
Acrylonitrile Derivatives against Trypanosoma cruzi: In Vitro Activity and Programmed Cell Death Study

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Antiparasitic Activity
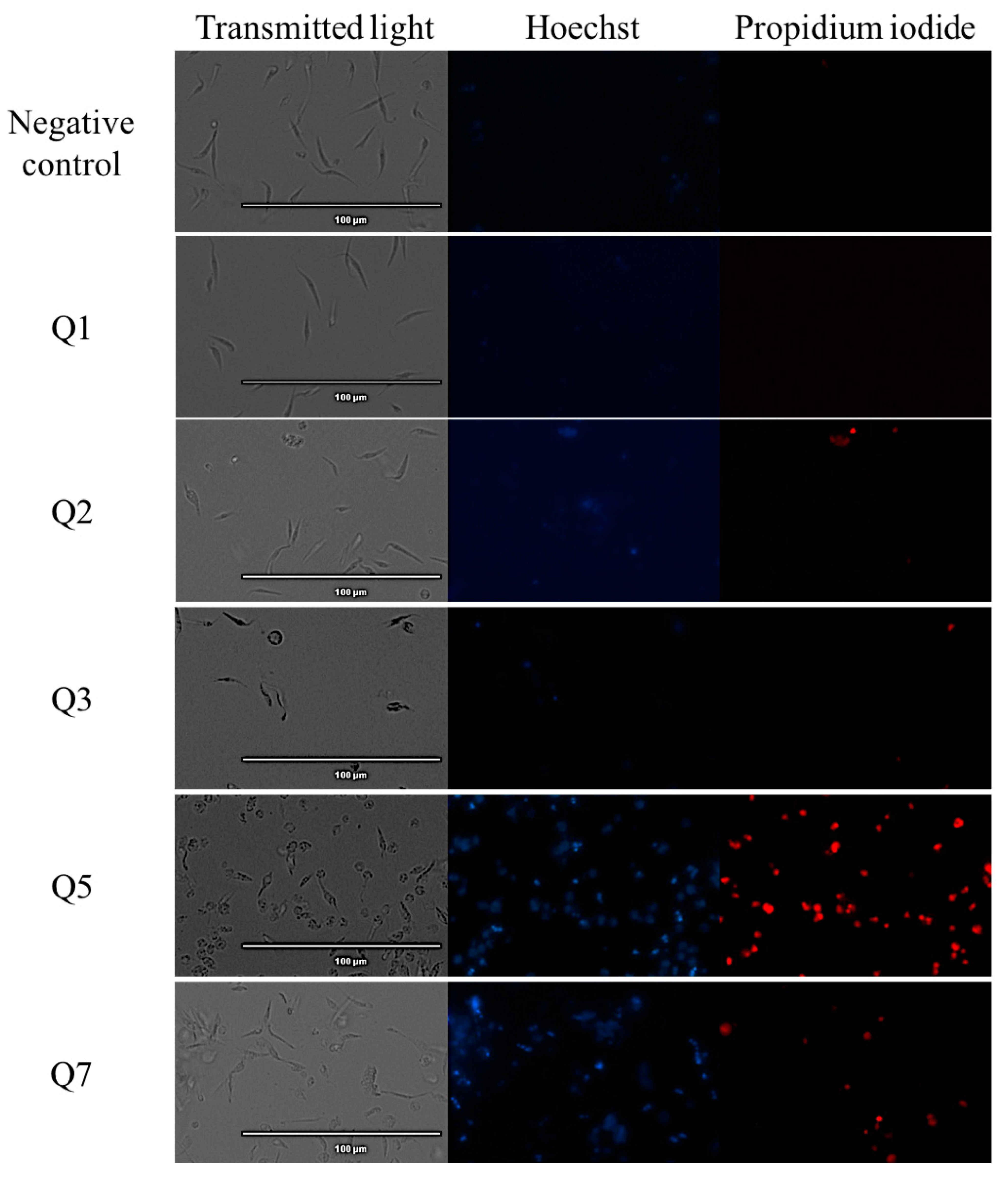
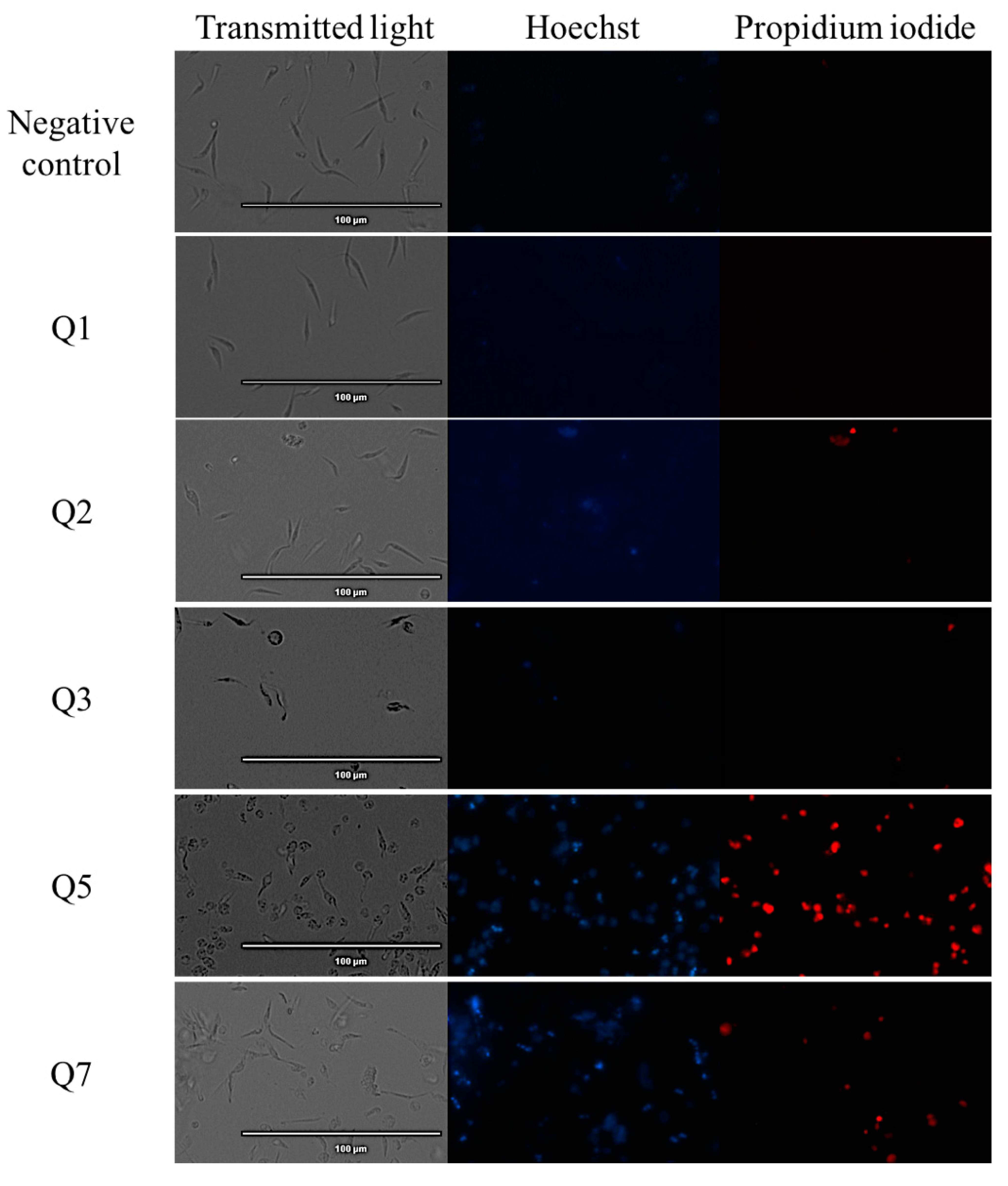
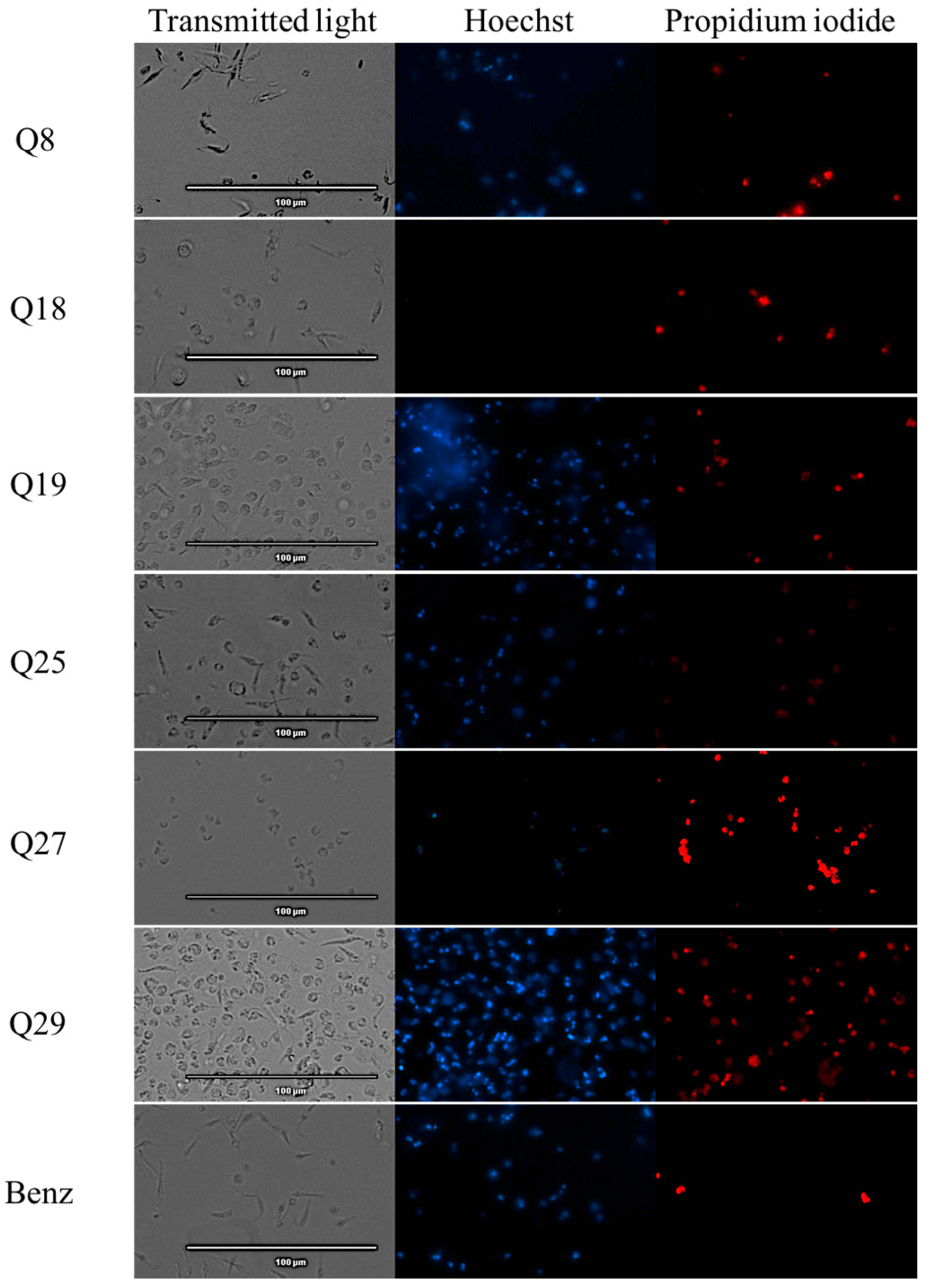
2.2. Chromatin Condensation Analysis
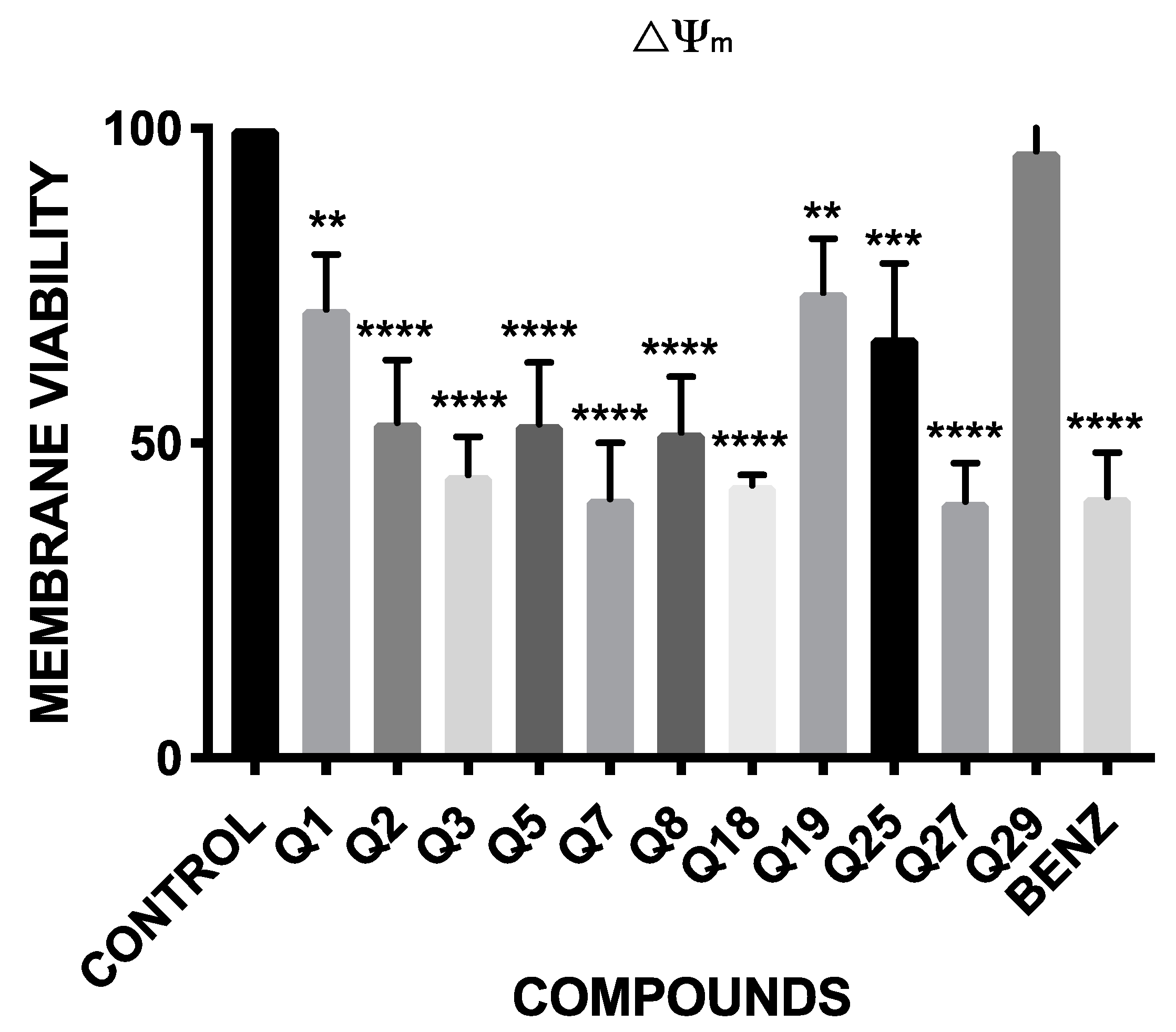
2.3. Mitochondrial Membrane Potential Analysis
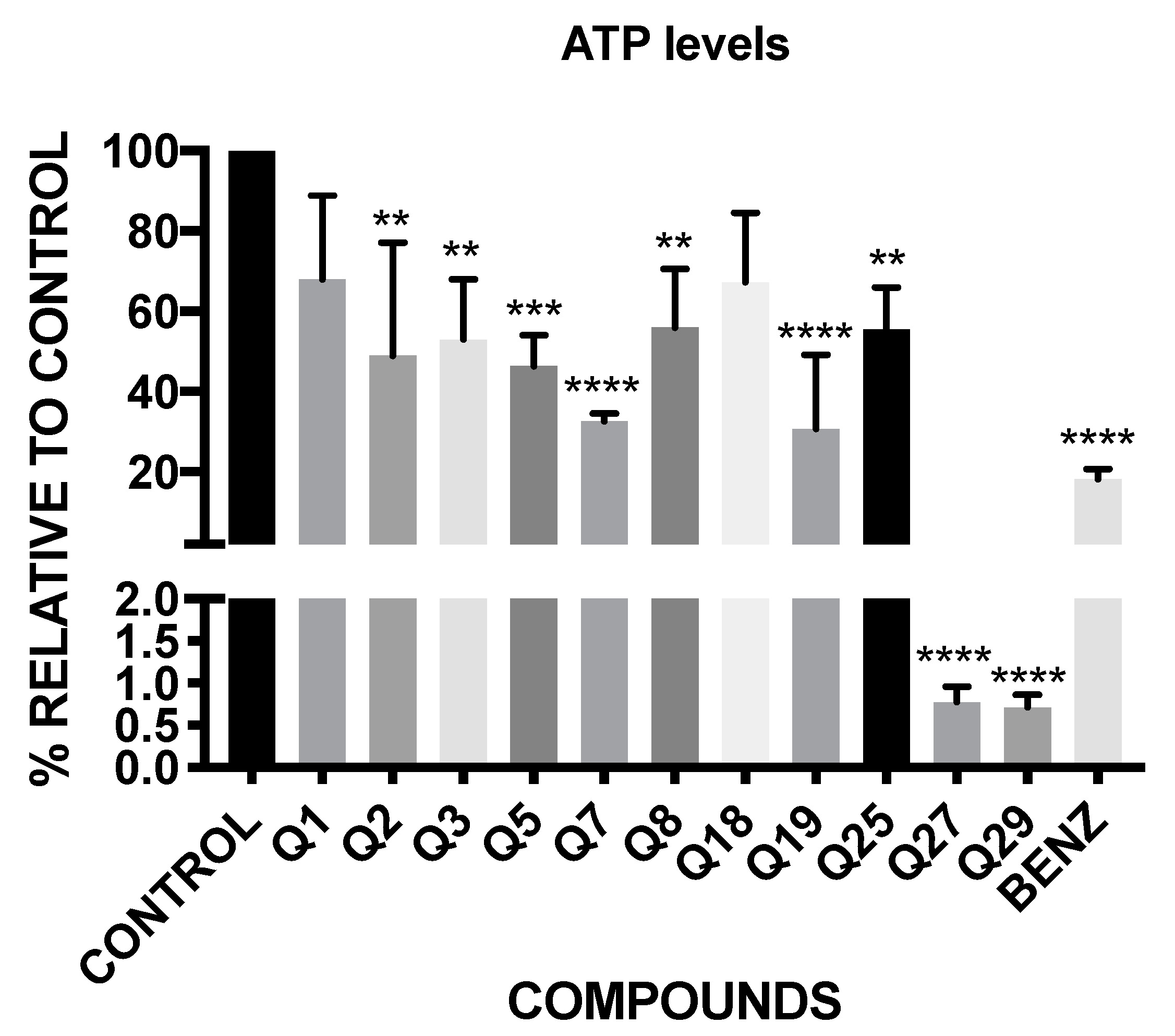
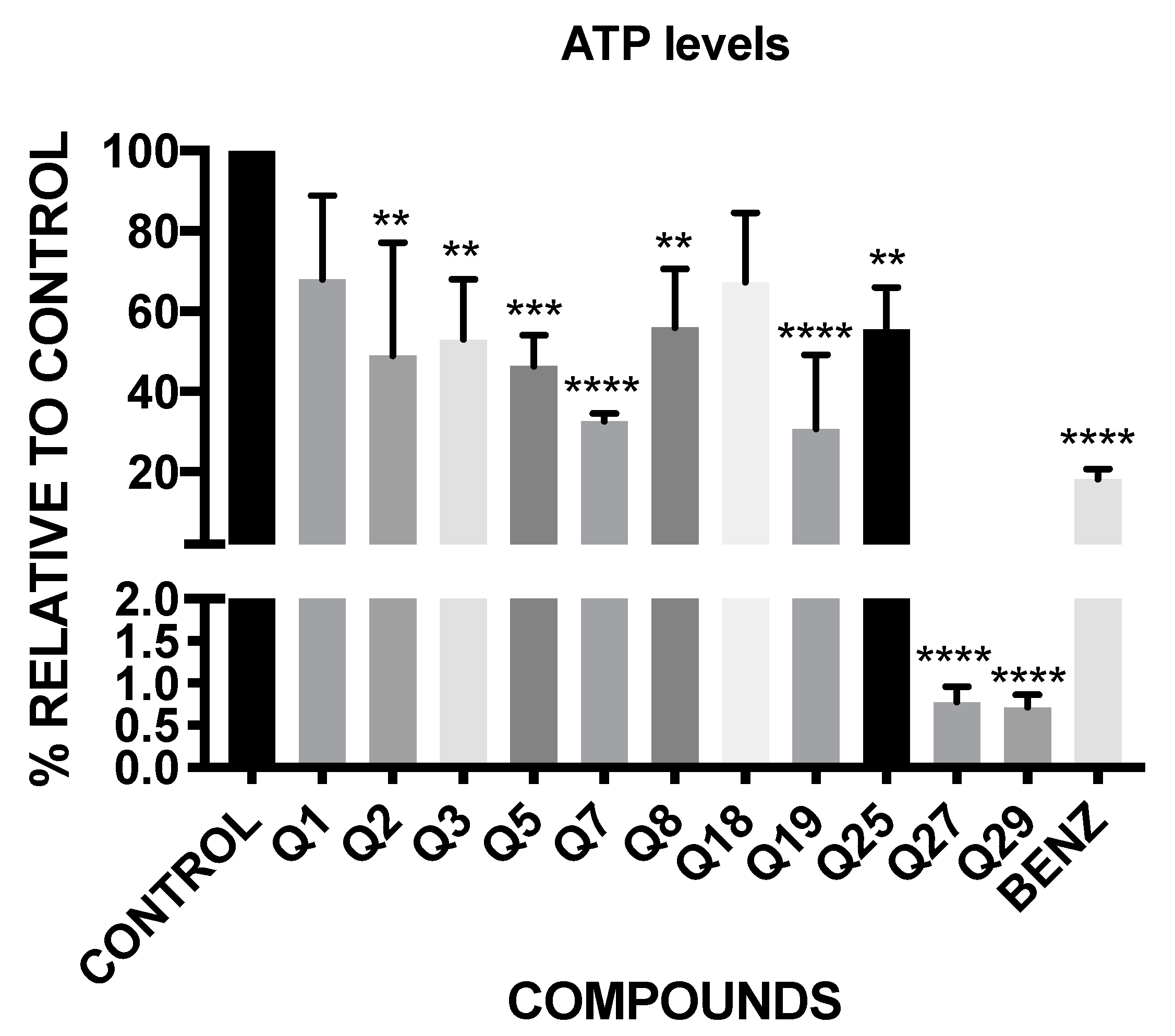
2.4. ATP Level Analysis
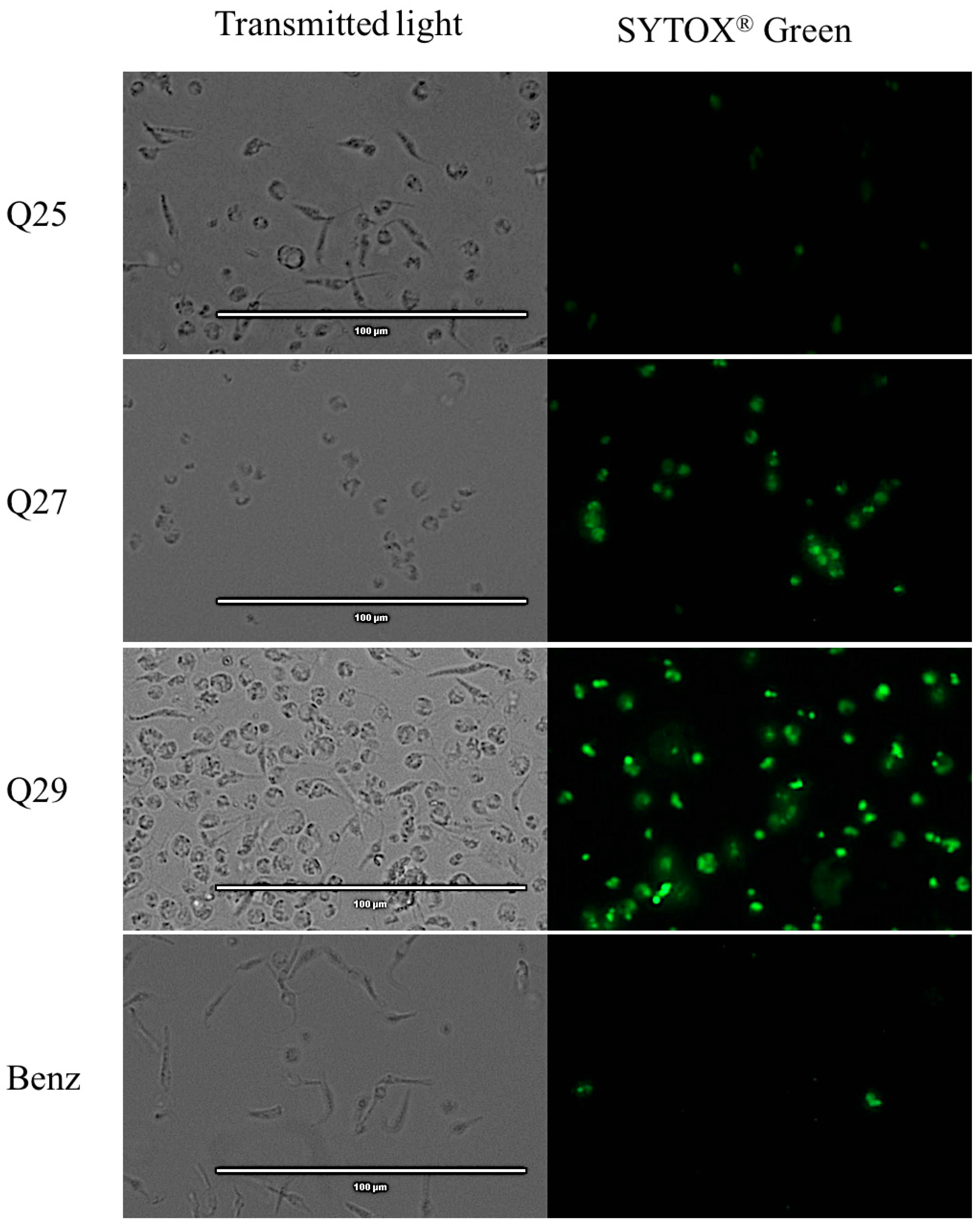
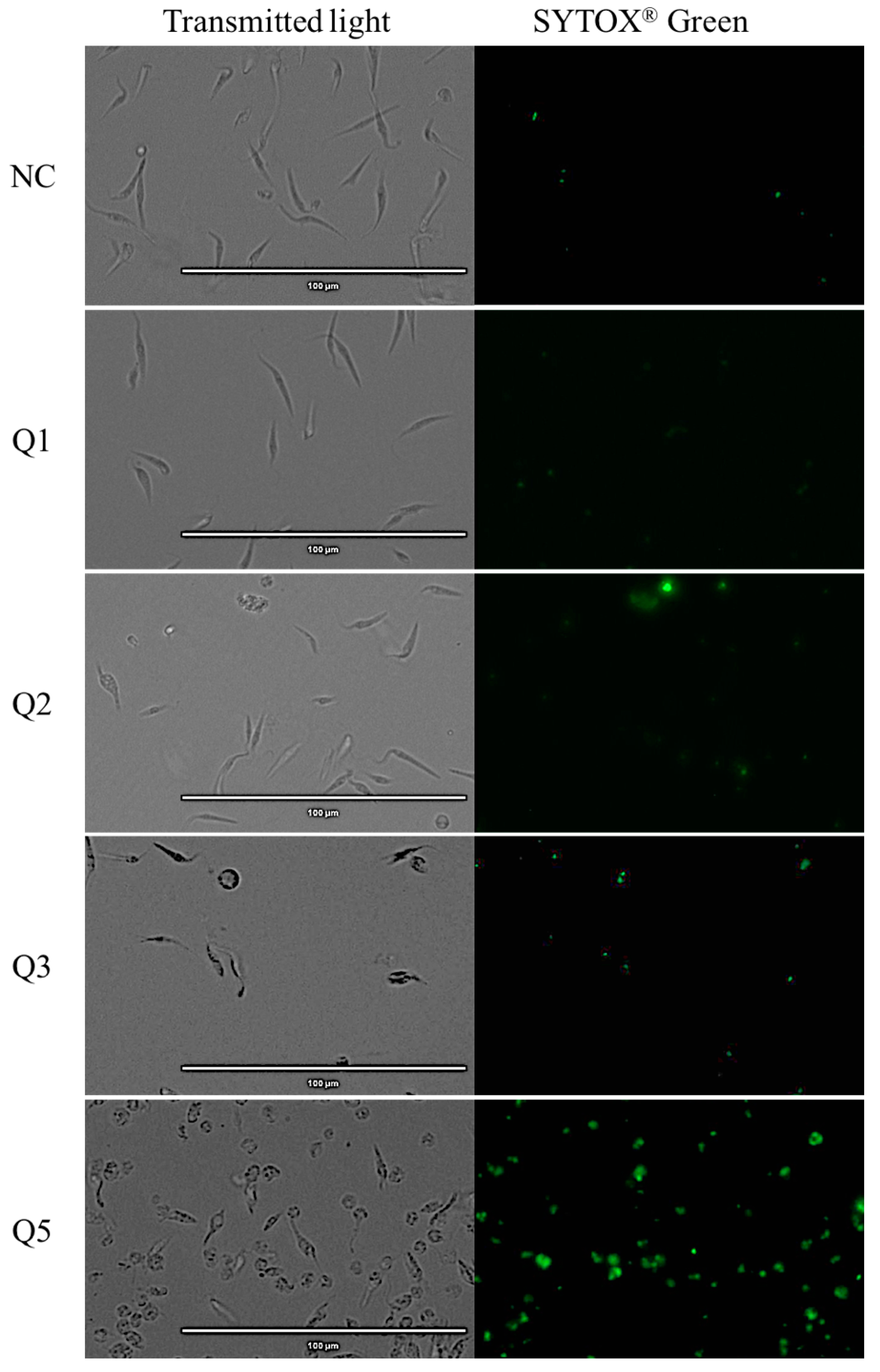
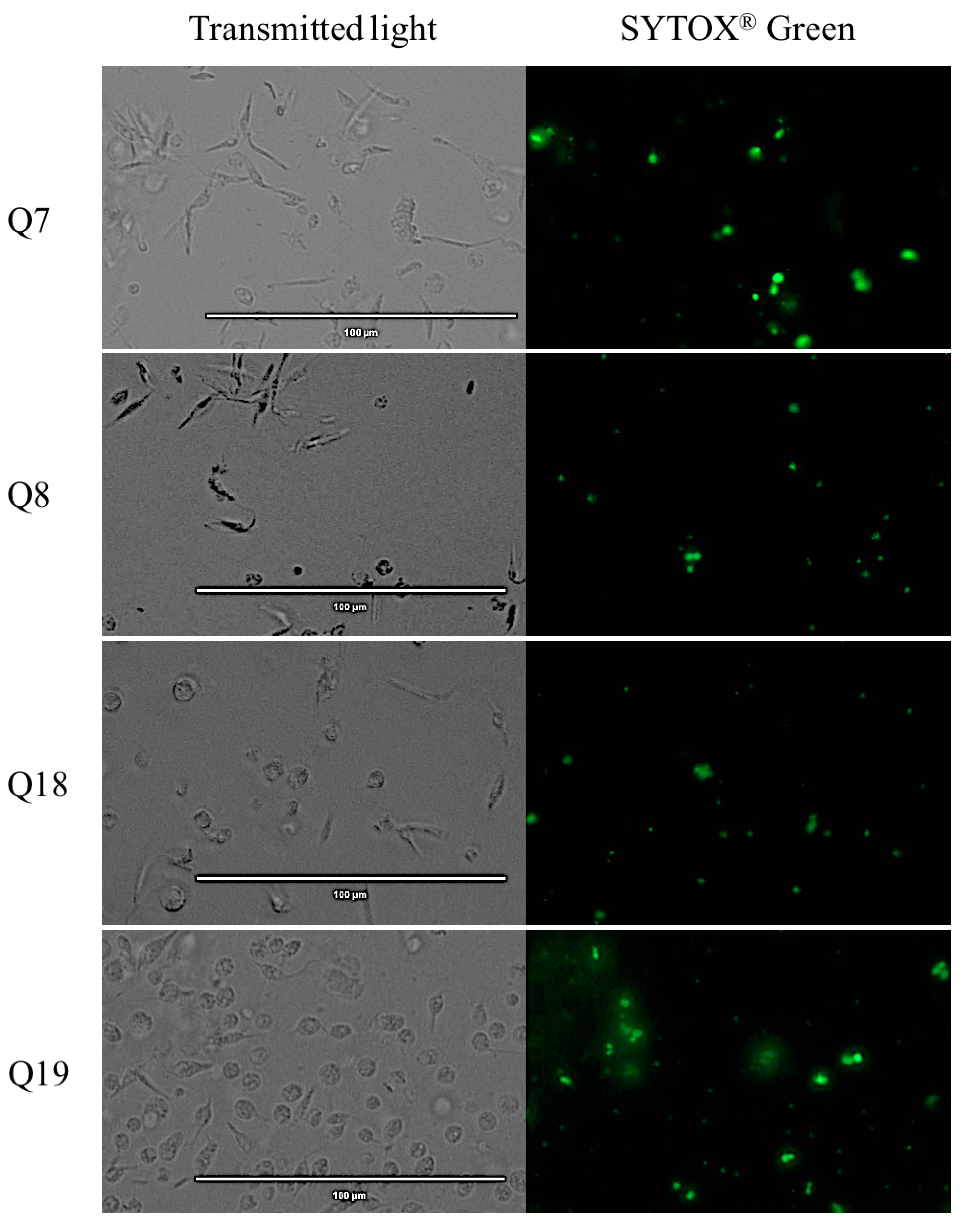
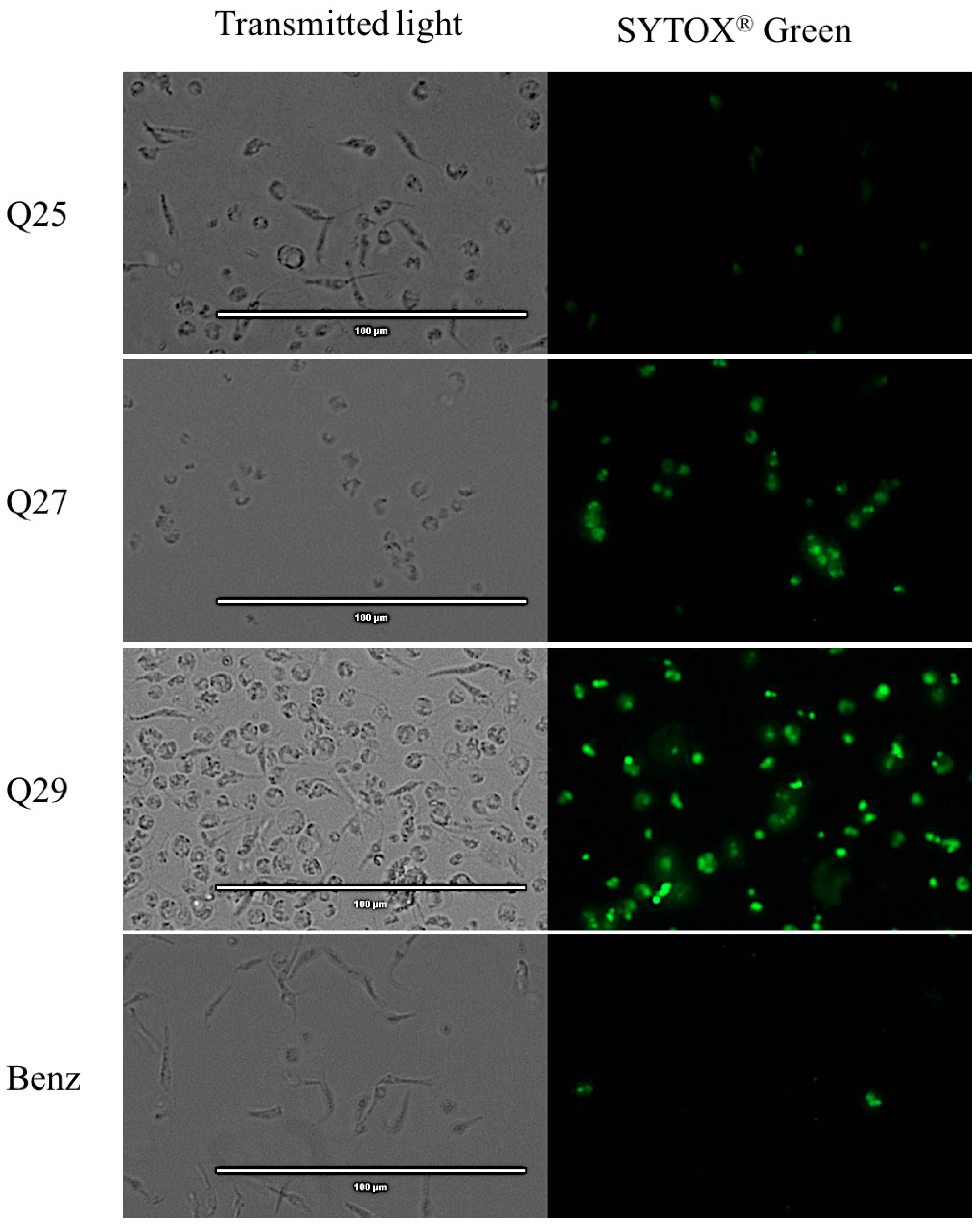
2.5. Plasmatic Membrane Permeability Analysis
2.6. Reactive Oxygen Species Analysis
3. Discussion
4. Materials and Methods
4.1. Compounds
4.2. Parasite Cultures
4.3. Antiparasitic Activity
4.4. Cytotoxicity Activity
4.5. Chromatin Condensation Analysis
4.6. Mitochondrial Membrane Potential Analysis
4.7. ATP Level Analysis
4.8. Plasmatic Membrane Permeability Analysis
4.9. Reactive Oxygen Species Analysis
4.10. Statistic Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Investing to Overcome the Global Impact of Neglected Tropical Diseases: Third WHO Report on Neglected Diseases 2015; Invest. to Overcome Glob. Impact Neglected Trop. Dis. Third WHO Rep. Neglected Dis.; World Health Organization: Geneva, Switzerland, 2015; p. 191. [Google Scholar]
- WHO. Integrating Neglected Tropical Diseases into Global Health and Development: Fourth WHO Report on Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2017; ISBN 9789241550116. [Google Scholar]
- Pinazo, M.J.; Gascon, J. The importance of the multidisciplinary approach to deal with the new epidemiological scenario of Chagas disease (global health). Acta Trop. 2015, 151, 16–20. [Google Scholar] [CrossRef] [Green Version]
- Gascon, J.; Bern, C.; Pinazo, M.J. Chagas disease in Spain, the United States and other non-endemic countries. Acta Trop. 2010, 115, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Guarner, J. Chagas disease as example of a reemerging parasite. Semin. Diagn. Pathol. 2019, 36, 164–169. [Google Scholar] [CrossRef]
- Prata, A. Clinical and epidemiological aspects of Chagas disease. Lancet Infect. Dis. 2001, 1, 92–100. [Google Scholar] [CrossRef]
