
Evaluation of Dexamethasone-Induced Osteoporosis In Vivo Using Zebrafish Scales

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Intraperitoneal Dexamethasone Injection Reduced the Regeneration of Zebrafish Scales
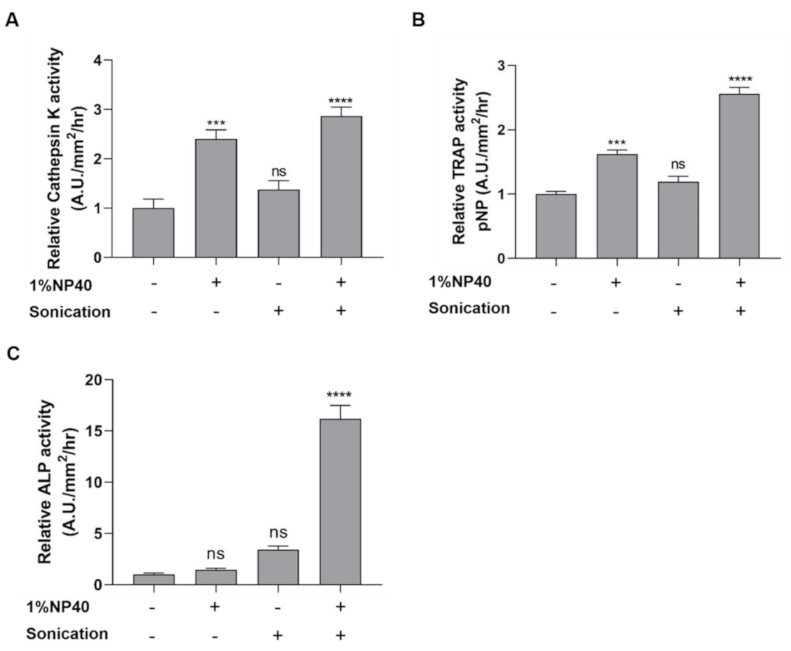
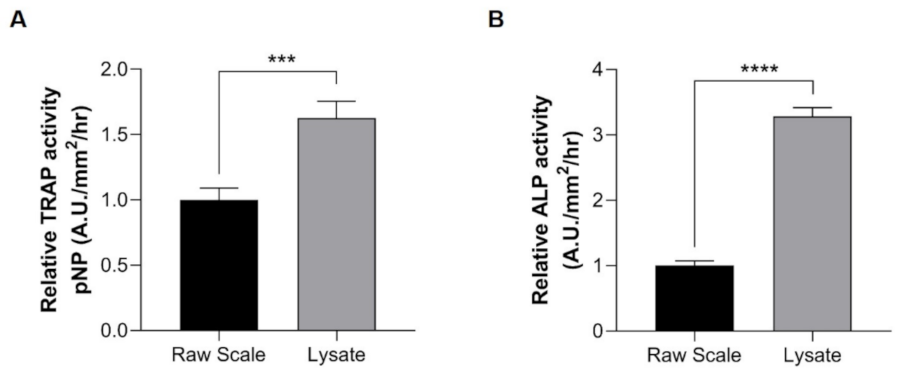
2.2. Lysate from Scales Is Beneficial for the Measurement of Cathepsin K, TRAP, and ALP Activities
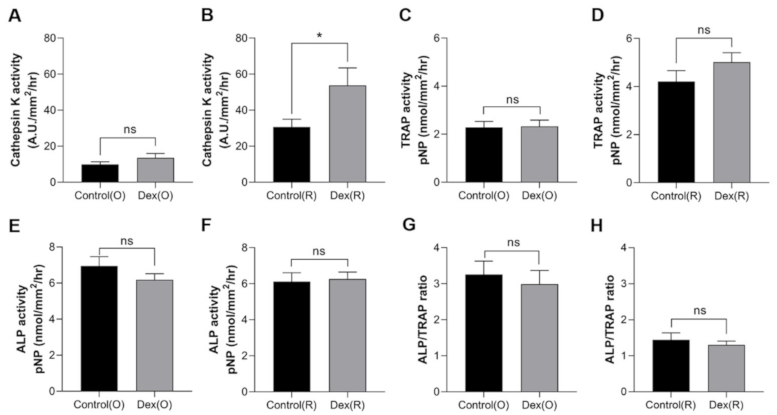
2.3. Effect of Dexamethasone on Cathepsin K, TRAP and ALP Activities
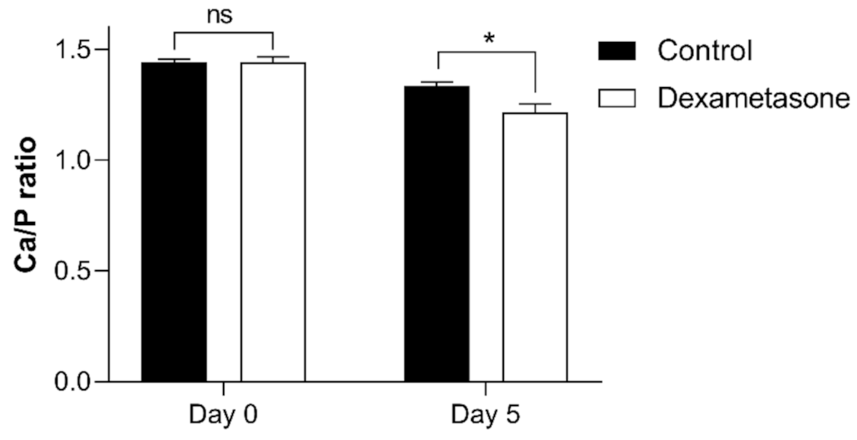
2.4. Effect of Dexamethasone on the Mineral Composition of Zebrafish Scales
3. Discussion
3.1. Dexamethasone-Induced Osteoporosis Using Zebrafish Scales
3.2. Our Developed Assay Allows the Effective Measurement of the Activities of Three Bone-Related Enzymes
4. Materials and Methods
4.1. Zebrafish Husbandry
4.2. Dexamethasone Treatment and Sample Collection
4.3. Morphometric Analysis
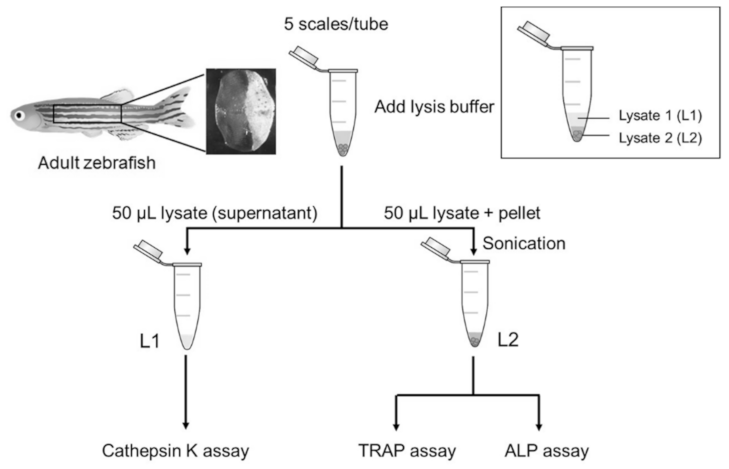
4.4. Scale Lysis Method
4.5. Biochemical Cathepsin K, TRAP, and ALP Scale Assays
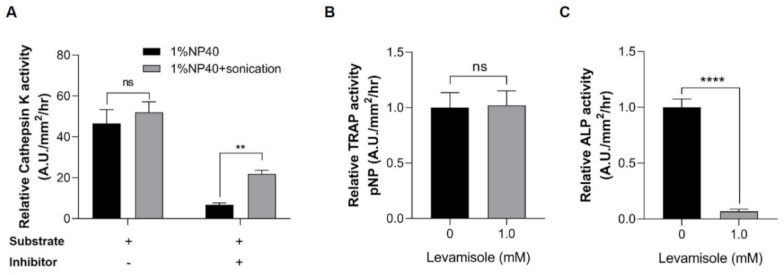
4.6. Inhibitor Specificity of Cathepsin K, TRAP and ALP in Zebrafish Scale Lysates
4.7. Mineral Content Determination Using the Inductively Coupled Plasma Mass Spectrometry (ICP-MS) Method
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Osterhoff, G.; Morgan, E.F.; Shefelbine, S.J.; Karim, L.; McNamara, L.M.; Augat, P. Bone mechanical properties and changes with osteoporosis. Injury 2016, 47 (Suppl. 2), S11–S20. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M. Aging mechanisms in bone. Bonekey Rep. 2012, 1, 102. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; McDonald, J.M. Disorders of bone remodeling. Annu. Rev. Pathol. 2011, 6, 121–145. [Google Scholar] [CrossRef] [Green Version]
- Guido, G.; Scaglione, M.; Fabbri, L.; Ceglia, M.J. The “osteoporosis disease”. Clin. Cases Min. Bone Metab. 2009, 6, 114–116. [Google Scholar]
- Briot, K.; Roux, C. Glucocorticoid-induced osteoporosis. RMD Open 2015, 1, e000014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutinho, A.E.; Chapman, K.E. The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights. Mol. Cell Endocrinol. 2011, 335, 2–13. [Google Scholar] [CrossRef]
- Marques, I.J.; Lupi, E.; Mercader, N. Model systems for regeneration: Zebrafish. Development 2019, 146, dev167692. [Google Scholar] [CrossRef] [Green Version]
- Hsu, E.; Nanes, M. Advances in treatment of glucocorticoid-induced osteoporosis. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 411–417. [Google Scholar] [CrossRef]
