infection 1993, 21, S4–S9. [Google Scholar] [CrossRef]
- Mathers, A.J.; Peirano, G.; Pitout, J.D.D. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coque, T.M.; Baquero, F.; Cantón, R. Increasing prevalence of ESBL-producing Enterobacteriaceae in Europe. Eurosurveillance 2008, 13, 19044. [Google Scholar] [CrossRef] [PubMed]
- Vatopoulos, A. High rates of metallo-beta-lactamase-producing Klebsiella pneumoniae in Greece—A review of the current evidence. Eurosurveillance 2008, 13, 7–8. [Google Scholar] [CrossRef]
- Brolund, A.; Sandegren, L. Characterization of ESBL disseminating plasmids. Infect. Dis. 2015, 48, 18–25. [Google Scholar] [CrossRef]
- De Angelis, G.; Del Giacomo, P.; Posteraro, B.; Sanguinetti, M.; Tumbarello, M. Molecular mechanisms, epidemiology, and clinical importance of β-lactam resistance in enterobacteriaceae. Int. J. Mol. Sci. 2020, 21, 5090. [Google Scholar] [CrossRef] [PubMed]
- Schultsz, C.; Geerlings, S. Plasmid-mediated resistance in enterobacteriaceae: Changing landscape and implications for therapy. Drugs 2012, 72, 1–16. [Google Scholar] [CrossRef] [PubMed]
- El Salabi, A.; Walsh, T.R.; Chouchani, C. Extended spectrum β-lactamases, carbapenemases and mobile genetic elements responsible for antibiotics resistance in Gram-negative bacteria. Crit. Rev. Microbiol. 2013, 39, 113–122. [Google Scholar] [CrossRef]
- Perez, F.; El Chakhtoura, N.G.; Papp-Wallace, K.M.; Wilson, B.M.; Bonomo, R.A. Treatment options for infections caused by carbapenem-resistant Enterobacteriaceae: Can we apply “precision medicine” to antimicrobial chemotherapy? Expert Opin. Pharm. 2016, 17, 761–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codjoe, F.S.; Donkor, E.S. Carbapenem Resistance: A Review. Med. Sci. 2017, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Mills, M.C.; Lee, J. The threat of carbapenem-resistant bacteria in the environment: Evidence of widespread contamination of reservoirs at a global scale. Environ. Pollut. 2019, 255, 113143. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-López, R.; Solano-Gálvez, S.; León-Chávez, B.A.; Thompson-Bonilla, M.R.; Guerrero-González, T.; Gómez-Conde, E.; Martínez-Fong, D.; González-Barrios, J.A. Characterization of gene families encoding beta-lactamases of gram-negative rods isolated from ready-to-eat vegetables in Mexico City. High. Throughput 2018, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauretti, L.; Riccio, M.L.; Mazzariol, A.; Cornaglia, G.; Amicosante, G.; Fontana, R.; Rossolini, G.M. Cloning and characterization of bla(VIM), a new integron-borne metallo- β-lactamase gene from a Pseudomonas aeruginosa clinical isolate. Antimicrob. Agents Chemother. 1999, 43, 1584–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de M. Rangel, W.; de Oliveira Longatti, S.M.; Ferreira, P.A.A.; Bonaldi, D.S.; Guimarães, A.A.; Thijs, S.; Weyens, N.; Vangronsveld, J.; Moreira, F.M.S. Leguminosae native nodulating bacteria from a gold mine As-contaminated soil: Multi-resistance to trace elements, and possible role in plant growth and mineral nutrition. Int. J. Phytoremediation 2017, 19, 925–936. [Google Scholar] [CrossRef]
- Sievert, D.M.; Ricks, P.; Edwards, J.R.; Schneider, A.; Patel, J.; Srinivasan, A.; Kallen, A.; Limbago, B.; Fridkin, S. National Healthcare Safety Network (NHSN) Team and Participating NHSN Facilities Antimicrobial-Resistant Pathogens Associated with Healthcare-Associated Infections Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–2010. Infect. Control Hosp. Epidemiol. 2013, 34, 1–14. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention Vital Signs: Carbapenem-Resistant Enterobacteriaceae. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm6209a3.htm (accessed on 12 April 2021).