- Sales Junior, P.A.; Molina, I.; Fonseca Murta, S.M.; Sánchez-Montalvá, A.; Salvador, F.; Corrêa-Oliveira, R.; Carneiro, C.M. Experimental and Clinical Treatment of Chagas Disease: A Review. Am. J. Trop. Med. Hyg. 2017, 97, 1289–1303. [Google Scholar] [CrossRef]
- Rassi, A.; Rassi, A.; Marin-Neto, J.A. Chagas disease. Lancet 2010, 375, 1388–1402. [Google Scholar] [CrossRef]
- Meymandi, S.; Hernandez, S.; Park, S.; Sanchez, D.R.; Forsyth, C. Treatment of Chagas Disease in the United States. Curr. Treat. Options Infect. Dis. 2018, 10, 373–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Moura Ferraz, L.R.; Alves, A.É.; da Silva Nascimento, D.D.; e Amariz, I.A.; Ferreira, A.S.; Costa, S.P.; Rolim, L.A.; de Lima, Á.A.; Neto, P.J. Technological innovation strategies for the specific treatment of Chagas disease based on Benznidazole. Acta Trop. 2018, 185, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.A.; de Mecca, M.M.; Bartel, L.C. Toxic side effects of drugs used to treat Chagas’ disease (American trypanosomiasis). Hum. Exp. Toxicol. 2006, 25, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Molina, J.A.; Sojo-Dorado, J.; Norman, F.; Monge-Maillo, B.; Díaz-Menéndez, M.; Albajar-Viñas, P.; López-Vélez, R. Nifurtimox therapy for Chagas disease does not cause hypersensitivity reactions in patients with such previous adverse reactions during benznidazole treatment. Acta Trop. 2013, 127, 101–104. [Google Scholar] [CrossRef]
- Pérez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- De Castro, E.; Reus, T.L.; de Aguiar, A.M.; Ávila, A.R.; de Arruda Campos Brasil de Souza, T. Procaspase-activating compound-1 induces apoptosis in Trypanosoma cruzi. Apoptosis 2017, 22, 1564–1577. [Google Scholar] [CrossRef]
- Jimenez, V.; Paredes, R.; Sosa, M.A.; Galanti, N. Natural programmed cell death in T. cruzi epimastigotes maintained in axenic cultures. J. Cell. Biochem. 2008, 105, 688–698. [Google Scholar] [CrossRef] [PubMed]
- De Menezes, R.R.P.P.B.; Sampaio, T.L.; Lima, D.B.; Sousa, P.L.; de Azevedo, I.E.P.; Magalhães, E.P.; Tessarolo, L.D.; Marinho, M.M.; Dos Santos, R.P.; Martins, A.M.C. Antiparasitic effect of (-)-α-bisabolol against Trypanosoma cruzi Y strain forms. Diagn. Microbiol. Infect. Dis. 2019, 95, 114860. [Google Scholar] [CrossRef] [PubMed]
- De Castro Andreassa, E.; Dos Santos, M.D.; Wassmandorf, R.; Wippel, H.H.; Carvalho, P.C.; da Gama Fischer, J.D. Proteomic changes in Trypanosoma cruzi epimastigotes treated with the proapoptotic compound PAC-1. Biochim. Biophys. Acta Proteins Proteom. 2021, 1869, 140582. [Google Scholar] [CrossRef] [PubMed]