- Walsh, L.J.; Lewis, S.A.; Wong, C.A.; Cooper, S.; Oborne, J.; Cawte, S.A.; Harrison, T.; Green, D.J.; Pringle, M.; Hubbard, R.; et al. The impact of oral corticosteroid use on bone mineral density and vertebral fracture. Am. J. Respir. Crit. Care Med. 2002, 166, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Henneicke, H.; Gasparini, S.J.; Brennan-Speranza, T.C.; Zhou, H.; Seibel, M.J. Glucocorticoids and bone: Local effects and systemic implications. Trends Endocrinol. Metab. 2014, 25, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-Y.; Chiang, C.-Y.; Tsai, H.-J. Zebrafish and Medaka: New model organisms for modern biomedical research. J. Biomed Sci. 2016, 23, 19. [Google Scholar] [CrossRef] [Green Version]
- Schartl, M. Beyond the zebrafish: Diverse fish species for modeling human disease. Dis. Models Mech. 2014, 7, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Veldman, M.B.; Lin, S. Zebrafish as a developmental model organism for pediatric research. Pediatric Res. 2008, 64, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, H.; Kim, J.H.; Kim, C.Y.; Hwang, S.; Kim, H.; Yang, S.; Lee, J.E.; Lee, I. Function-driven discovery of disease genes in zebrafish using an integrated genomics big data resource. Nucleic Acids Res. 2016, 44, 9611–9623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.B.; Westerfield, M. Zebrafish models in translational research: Tipping the scales toward advancements in human health. Dis. Model Mech. 2014, 7, 739–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vrieze, E.; Sharif, F.; Metz, J.; Flik, G.; Richardson, M. Matrix metalloproteinases in osteoclasts of ontogenetic and regenerating zebrafish scales. Bone 2011, 48, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, N.; Ura, K.; Takagi, Y. Scale calcification in the goldfish in vitro: Histological and quantitative analysis. Fish. Sci. 2010, 76, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Sire, J.Y.; Allizard, F.; Babiar, O.; Bourguignon, J.; Quilhac, A. Scale development in zebrafish (Danio rerio). J. Anat. 1997, 190 Pt 4, 545–561. [Google Scholar] [CrossRef]
- Sire, J.Y.; Akimenko, M.A. Scale development in fish: A review, with description of sonic hedgehog (shh) expression in the zebrafish (Danio rerio). Int. J. Dev. Biol. 2003, 48, 233–247. [Google Scholar] [CrossRef]
- Pasqualetti, S.; Banfi, G.; Mariotti, M. The zebrafish scale as model to study the bone mineralization process. J. Mol. Histol. 2012, 43, 589–595. [Google Scholar] [CrossRef] [PubMed]