- Ye, Q.; Wu, Q.; Zhang, S.; Zhang, J.; Yang, G.; Wang, J.; Xue, L.; Chen, M. Characterization of extended-spectrum β-lactamase-producing Enterobacteriaceae from retail food in China. Front. Microbiol. 2018, 9, 1709. [Google Scholar] [CrossRef]
- Babraham Bioinformatics—FastQC Babraham Bioinformatics—FastQC A Quality Control tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 12 April 2021).
- Burrows-Wheeler Aligner Burrows-Wheeler Aligner / [Bio-bwa-help] Citing bwa-mem. Available online: https://sourceforge.net/p/bio-bwa/mailman/message/30894287/ (accessed on 12 April 2021).
- CodonCode Corporation Sequence Assembly and Alignment with CodonCode Aligner. Available online: https://www.codoncode.com/aligner/ (accessed on 12 April 2021).
- System, K.T.S.C. Kraken Taxonomic Sequence Classification System. Available online: https://0-ccb-jhu-edu.brum.beds.ac.uk/software/kraken/ (accessed on 12 April 2021).
- BLAST BLAST: Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 12 April 2021).
- Hu, Y.; Yang, X.; Qin, J.; Lu, N.; Cheng, G.; Wu, N.; Pan, Y.; Li, J.; Zhu, L.; Wang, X.; et al. Metagenome-wide analysis of antibiotic resistance genes in a large cohort of human gut microbiota. Nat. Commun. 2013, 4, 2151. [Google Scholar] [CrossRef] [Green Version]
- Lagier, J.C.; Million, M.; Hugon, P.; Armougom, F.; Raoult, D. Human gut microbiota: Repertoire and variations. Front. Cell. Infect. Microbiol. 2012, 2, 136. [Google Scholar] [CrossRef] [Green Version]
- Parma, L.; Candela, M.; Soverini, M.; Turroni, S.; Consolandi, C.; Brigidi, P.; Mandrioli, L.; Sirri, R.; Fontanillas, R.; Gatta, P.P.; et al. Next-generation sequencing characterization of the gut bacterial community of gilthead sea bream (Sparus aurata, L.) fed low fishmeal based diets with increasing soybean meal levels. Anim. Feed Sci. Technol. 2016, 222, 204–216. [Google Scholar] [CrossRef]
- Tidjani Alou, M.; Lagier, J.-C.; Raoult, D. Diet influence on the gut microbiota and dysbiosis related to nutritional disorders. Hum. Microbiome J. 2016, 1, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Korzeniewska, E.; Harnisz, M. Beta-lactamase-producing Enterobacteriaceae in hospital effluents. J. Environ. Manag. 2013, 123, 1–7. [Google Scholar] [CrossRef]
- Bush, K. Alarming β-lactamase-mediated resistance in multidrug-resistant Enterobacteriaceae. Curr. Opin. Microbiol. 2010, 13, 558–564. [Google Scholar] [CrossRef]
- Joo, E.-J.; Kim, S.J.; Baek, M.; Choi, Y.; Seo, J.; Yeom, J.-S.; Ko, K.S. Fecal carriage of antimicrobial-resistant enterobacteriaceae in healthy Korean adults. J. Microbiol. Biotechnol. 2018, 28, 1178–1184. [Google Scholar] [CrossRef]