- Bahia, M.T.; de Andrade, I.M.; Martins, T.A.F.; da Silva do Nascimento, Á.F.; de Figueiredo Diniz, L.; Caldas, I.S.; Talvani, A.; Trunz, B.B.; Torreele, E.; Ribeiro, I. Fexinidazole: A potential new drug candidate for Chagas disease. PLoS Negl. Trop. Dis. 2012, 6, e1870. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, V.; Dias, N.; Paiva, T.; Hagström-Bex, L.; Nitz, N.; Pratesi, R.; Hecht, M. Current trends in the pharmacological management of Chagas disease. Int. J. Parasitol. Drugs Drug Resist. 2020, 12, 7–17. [Google Scholar] [CrossRef]
- Apt, W.; Arribada, A.; Zulantay, I.; Rodríguez, J.; Saavedra, M.; Muñoz, A. Treatment of Chagas’ disease with itraconazole: Electrocardiographic and parasitological conditions after 20 years of follow-up. J. Antimicrob. Chemother. 2013, 68, 2164–2169. [Google Scholar] [CrossRef] [Green Version]
- Molina, I.; Gómez i Prat, J.; Salvador, F.; Treviño, B.; Sulleiro, E.; Serre, N.; Pou, D.; Roure, S.; Cabezos, J.; Valerio, L.; et al. Randomized trial of posaconazole and benznidazole for chronic Chagas’ disease. N. Engl. J. Med. 2014, 370, 1899–1908. [Google Scholar] [CrossRef]
- Vilar-Pereira, G.; Carneiro, V.C.; Mata-Santos, H.; Vicentino, A.R.R.; Ramos, I.P.; Giarola, N.L.L.; Feijó, D.F.; Meyer-Fernandes, J.R.; Paula-Neto, H.A.; Medei, E.; et al. Resveratrol Reverses Functional Chagas Heart Disease in Mice. PLoS Pathog. 2016, 12, e1005947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, D.D.; Mesquita, J.T.; da Costa Silva, T.A.; Romanelli, M.M.; da Gama Jaen Batista, D.; da Silva, C.F.; da Gama, A.N.S.; Neves, B.J.; Melo-Filho, C.C.; de Nazare Correia Soeiro, M.; et al. Efficacy of sertraline against Trypanosoma cruzi: An in vitro and in silico study. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 30. [Google Scholar] [CrossRef] [PubMed]
- Simões-Silva, M.R.; De Araújo, J.S.; Peres, R.B.; Da Silva, P.B.; Batista, M.M.; De Azevedo, L.D.; Bastos, M.M.; Bahia, M.T.; Boechat, N.; Soeiro, M.N.C. Repurposing strategies for Chagas disease therapy: The effect of imatinib and derivatives against Trypanosoma cruzi. Parasitology 2019, 146, 1006–1012. [Google Scholar] [CrossRef]