- de Vrieze, E.; Moren, M.; Metz, J.R.; Flik, G.; Lie, K.K. Arachidonic acid enhances turnover of the dermal skeleton: Studies on zebrafish scales. PLoS ONE 2014, 9, e89347. [Google Scholar] [CrossRef] [Green Version]
- Sire, J.-Y. The same cell lineage is involved in scale formation and regeneration in the teleost fish Hemichromis bimaculatus. Tissue Cell 1989, 21, 447–462. [Google Scholar] [CrossRef]
- Thamamongood, T.A.; Furuya, R.; Fukuba, S.; Nakamura, M.; Suzuki, N.; Hattori, A. Expression of osteoblastic and osteoclastic genes during spontaneous regeneration and autotransplantation of goldfish scale: A new tool to study intramembranous bone regeneration. Bone 2012, 50, 1240–1249. [Google Scholar] [CrossRef]
- de Vrieze, E.; van Kessel, M.; Peters, H.; Spanings, F.; Flik, G.; Metz, J. Prednisolone induces osteoporosis-like phenotype in regenerating zebrafish scales. Osteoporos. Int. 2014, 25, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Wang, J.; Wu, M.; Ma, Y.; Wang, S.; Su, Q. Preventive Effects of Evodiamine on Dexamethasone-Induced Osteoporosis in Zebrafish. BioMed Res. Int. 2019, 2019, 5859641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttgereit, F.; Brand, M.D.; Burmester, G.-R. Equivalent doses and relative drug potencies for non-genomic glucocorticoid effects: A novel glucocorticoid hierarchy. Biochem. Pharmacol. 1999, 58, 363–368. [Google Scholar] [CrossRef]
- Kuo, T.-R.; Chen, C.-H. Bone biomarker for the clinical assessment of osteoporosis: Recent developments and future perspectives. Biomark. Res. 2017, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Halleen, J.M.; Alatalo, S.L.; Suominen, H.; Cheng, S.; Janckila, A.J.; Väänänen, H.K. Tartrate-resistant acid phosphatase 5b: A novel serum marker of bone resorption. J. Bone Miner. Res. 2000, 15, 1337–1345. [Google Scholar] [CrossRef]
- Bühling, F.; Reisenauer, A.; Gerber, A.; Krüger, S.; Weber, E.; Brömme, D.; Roessner, A.; Ansorge, S.; Welte, T.; Röcken, C. Cathepsin K–a marker of macrophage differentiation? J. Pathol. 2001, 195, 375–382. [Google Scholar] [CrossRef]
- Hou, W.S.; Li, Z.; Gordon, R.E.; Chan, K.; Klein, M.J.; Levy, R.; Keysser, M.; Keyszer, G.; Brömme, D. Cathepsin k is a critical protease in synovial fibroblast-mediated collagen degradation. Am. J. Pathol. 2001, 159, 2167–2177. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Hou, W.-S.; Brömme, D. Collagenolytic activity of cathepsin k is specifically modulated by cartilage-resident chondroitin sulfates. Biochemistry 2000, 39, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Bergen, D.; Kague, E.; Hammond, C. Zebrafish as an emerging model for osteoporosis: A primary testing platform for screening new osteo-active compounds. Front. Endocrinol. (Lausanne) 2019, 10, 1–20. [Google Scholar] [CrossRef] [Green Version]