- Geser, N.; Stephan, R.; Korczak, B.M.; Beutin, L.; Hächler, H. Molecular identification of extended-spectrum-β-lactamase genes from Enterobacteriaceae isolated from healthy human carriers in Switzerland. Antimicrob. Agents Chemother. 2012, 56, 1609–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacace, D.; Fatta-Kassinos, D.; Manaia, C.M.; Cytryn, E.; Kreuzinger, N.; Rizzo, L.; Karaolia, P.; Schwartz, T.; Alexander, J.; Merlin, C.; et al. Antibiotic resistance genes in treated wastewater and in the receiving water bodies: A pan-European survey of urban settings. Water Res. 2019, 162, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Adesoji, A.T.; Ogunjobi, A.A. Detection of Extended Spectrum Beta-Lactamases Resistance Genes among Bacteria Isolated from Selected Drinking Water Distribution Channels in Southwestern Nigeria. Biomed. Res. Int. 2016, 2016, 7149295. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Gardea, M.C.E.; Tamez-Guerra, P.; Gomez-Flores, R.; de la Serna, F.J.Z.-D.; Eroza-de la Vega, G.; Nevárez-Moorillón, G.V.; Pérez-Recoder, M.C.; Sánchez-Ramírez, B.; González-Horta, M.D.C.; Infante-Ramírez, R. Multidrug-resistant bacteria isolated from surface water in Bassaseachic Falls National Park, Mexico. Int. J. Environ. Res. Public Health 2016, 13, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, D. β-lactam Resistance in Gram-negative Pathogens Isolated from Animals. Curr. Pharm. Des. 2012, 19, 239–249. [Google Scholar] [CrossRef]
- Chen, C.-M.; Ke, S.-C.; Li, C.-R.; Wu, Y.-C.; Chen, T.-H.; Lai, C.-H.; Wu, X.-X.; Wu, L.-T. High Diversity of Antimicrobial Resistance Genes, Class 1 Integrons, and Genotypes of Multidrug-Resistant Escherichia coli in Beef Carcasses. Microb. Drug Resist. 2017, 23, 915–924. [Google Scholar] [CrossRef]
- Heaton, J.C.; Jones, K. Microbial contamination of fruit and vegetables and the behaviour of enteropathogens in the phyllosphere: A review. J. Appl. Microbiol. 2008, 104, 613–626. [Google Scholar] [CrossRef]
- Gudda, F.O.; Waigi, M.G.; Odinga, E.S.; Yang, B.; Carter, L.; Gao, Y. Antibiotic-contaminated wastewater irrigated vegetables pose resistance selection risks to the gut microbiome. Environ. Pollut. 2020, 264, 114752. [Google Scholar] [CrossRef]
- Iwu, C.D.; Korsten, L.; Okoh, A.I. The incidence of antibiotic resistance within and beyond the agricultural ecosystem: A concern for public health. Microbiologyopen 2020, 9, e1035. [Google Scholar] [CrossRef] [PubMed]
- Tekiner, İ.H.; Özpınar, H. Occurrence and characteristics of extended spectrum beta-lactamases-producing enterobacteriaceae from foods of animal origin. Braz. J. Microbiol. 2016, 47, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesa, R.J.; Blanc, V.; Blanch, A.R.; Cortés, P.; González, J.J.; Lavilla, S.; Miró, E.; Muniesa, M.; Saco, M.; Tórtola, M.T.; et al. Extended-spectrum β-lactamase-producing Enterobacteriaceae in different environments (humans, food, animal farms and sewage). J. Antimicrob. Chemother. 2006, 58, 211–215. [Google Scholar] [CrossRef] [Green Version]