- AlNeyadi, S.S.; Salem, A.A.; Ghattas, M.A.; Atatreh, N.; Abdou, I.M. Antibacterial activity and mechanism of action of the benzazole acrylonitrile-based compounds: In vitro, spectroscopic, and docking studies. Eur. J. Med. Chem. 2017, 136, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Sirim, M.M.; Krishna, V.S.; Sriram, D.; Unsal Tan, O. Novel benzimidazole-acrylonitrile hybrids and their derivatives: Design, synthesis and antimycobacterial activity. Eur. J. Med. Chem. 2020, 188, 112010. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Cheng, Y.; Xu, M.; Song, Y.; Luo, Y.; Li, B. Synthesis, Acaricidal Activity, and Structure-Activity Relationships of Pyrazolyl Acrylonitrile Derivatives. J. Agric. Food Chem. 2016, 64, 9586–9591. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Shrivastava, A.; Mehra, R.N.; Deora, G.S.; Alam, M.M.; Zaman, M.S.; Akhter, M. Synthesis of novel benzimidazole acrylonitriles for inhibition of Plasmodium falciparum growth by dual target inhibition. Arch. Pharm. 2018, 351. [Google Scholar] [CrossRef]
- Li, J.-J.; Ma, J.; Xin, Y.-B.; Quan, Z.-S.; Tian, Y.-S. Synthesis and pharmacological evaluation of 2,3-diphenyl acrylonitriles-bearing halogen as selective anticancer agents. Chem. Biol. Drug Des. 2018, 92, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Tejedor, D.; Delgado-Hernández, S.; Colella, L.; García-Tellado, F. Catalytic Hydrocyanation of Activated Terminal Alkynes. Chemistry 2019, 25, 15046–15049. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, J.M.; Nunes, F.O.; Ceole, L.F.; Klimeck, T.D.F.; da Cruz, L.A.; Tófoli, D.; Borges, B.S.; Garcez, W.S.; Tozetti, I.A.; Medeiros, L.C.S.; et al. Synergistic effect and ultrastructural changes in Trypanosoma cruzi caused by isoobtusilactone A in short exposure of time. PLoS ONE 2021, 16, e0245882. [Google Scholar] [CrossRef]
- Herrera-Mayorga, V.; Lara-Ramírez, E.E.; Chacón-Vargas, K.F.; Aguirre-Alvarado, C.; Rodríguez-Páez, L.; Alcántara-Farfán, V.; Cordero-Martínez, J.; Nogueda-Torres, B.; Reyes-Espinosa, F.; Bocanegra-García, V.; et al. Structure-Based Virtual Screening and In Vitro Evaluation of New Trypanosoma cruzi Cruzain Inhibitors. Int. J. Mol. Sci. 2019, 20, 1742. [Google Scholar] [CrossRef] [Green Version]
- Gomes, J.C.; Cianni, L.; Ribeiro, J.; dos Reis Rocho, F.; da Costa Martins Silva, S.; Batista, P.H.J.; Moraes, C.B.; Franco, C.H.; Freitas-Junior, L.H.G.; Kenny, P.W.; et al. Synthesis and structure-activity relationship of nitrile-based cruzain inhibitors incorporating a trifluoroethylamine-based P2 amide replacement. Bioorg. Med. Chem. 2019, 27, 115083. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Miao, J.; Pan, L.; Zheng, L.; Wang, X.; Lin, Y.; Wu, J. Acute and chronic toxicity effects of acrylonitrile to the juvenile marine flounder Paralichthys olivaceus. Environ. Sci. Pollut. Res. Int. 2018, 25, 35301–35311. [Google Scholar] [CrossRef]