- de Vrieze, E.; Metz, J.; von den Hoff, J.; Flik, G. ALP, TRAcP and cathepsin K in elasmoid scales: A role in mineral metabolism? J. Appl. Ichthyol. 2010, 26, 210–213. [Google Scholar] [CrossRef]
- Pasqualetti, S.; Congiu, T.; Banfi, G.; Mariotti, M. Alendronate rescued osteoporotic phenotype in a model of glucocorticoid-induced osteoporosis in adult zebrafish scale. Int. J. Exp. Pathol. 2015, 96, 11–20. [Google Scholar] [CrossRef]
- Ek-Rylander, B.; Flores, M.; Wendel, M.; Heinegård, D.; Andersson, G. Dephosphorylation of osteopontin and bone sialoprotein by osteoclastic tartrate-resistant acid phosphatase. Modulation of osteoclast adhesion in vitro. J. Biol. Chem. 1994, 269, 14853–14856. [Google Scholar] [CrossRef]
- Orimo, H. The mechanism of mineralization and the role of alkaline phosphatase in health and disease. J. Nippon. Med. School 2010, 77, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Bossard, M.J.; Tomaszek, T.A.; Thompson, S.K.; Amegadzie, B.Y.; Hanning, C.R.; Jones, C.; Kurdyla, J.T.; McNulty, D.E.; Drake, F.H.; Gowen, M.; et al. Proteolytic activity of human osteoclast cathepsin K. Expression, purification, activation, and substrate identification. J. Biol. Chem. 1996, 271, 12517–12524. [Google Scholar] [CrossRef] [Green Version]
- Bull, H.; Murray, P.G.; Thomas, D.; Fraser, A.M.; Nelson, P.N. Acid phosphatases. Mol. Pathol. 2002, 55, 65–72. [Google Scholar] [CrossRef]
- Koncki, R.; Ogończyk, D.; Głąb, S. Potentiometric assay for acid and alkaline phosphatase. Anal. Chim. Acta 2005, 538, 257–261. [Google Scholar] [CrossRef]
- de Vrieze, E.; Zethof, J.; Schulte-Merker, S.; Flik, G.; Metz, J.R. Identification of novel osteogenic compounds by an ex-vivo sp7:luciferase zebrafish scale assay. Bone 2015, 74, 106–113. [Google Scholar] [CrossRef]
- Persson, P.; Björnsson, B.T.; Takagi, Y. Characterization of morphology and physiological actions of scale osteoclasts in the rainbow trout. J. Fish Biol. 1999, 54, 669–684. [Google Scholar] [CrossRef]
- Compston, J. Glucocorticoid-induced osteoporosis: An update. Endocrine 2018, 61, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Nakamura, S.; Chinen, N.; Shimazawa, M.; Hara, H. Effects of anti-osteoporosis drugs against dexamethasone-induced osteoporosis-like phenotype using a zebrafish scale-regeneration model. J. Pharmacol. Sci. 2020, 143, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Dorozhkin, S.V. Calcium orthophosphates. J. Mater. Sci. 2007, 42, 1061–1095. [Google Scholar] [CrossRef]