- Ruppé, E.; Andremont, A.; Armand-Lefèvre, L. Digestive tract colonization by multidrug-resistant Enterobacteriaceae in travellers: An update. Travel Med. Infect. Dis. 2018, 21, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Armand-Lefèvre, L.; Andremont, A.; Ruppé, E. Travel and acquisition of multidrug-resistant Enterobacteriaceae. Med. Mal. Infect. 2018, 48, 431–441. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Bacteria | Total Reads | Classified Reads | Domain | Phylum | Class | Order | Family | Genus | Species |
|---|---|---|---|---|---|---|---|---|---|
| Aerobic | |||||||||
| Aeromonas hydrophila | 4,502,897 | 4,412,839 (98%) | 4,103,940 (93%) | 3,734,586 (91%) | 3,211,744 (86%) | 2,697,865(84%) | 2,212,249 (82%) | 1,791,922 (81%) | 1,361,861 (76%) |
| Citrobacter freundii | 1,816,430 | 1,798,266 (99%) | 1,744,318 (97%) | 1,657,102 (95%) | 1,491,392 (90%) | 1, 327,339(89%) | 1,101,691 (83%) | 870,336 (79%) | 539,608 (62%) |
| Enterobacter aerogenes | 897,648 | 727,095 (81%) | 567,134 (78%) | 436,693 (77%) | 331,887 (76%) | 248,915 (75%) | 1817,08 (73%) | 130,830 (72%) | 91,581 (70%) |
| Enterococcus faecalis | 3,134,872 | 3,040,826 (97%) | 2,852,734 (91%) | 2,821,385 (90%) | 2,695,990 (86%) | 2,664,641 (85%) | 2,539,246 (81%) | 2,507,898 (80%) | 2,476,549 (79%) |
| Enterobacter cloacae | 945,761 | 898,473 (95%) | 892,798 (94%) | 871,046 (92%) | 866,317 (91%) | 850,239 (89%) | 842,673 (89%) | 836,998 (85%) | 835,107 (88%) |
| Escherichia coli | 2,361,827 | 2,078,408 (88%) | 1,936,698 (82%) | 1,865,843 (79%) | 1,606,042 (68%) | 1,511,569 (64%) | 1,464,333 (62%) | 1,440,714 (61%) | 1,393,478 (59%) |
| Klebsiella pneumoniae | 1,129,675 | 1,039,301 (92%) | 1,016,708 (90%) | 1,005,411 (89%) | 994,114 (88%) | 915,037 (81%) | 881,147 (78%) | 869,850 (77%) | 835,960 (74%) |
| Proteus mirabilis | 816,524 | 808,359 (99%) | 805,909 (98%) | 803,460 (97%) | 767,533 (94%) | 751,202 (92%) | 743,037 (91%) | 738,954 (90%) | 726,706 (89%) |
| Providencia rettgeri | 354,869 | 273,249 (77%) | 266,152 (75%) | 259,054 (73%) | 255,506 (72%) | 237,762 (67%) | 223,567 (63%) | 216,470 (61%) | 191,629 (54%) |
| Pantoea agglomerans | 497,802 | 438,066 (88%) | 433,088 (87%) | 408,198 (82%) | 393,264 (79%) | 388,286 (78%) | 353,439 (71%) | 338,505 (68%) | 333,527 (67%) |
| Serratia fonticola | 1,347,964 | 1,334,484 (99%) | 1,267,086 (94%) | 1,253,607 (93%) | 1,226,647 (91%) | 1,186,208 (88%) | 1,145,769 (85%) | 1,132,290 (84%) | 997,493 (74%) |
| Serratia liquefaciens | 256,789 | 190,024 (74%) | 182,320 (71%) | 179,752 (70%) | 174,617 (68%) | 159,209 (62%) | 156,641 (61%) | 154,073 (60%) | 133,530 (52%) |
| Anaerobic | |||||||||
| Bacteroides distasonis | 845,168 | 752,201 (89%) | 726,844 (86%) | 718,393 (85%) | 693,038 (82%) | 667,683 (79%) | 659,231 (78%) | 625,424 (74%) | 608,521 (72%) |