- Farcas, M.T.; McKinney, W.; Qi, C.; Mandler, K.W.; Battelli, L.; Friend, S.A.; Stefaniak, A.B.; Jackson, M.; Orandle, M.; Winn, A.; et al. Pulmonary and systemic toxicity in rats following inhalation exposure of 3-D printer emissions from acrylonitrile butadiene styrene (ABS) filament. Inhal. Toxicol. 2020, 32, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Dong, Y.; Lu, R.; Yang, B.; Wang, S.; Xing, G.; Jiang, Y. Susceptibility to the acute toxicity of acrylonitrile in streptozotocin-induced diabetic rats: Protective effect of phenethyl isothiocyanate, a phytochemical CYP2E1 inhibitor. Drug Chem. Toxicol. 2021, 44, 130–139. [Google Scholar] [CrossRef]
- Yu, B.; Changsheng, Y.; Wenjun, Z.; Ben, L.; Hai, Q.; Jing, M.; Guangwei, X.; Shuhua, W.; Fang, L.; Aschner, M.; et al. Differential protection of pre- versus post-treatment with curcumin, Trolox, and N-acetylcysteine against acrylonitrile-induced cytotoxicity in primary rat astrocytes. Neurotoxicology 2015, 51, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Bordat, A.; Soliman, N.; Ben Chraït, I.; Manerlax, K.; Yagoubi, N.; Boissenot, T.; Nicolas, J.; Tsapis, N. The crucial role of macromolecular engineering, drug encapsulation and dilution on the thermoresponsiveness of UCST diblock copolymer nanoparticles used for hyperthermia. Eur. J. Pharm. Biopharm. 2019, 142, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.P.; Salehi, B.; Sharifi-Rad, M.; Pezzani, R.; Kobarfard, F.; Sharifi-Rad, J.; Nigam, M. Programmed Cell Death, from a Cancer Perspective: An Overview. Mol. Diagn. Ther. 2018, 22, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Hernández, M.A.; de la Cruz-Ojeda, P.; López-Grueso, M.J.; Navarro-Villarán, E.; Requejo-Aguilar, R.; Castejón-Vega, B.; Negrete, M.; Gallego, P.; Vega-Ochoa, Á.; Victor, V.M.; et al. Integrated molecular signaling involving mitochondrial dysfunction and alteration of cell metabolism induced by tyrosine kinase inhibitors in cancer. Redox Biol. 2020, 36, 101510. [Google Scholar] [CrossRef]