- Zoorob, R.J.; Cender, D. A different look at corticosteroids. Am. Fam. Physician 1998, 58, 443–450. [Google Scholar] [PubMed]
- Lindenstrøm; Buchmann. Dexamethasone treatment increases susceptibility of rainbow trout, Oncorhynchus mykiss (Walbaum), to infections with Gyrodactylus derjavini Mikailov. J. Fish Dis. 1998, 21, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Rosen, A.; Saffitz, J.E.; Asimaki, A.; Litovsky, S.H.; Mackey-Bojack, S.M.; Halushka, M.K. Expression of cathepsin K and tartrate-resistant acid phosphatase is not confined to osteoclasts but is a general feature of multinucleated giant cells: Systematic analysis. Rheumatology 2013, 52, 1529–1533. [Google Scholar] [CrossRef] [Green Version]
- Reithmeier, A.; Norgård, M.; Ek-Rylander, B.; Näreoja, T.; Andersson, G. Cathepsin K regulates localization and secretion of Tartrate-Resistant Acid Phosphatase (TRAP) in TRAP-overexpressing MDA-MB-231 breast cancer cells. BMC Mol. Cell Biol. 2020, 21, 15. [Google Scholar] [CrossRef]
- Aghajanian, P.; Mohan, S. The art of building bone: Emerging role of chondrocyte-to-osteoblast transdifferentiation in endochondral ossification. Bone Res. 2018, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Pullig, O.; Weseloh, G.; Ronneberger, D.L.; Käkönen, S.M.; Swoboda, B. Chondrocyte differentiation in human osteoarthritis: Expression of osteocalcin in normal and osteoarthritic cartilage and bone. Calcif. Tissue Int. 2000, 67, 230–240. [Google Scholar] [CrossRef]
- Delmas, P.D.; Eastell, R.; Garnero, P.; Seibel, M.J.; Stepan, J.; Committee of Scientific Advisors of the International Osteoporosis Foundation. The use of biochemical markers of bone turnover in osteoporosis. Osteoporos. Int. 2000, 11, S2–S17. [Google Scholar] [CrossRef]
- Stoch, S.A.; Wagner, J.A. Cathepsin K Inhibitors: A novel target for osteoporosis therapy. Clin. Pharmacol. Ther. 2008, 83, 172–176. [Google Scholar] [CrossRef]
- Brömme, D.; Lecaille, F. Cathepsin K inhibitors for osteoporosis and potential off-target effects. Expert Opin. Investig. Drugs 2009, 18, 585–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowen, M.; Lazner, F.; Dodds, R.; Kapadia, R.; Feild, J.; Tavaria, M.; Bertoncello, I.; Drake, F.; Zavarselk, S.; Tellis, I.; et al. Cathepsin K knockout mice develop osteopetrosis due to a deficit in matrix degradation but not demineralization. Journal of bone and mineral research. Off. J. Am. Soc. Bone Miner. Res. 1999, 14, 1654–1663. [Google Scholar] [CrossRef] [PubMed]