| Bifidobacterium adolescentis | 748,751 | 718,801 (96%) | 715,806 (95%) | 704,575 (94%) | 700,082 (93%) | 689,600 (92%) | 687,353 (91%) | 682,861 (91%) | 658,901 (88%) |
| Bacteroides ovatus | 965,743 | 905,867 (94%) | 902,004 (94%) | 898,141 (93%) | 893,312 (92%) | 888,484 (91%) | 830,539 (86%) | 743,622 (77%) | 637,390 (66%) |
| Bacteroides fragilis | 986,241 | 976,379 (99%) | 966,516 (98%) | 956,654 (97%) | 951,723 (96.4%) | 946,791 (96.1%) | 927,067 (94.3%) | 917,204 (93%) | 915,232 (92.6%) |
| Bacteroides species | 567,284 | 486,162 (85.7%) | 483,893 (85.3%) | 482,759 (85.1%) | 481,624 (84.9%) | 478,788 (84.4%) | 478,220 (84.3%) | 477,086 (84.1%) | 474,817 (83.7%) |
| Bacteroides vulgatus | 928,712 | 911,995 (98.2%) | 911,066 (98.1%) | 833,055 (89.7%) | 830,269 (89.4%) | 828,411 (89.2%) | 825,625 (88.9%) | 819,124 (88.2%) | 816,338 (87.9%) |
| Capnocytophaga species | 349,817 | 319,733 (91%) | 315,535 (90.2%) | 313,471 (89.6%) | 312,911 (89.4%) | 312,282 (89.2%) | 312,212 (89.1%) | 299,793 (85.7%) | 295,211 (84.3%) |
| Eubacterium species | 567,492 | 542,522 (95.6%) | 541,387 (95.4%) | 540,309 (95.2%) | 539,685 (95.1%) | 537,982 (94.8%) | 536,280 (94.5%) | 535,712 (94.4%) | 534,010 (94.1%) |
| Eubacterium aerofaciens | 814,927 | 620,974 (76.2%) | 616,085 (75.5%) | 604,676 (74.2%) | 599,786 (73.6%) | 595,712 (73.1%) | 594,082 (72.9%) | 581,043 (71.3%) | 576,968 (70.8%) |
| Fusobacterium species | 237,491 | 204,717 (86.2%) | 203,292 (85.6%) | 192,843 (81.2%) | 191,418 (80.6%) | 189,755 (79.9%) | 187,855 (79.1%) | 186,193 (78.4%) | 185,955 (78.3%) |
| Peptostreptococcus anaerobius | 729,358 | 686,691 (94.1%) | 686,399 (94.1%) | 683,044 (93.6%) | 682,314 (93.5%) | 679,251 (93.1%) | 677,282 (92.8%) | 612,442 (83.9%) | 587,498 (80.5%) |
| Bacteria | CDS | Number of Sequence Contigs | Genome Size (bp) | ||
|---|---|---|---|---|---|
| Assembled | Reported | Difference | |||
| Aerobic | |||||
| Aeromonas hydrophila | 4,428 | 1,124 | 5,124,487 | 4,911,246 | 213,241 |
| Citrobacter freundii | 5,064 | 789 | 5,343,952 | 5,297,052 | 46,900 |
| Enterobacter aerogenes | 4,545 | 1,484 | 5,578,724 | 5,280,350 | 298,374 |
| Enterococcus faecalis | 2,969 | 951 | 3,111,017 | 3,038,914 | 72,103 |
| Enterobacter cloacae | 4,545 | 1,484 | 4,982,176 | 4,772,910 | 209,266 |
| Escherichia coli | 5,704 | 1,561 | 5,689,156 | 5,615,389 | 73,767 |
| Klebsiella pneumoniae | 5,071 | 583 | 5,479,173 | 5,315,120 | 164,053 |
| Proteus mirabilis | 3,772 | 584 | 4,183,869 | 4,209,445 | 25,576 |
| Providencia rettgeri | 4,497 | 638 | 4,954,326 | 4,780,676 | 173,650 |
| Pantoea agglomerans | 4,255 | 738 | 4,090,220 | 4,047,712 | 42,508 |
| Serratia fonticola | 5,945 | 916 | 6,483,043 | 6,000,511 | 482,532 |
| Serratia liquefaciens | 4,936 | 495 | 5,706,987 | 5,395,544 | 311,443 |