- Mosquillo, M.F.; Smircich, P.; Lima, A.; Gehrke, S.A.; Scalese, G.; Machado, I.; Gambino, D.; Garat, B.; Pérez-Díaz, L. High Throughput Approaches to Unravel the Mechanism of Action of a New Vanadium-Based Compound against Trypanosoma cruzi. Bioinorg. Chem. Appl. 2020, 2020, 1634270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menna-Barreto, R.F.S. Cell death pathways in pathogenic trypanosomatids: Lessons of (over)kill. Cell Death Dis. 2019, 10, 93. [Google Scholar] [CrossRef]
- Cartuche, L.; Sifaoui, I.; López-Arencibia, A.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Lorenzo-Morales, J.; Piñero, J.E.; Díaz-Marrero, A.R.; Fernández, J.J. Antikinetoplastid Activity of Indolocarbazoles from Streptomyces sanyensis. Biomolecules 2020, 10, 657. [Google Scholar] [CrossRef] [PubMed]
- Sifaoui, I.; Reyes-Batlle, M.; López-Arencibia, A.; Chiboub, O.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Rodríguez Expósito, R.L.; Rizo-Liendo, A.; Piñero, J.E.; Lorenzo-Morales, J. Screening of the pathogen box for the identification of anti-Acanthamoeba agents. Exp. Parasitol. 2019, 201, 90–92. [Google Scholar] [CrossRef]
- Díaz-Marrero, A.R.; López-Arencibia, A.; Bethencout-Estrella, C.J.; Cen-Pacheco, F.; Sifaoui, I.; Hernández Creus, A.; Duque-Ramírez, M.C.; Souto, M.L.; Hernández Daranas, A.; Lorenzo-Morales, J.; et al. Antiprotozoal activities of marine polyether triterpenoids. Bioorg. Chem. 2019, 92, 103276. [Google Scholar] [CrossRef] [PubMed]
- López-Arencibia, A.; Reyes-Batlle, M.; Freijo, M.B.; Sifaoui, I.; Bethencourt-Estrella, C.J.; Rizo-Liendo, A.; Chiboub, O.; McNaughton-Smith, G.; Lorenzo-Morales, J.; Abad-Grillo, T.; et al. In vitro activity of 1H-phenalen-1-one derivatives against Leishmania spp. and evidence of programmed cell death. Parasit. Vectors 2019, 12, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Arencibia, A.; Martín-Navarro, C.; Sifaoui, I.; Reyes-Batlle, M.; Wagner, C.; Lorenzo-Morales, J.; Maciver, S.K.; Piñero, J.E. Perifosine Mechanisms of Action in Leishmania Species. Antimicrob. Agents Chemother. 2017, 61, e02127-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Arencibia, A.; San Nicolás-Hernández, D.; Bethencourt-Estrella, C.J.; Sifaoui, I.; Reyes-Batlle, M.; Rodríguez-Expósito, R.L.; Rizo-Liendo, A.; Lorenzo-Morales, J.; Bazzocchi, I.L.; Piñero, J.E.; et al. Withanolides from Withania aristata as Antikinetoplastid Agents through Induction of Programmed Cell Death. Pathogens 2019, 8, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeouk, I.; Sifaoui, I.; López-Arencibia, A.; Reyes-Batlle, M.; Bethencourt-Estrella, C.J.; Bazzocchi, I.L.; Bekhti, K.; Lorenzo-Morales, J.; Jiménez, I.A.; Piñero, J.E. Sesquiterpenoids and flavonoids from Inula viscosa induce programmed cell death in kinetoplastids. Biomed. Pharmacother. 