- Desmarais, S.; Black, W.C.; Oballa, R.; Lamontagne, S.; Riendeau, D.; Tawa, P.; Duong, L.T.; Pickarski, M.; Percival, M.D. Effect of cathepsin k inhibitor basicity on in vivo off-target activities. Mol. Pharmacol. 2008, 73, 147–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, S.; Logan, J.G.; Mellis, D.; Capulli, M. Generation and culture of osteoclasts. Bonekey Rep. 2014, 3, 570. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Kitamura, K.-I.; Omori, K.; Nemoto, T.; Satoh, Y.; Tabata, M.J.; Ikegame, M.; Yamamoto, T.; Ijiri, K.; Furusawa, Y. Response of osteoblasts and osteoclasts in regenerating scales to gravity loading. Biol. Sci. Space 2009, 23, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Golub, E.E.; Boesze-Battaglia, K. The role of alkaline phosphatase in mineralization. Curr. Opin. Orthop. 2007, 18, 444–448. [Google Scholar] [CrossRef]
- Gade, T.P.; Motley, M.W.; Beattie, B.J.; Bhakta, R.; Boskey, A.L.; Koutcher, J.A.; Mayer-Kuckuk, P. Imaging of alkaline phosphatase activity in bone tissue. PLoS ONE 2011, 6, e22608. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wandler, A.; Postlethwait, J.; Guillemin, K. Dynamic evolution of the LPS-detoxifying enzyme intestinal alkaline phosphatase in zebrafish and other vertebrates. Front. Immunol. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio Rerio), 4th ed.; University of Oregon Press: 5274 University of Oregon, Eugene, OR, USA, 2000. [Google Scholar]
- Suzuki, N.; Hayakawa, K.; Kameda, T.; Triba, A.; Tang, N.; Tabata, M.J.; Takada, K.; Wada, S.; Omori, K.; Srivastav, A.K.; et al. Monohydroxylated polycyclic aromatic hydrocarbons inhibit both osteoclastic and osteoblastic activities in teleost scales. Life Sci. 2009, 84, 482–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaichit, S.; Sato, T.; Yu, H.; Tanaka, Y.-k.; Ogra, Y.; Mizoguchi, T.; Itoh, M. Evaluation of Dexamethasone-Induced Osteoporosis In Vivo Using Zebrafish Scales. Pharmaceuticals 2021, 14, 536. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060536
Chaichit S, Sato T, Yu H, Tanaka Y-k, Ogra Y, Mizoguchi T, Itoh M. Evaluation of Dexamethasone-Induced Osteoporosis In Vivo Using Zebrafish Scales. Pharmaceuticals. 2021; 14(6):536. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060536
Chicago/Turabian StyleChaichit, Siripat, Takuto Sato, Huiqing Yu, Yu-ki Tanaka, Yasumitsu Ogra, Takamasa Mizoguchi, and Motoyuki Itoh. 2021. "Evaluation of Dexamethasone-Induced Osteoporosis In Vivo Using Zebrafish Scales" Pharmaceuticals 14, no. 6: 536. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060536