| Anaerobic | |||||
| Bacteroides distasonis | 3,896 | 603 | 4,952,323 | 4,812,038 | 140,285 |
| Bifidobacterium adolescentis | 1,742 | 418 | 2,173,720 | 2,089,645 | 84,075 |
| Bacteroides ovatus | 4,803 | 756 | 6,425,267 | 6,472,489 | 47,222 |
| Bacteroides fragilis | 4,100 | 1,025 | 5,188,967 | 5,474,541 | 285,574 |
| Bacteroides species | 2,401 | 904 | 2,658,624 | 2,628,345 | 30,279 |
| Bacteroides vulgatus | 5,055 | 406 | 5,311,454 | 5,681,290 | 369,836 |
| Capnocytophaga species | 2,208 | 502 | 2,614,527 | 2,837,214 | 222,687 |
| Eubacterium species | 2,197 | 505 | 2,628,803 | 2,450,450 | 178,353 |
| Eubacterium aerofaciens | 1,887 | 1,548 | 2,128,754 | 2,264,854 | 136,100 |
| Fusobacterium species | 2,113 | 738 | 2,159,799 | 2,185,897 | 26,098 |
| Peptostreptococcus anaerobius | 1,793 | 847 | 1,989,753 | 2,106,123 | 116,370 |
| Antibiotic Families | Drugs | Aerobics | Anaerobic | ||
|---|---|---|---|---|---|
| Frequency | % | Frequency | % | ||
| Beta-Lactam | Amoxicillin/Clavulanic acid (AmC-30) | 24 | 88.9 | 12 | 54.5 |
| Piperacillin (PIP-100) | 17 | 63.0 | 16 | 72.7 | |
| Piperacillin/Tazobactam (TZP-110) | 5 | 18.5 | 6 | 27.3 | |
| Doxycycline (D-30) | 14 | 51.9 | 14 | 63.6 | |
| Ampicillin 10(AM-10) | 24 | 88.9 | 12 | 54.5 | |
| Cephalothin 1° (CF-30) | 19 | 70.4 | 16 | 72.7 | |
| Cefazolin 1° (CZ-30) | 18 | 66.7 | 15 | 68.2 | |
| Cefaclor 2° (CEC-30) | 18 | 66.7 | 16 | 72.7 | |
| Cefuroxime 2° (CXM-30) | 19 | 70.4 | 16 | 72.7 | |
| Cefixime 3° (CFM-5) | 22 | 81.5 | 18 | 81.8 | |
| Ceftizoxime 3a (ZOX-30) | 4 | 14.8 | 10 | 45.5 | |
| Meropenem (MEM-10) | 3 | 11.1 | 1 | 4.5 | |
| Imipenem (IPM-10) | 3 | 11.1 | 1 | 4.5 | |
| Gene Family | Positive Strains (n) | % | Gene Family | Positive Strains (n) | % |
|---|---|---|---|---|---|
| blaTEM | 49 | 100.0 | blaBIL | 3 | 6.1 |
| blaSHV | 5 | 10.2 | blaP | 1 | 2.0 |
| blaKPC | 5 | 10.2 | blaCFX | 4 | 8.2 |
| blaCYM | 7 | 14.3 |
| Aerobic Bacteria | Beta-Lactamases Families | Anaerobic Bacteria | Beta-Lactamases Families | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TEM | SHV | KPC | CYM | BIL | CFX | TEM | CYM | BIL | P | ||
| Aeromonas hydrophila | + | + | + | Capnocytophaga species | + | ||||||
| Citrobacter freundii | + | Bacteroides distasonis | + | + | |||||||
| Proteus mirabilis | + | + | + | + | Bifidobacterium adolescenti | + | |||||
| Providencia rettgeri | + | Bacteroides ovatus | + | ||||||||
| Serratia fonticola | + | + | Bacteroides fragilis | + | + | ||||||
| Serratia liquefaciens | + | Eubacterium species | + | ||||||||
| Enterobacter aerogenes | + | + | Eubacterium aerofaciens | + | |||||||
| Escherichia coli | + | + | + | + | Peptostreptocuccus anaerobius | + | |||||
| Klebsiella pneumoniae | + | + | + | + | + | Fusobacterium species | + | + | |||
| Pantoea agglomerans | + | + | Bacteroides vulgatus | + | + | ||||||