2020, 130, 110518. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 | Compound | IC50 | Compound | IC50 |
|---|---|---|---|---|---|
| Q1 | 17.41 ± 4.75 | Q12 | 20.94 ± 4.61 | Q23 | NA |
| Q2 | 25.87 ± 2.78 | Q13 | 25.04 ± 0.62 | Q24 | NA |
| Q3 | 46.36 ± 5.73 | Q14 | 26.46 ± 2.83 | Q25 | 153.55 ± 19.79 |
| Q4 | 47.3 ± 10.62 | Q15 | 32.9 ± 0.83 | Q26 | NA |
| Q5 | 34.32 ± 1.67 | Q16 | 7.65 ± 1.51 | Q27 | 290.91 ± 46.99 |
| Q6 | 51.85 ± 7.69 | Q17 | 14.08 ± 2.53 | Q28 | NA |
| Q7 | 15.52 ± 1.27 | Q18 | 14.83 ± 3.06 | Q29 | 137.59 ± 14.01 |
| Q8 | 4.36 ± 0.26 | Q19 | 14.64 ± 3.44 | Q30 | NA |
| Q9 | NA | Q20 | 49.13 ± 1.12 | Q31 | 20.38 ± 0.13 |
| Q10 | 11.01 ± 1.38 | Q21 | 3.86 ± 0.67 | Q32 | 3.73 ± 0.41 |
| Q11 | 5.53 ± 1.36 | Q22 | NA | BENZ | 6.92 ± 0.77 |
| Compound | CC50 | Compound | CC50 | Compound | CC50 |
|---|---|---|---|---|---|
| Q1 | 87.4 ± 7.02 | Q12 | 7.64 ± 1.21 | Q23 | ND |
| Q2 | 112.06 ± 15.75 | Q13 | 8.52 ± 0.04 | Q24 | ND |
| Q3 | 86.65 ± 22.78 | Q14 | 8.39 ± 0.25 | Q25 | 374.5 ± 47.87 |
| Q4 | 43.25 ± 9.83 | Q15 | 6.66 ± 1.77 | Q26 | ND |
| Q5 | 53.85 ± 12.76 | Q16 | 4.35 ± 0.25 | Q27 | 480.44 ± 29.05 |
| Q6 | 28.76 ± 2.53 | Q17 | 5.24 ± 0.7 | Q28 | 210.28 ± 8.99 |
| Q7 | 46.45 ± 3.81 | Q18 | 34.37 ± 1 | Q29 | 327.87 ± 2.85 |
| Q8 | 52.86 ± 4.51 | Q19 | 22.16 ± 3.24 | Q30 | ND |
| Q9 | ND | Q20 | 50.55 ± 10.82 | Q31 | 10.37 ± 1.06 |
| Q10 | 10.12 ± 1.46 | Q21 | 3.19 ± 0.68 | Q32 | 2.89 ± 0.74 |
| Q11 | 5.22 ± 1.02 | Q22 | 3.43 ± 0.68 | BENZ | >1500 |
| Compound | SI | Compound | SI | Compound | SI |
|---|---|---|---|---|---|
| Q1 | 5 | Q12 | 0.36 | Q23 | ND |
| Q2 | 4.32 | Q13 | 0.34 | Q24 | ND |
| Q3 | 1.87 | Q14 | 0.28 | Q25 | 2.44 |
| Q4 | 0.91 | Q15 | 0.20 | Q26 | ND |
| Q5 | 1.57 | Q16 | 0.57 | Q27 | 1.65 |
| Q6 | 0.55 | Q17 | 0.37 | Q28 | ND |
| Q7 | 2.99 | Q18 | 2.31 | Q29 | 2.38 |
| Q8 | 12.2 | Q19 | 1.51 | Q30 | ND |
| Q9 | ND | Q20 | 1.03 | Q31 | 0.51 |
| Q10 | 0.92 | Q21 | 0.83 | Q32 | 0.78 |
| Q11 | 0.94 | Q22 | ND | BENZ | >222 |
| ID | Compound | Structure | ID | Compound | Structure |
|---|---|---|---|---|---|
| Q1 | (E)-Methyl 3-cyanoacrylate |  | Q2 | (Z)-Methyl 3-cyanoacrylate |  |