| Enterococcus faecalis | + | + | Bacteroides species | + | |||||||
| Enterobacter cloacae | + | + | + | + | + | ||||||
| Bacteria | Phenotype of Beta-Lactamases | ||||||
|---|---|---|---|---|---|---|---|
| TEM | SHV | KPC | CYM | BIL | CFX | P | |
| Aerobic | |||||||
| Aeromonas hydrophila | blaTEM-1 | blaSHV-12 | blaBIL-1 | ||||
| Citrobacter freundii | blaTEM-2 | ||||||
| Enterobacter aerogenes | blaTEM-1 | blaKPC-2 | |||||
| Enterococcus faecalis | blaTEM-1 | blaSHV-1 | |||||
| Enterobacter cloacae | blaTEM-12 | blaSHV-1 | |||||
| Escherichia coli | blaTEM-52 | blaSHV-12 | blaCMY-2 | blaCTX-M-15 | |||
| Klebsiella pneumoniae | blaTEM-10 | blaSHV-12 | blaKPC-3 | blaCMY-2 | blaCTX-M-15 | ||
| Proteus mirabilis | blaTEM-1 | blaKPC-2 | blaCMY-2 | blaCTX-M-15 | |||
| Providencia rettgeri | blaTEM-24 | ||||||
| Pantoea agglomerans | blaTEM-1 | blaCMY-2 | |||||
| Serratia fonticola | blaTEM-1 | blaKPC-2 | |||||
| Serratia liquefaciens | blaTEM-12 | ||||||
| Anaerobic | |||||||
| Bacteroides distasonis | blaTEM-1 | blaBIL-1 | |||||
| Bifidobacterium adolescentis | blaTEM-1 | ||||||
| Bacteroides ovatus | blaTEM-1 | ||||||
| Bacteroides fragilis | blaTEM-2 | blaCMY-2 | |||||
| Bacteroides species | blaTEM-1 | ||||||
| Bacteroides vulgatus | blaTEM-1 | blaCMY-2 | |||||
| Capnocytophaga species | blaTEM-1 | ||||||
| Eubacterium species | blaTEM-1 | ||||||
| Eubacterium aerofaciens | blaTEM-1 | ||||||
| Fusobacterium species | blaTEM-12 | blaP-1 | |||||
| Peptostreptococcus anaerobius | blaTEM-1 | ||||||
| Gene Family | Primer Name | Primer Sequence (5’ to 3’) | Tm (°C) | Position | Amplicon (bp) |
|---|---|---|---|---|---|
| blaOXA | BlaOXA-FW | GGTTTCGGTAATGCTGAAATTGG | 61.18 | 214–236 | 114 |
| BlaOXA-RW | GCTGTGTATGTGCTAATTGGGA | 61.19 | 327–306 | ||
| blaVIM | BlaVIM-FW | CGACAGTCARCGAAATTCC | 61.39 | 105–123 | 133 |
| BlaVIM-RW | CAATGGTCTSATTGTCCGTG | 61.34 | 238–219 | ||
| blaSHV | BlaSHV-FW1 | CGTAGGCATGATAGAAATGGATC | 61.04 | 133–155 | 106 |
| BlaSHV-RW1 | CGCAGAGCACTACTTTAAAGG | 61.33 | 239–218 | ||
| BlaSHV-FW2 | GCCTCATTCAGTTCCGTTTC | 61.62 | 399–418 | 141 | |
| BlaSHV-RW2 | CCATTACCATGAGCGATAACAG | 61.22 | 540–518 | ||
| blaTEM | BlaTEM-FW | GCCAACTTACTTCTGACAACG | 61.80 | 1699–1719 | 213 |
| BlaTEM-RW | CGTTTGGAATGGCTTCATTC | 60.13 | 1912–1892 | ||
| blaIMP | BlaIMP-FW1 | GGAATAGARTGGCTTAAYTCTCG | 60.92 | 319–332 | 183 |
| BlaIMP-RW1 | CYASTASGTTATCTKGAGTGTG | 62.45 | 502–480 | ||
| BlaIMP-FW2 | GGTGGAATAGARTGGCTTAAYTC | 61.11 | 316–339 | 192 | |
| BlaIMP-RW2 | CCAAACCACTACGTTATCTKGAG | 61.29 | 508–485 | ||
| blaROB | BlaROB-FW | CCAACATCGTGGAAAGTGTAG | 61.27 | 718–739 | 126 |
| BlaROB-RW | GTAAATTGCGTACTCATGATTGC | 60.90 | 844–821 | ||
| blaKPC | BlaKPC-FW | GCTAAACTCGAACAGGACTTTG | 61.79 | 100–121 | 117 |
| BlaKPC-RW | CTTGAATGAGCTGCACAGTG | 61.90 | 216–197 | ||
| blaCTX | BlaCTX-FW1 | GATACCGCAGATAATACGCAG | 60.79 | 161–181 | 116 |
| BlaCTX-RW1 | CGTTTTGCGTTTCACTCTG | 60.28 | 276–258 | ||
| BlaCTX-FW2 | GCTGATTCTGGTCACTTACTTC | 61.02 | 789–810 | 83 | |
| BlaCTX-RW2 | CGCCGACGCTAATACATC | 60.69 | 855–872 | ||
| BlaCTX-FW3 | CTGCTTAACTACAATCCSATTGC | 62.17 | 314–336 | 226 | |
| BlaCTX-RW3 | GGAATGGCGGTATTKAGC | 60.86 | 539–522 | ||
| blaCMY | BlaCMY-FW1 | GTTTGAGCTAGGATCGGTTAG | 60.25 | 337–357 | 123 |
| BlaCMY-RW1 | CTGTTTGCCTGTCAGTTCTG | 61.48 | 460–441 | ||
| BlaCMY-FW2 | GAACGAAGGCTACGTAGCT | 61.71 | 213–231 | 160 | |
| BlaCMY-RW2 | CTGAAACGTGATTCGATCATCA | 61.08 | 372–351 | ||
| blaDHA | BlaDHA-FW1 | GCATATTGATCTGCATATCTCCAC | 61.60 | 399–422 | 200 |
| BlaDHA-RW1 | GCTGCTGTAACTGTTCTGC | 61.62 | 598–580 | ||
| BlaDHA-FW2 | GCGGATCTGCTGAATTTCTATC | 61.54 | 464–485 | 147 | |
| BlaDHA-RW2 | GCAGTCAGCAACTGCTCATAC | 61.05 | 610–591 | ||
| BlaDHA-FW3 | GTAAGATTCCGCATCAAGCTG | 61.74 | 430–450 | 117 | |
| BlaDHA-RW3 | GGGTTATCTCACACCTTTATTACTG | 61.08 | 546–522 | ||
| blaP | BlaP-FW | GGAGAATATTGGGATTACAATGGC | 61.74 | 271–294 | 204 |
| BlaP-RW | CGCATCATCGAGTGTGATTG | 61.80 | 474–455 | ||
| blaCFX | BlaCFX-FW | CCAGTCATATCATTGACAGTGAG | 60.86 | 437–459 | 177 |
| BlaCFX-RW | GACATTTCCTCTTCCGTATAAGC | 61.16 | 613–591 | ||
| blaLAP | BlaLAP-FW | AGGGCTTGAACAACTTGAAC | 61.07 | 249–268 | 126 |
| BlaLAP-RW | GTAATGGCAGCATTGCATAAC | 60.59 | 374–354 | ||
| blaBIL | BlaBIL-FW | GCCGATATCGTTAATCGCAC | 61.65 | 100–119 | 128 |
| BlaBIL-RW | GTTATTGGCGATATCGGCTTTA | 60.98 | 227–206 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez-López, R.; Solano-Gálvez, S.; Álvarez-Hernández, D.A.; Ascencio-Aragón, J.A.; Gómez-Conde, E.; Piña-Leyva, C.; Lara-Lozano, M.; Guerrero-González, T.; González-Barrios, J.A. The Beta-Lactam Resistome Expressed by Aerobic and Anaerobic Bacteria Isolated from Human Feces of Healthy Donors. Pharmaceuticals 2021, 14, 533. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060533
Vázquez-López R, Solano-Gálvez S, Álvarez-Hernández DA, Ascencio-Aragón JA, Gómez-Conde E, Piña-Leyva C, Lara-Lozano M, Guerrero-González T, González-Barrios JA. The Beta-Lactam Resistome Expressed by Aerobic and Anaerobic Bacteria Isolated from Human Feces of Healthy Donors. Pharmaceuticals. 2021; 14(6):533. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060533
Chicago/Turabian StyleVázquez-López, Rosalino, Sandra Solano-Gálvez, Diego Abelardo Álvarez-Hernández, Jorge Alberto Ascencio-Aragón, Eduardo Gómez-Conde, Celia Piña-Leyva, Manuel Lara-Lozano, Tayde Guerrero-González, and Juan Antonio González-Barrios. 2021. "The Beta-Lactam Resistome Expressed by Aerobic and Anaerobic Bacteria Isolated from Human Feces of Healthy Donors" Pharmaceuticals 14, no. 6: 533. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060533