| Q3 | (E)-Ethyl 3-cyanoacrylate |  | Q4 | (Z)-Ethyl 3-cyanoacrylate |  |
| Q5 | (E)-Octyl 3-cyanoacrylate |  | Q6 | (Z)-Octyl 3-cyanoacrylate |  |
| Q7 | (E)-phenyl 3-cyanoacrylate |  | Q8 | (Z)-phenyl 3-cyanoacrylate |  |
| Q9 | (E)-tert-Butyl 3-cyanoacrylate |  | Q10 | (E)-4-Oxonon-2-enenitrile |  |
| Q11 | (E)-4-Oxo-4-phenylbut-2-enenitrile |  | Q12 | (Z)-4-Oxo-4-phenylbut-2-enenitrile |  |
| Q13 | (E)-4-Oxo-4-(thiophen-2-yl)but-2-enenitrile |  | Q14 | (Z)-4-Oxo-4-(thiophen-2-yl)but-2-enenitrile |  |
| Q15 | (E)-4-(Furan-2-yl)-4-oxobut-2-enenitrile |  | Q16 | (E)-4-Cyclohexyl-4-oxobut-2-enenitrile |  |
| Q17 | (E)-4-Oxo-6-phenylhex-2-enenitrile |  | Q18 | (E)-Diethyl 2-cyanovinylphosphonate |  |
| Q19 | (E)-3-(diphenylphosphoryl)acrylonitrile |  | Q20 | (Z)-3-(diphenylphosphoryl)acrylonitrile |  |
| Q21 | (E)-3-Tosylacrylonitrile |  | Q22 | (Z)-3-Tosylacrylonitrile |  |
| Q23 | (E)-3-Cyano-N,N-dimethylacrylamide |  | Q24 | (Z)-3-Cyano-N,N-dimethylacrylamide |  |
| Q25 | (E)-4-Oxo-4-(pyrrolidin-1-yl)but-2-enenitrile |  | Q26 | (Z)-4-Oxo-4-(pyrrolidin-1-yl)but-2-enenitrile |  |
| Q27 | (E)-3-Cyano-N-methoxy-N-methylacrylamide |  | Q28 | (Z)-3-Cyano-N-methoxy-N-methylacrylamide |  |
| Q29 | (E)-3-Cyano-N-methyl-N-phenylacrylamide |  | Q30 | (Z)-3-Cyano-N-methyl-N-phenylacrylamid |  |
| Q31 | (E)-(R,E)-4-Oxo-4-(2-oxo-4-phenyloxazolidin-3-yl)but-2-enenitrile |  | Q32 | (Z)-(R,Z)-4-Oxo-4-(2-oxo-4-phenyloxazolidin-3-yl)but-2-enenitrile |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bethencourt-Estrella, C.J.; Delgado-Hernández, S.; López-Arencibia, A.; San Nicolás-Hernández, D.; Sifaoui, I.; Tejedor, D.; García-Tellado, F.; Lorenzo-Morales, J.; Piñero, J.E. Acrylonitrile Derivatives against Trypanosoma cruzi: In Vitro Activity and Programmed Cell Death Study. Pharmaceuticals 2021, 14, 552. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060552
Bethencourt-Estrella CJ, Delgado-Hernández S, López-Arencibia A, San Nicolás-Hernández D, Sifaoui I, Tejedor D, García-Tellado F, Lorenzo-Morales J, Piñero JE. Acrylonitrile Derivatives against Trypanosoma cruzi: In Vitro Activity and Programmed Cell Death Study. Pharmaceuticals. 2021; 14(6):552. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060552
Chicago/Turabian StyleBethencourt-Estrella, Carlos J., Samuel Delgado-Hernández, Atteneri López-Arencibia, Desirée San Nicolás-Hernández, Ines Sifaoui, David Tejedor, Fernando García-Tellado, Jacob Lorenzo-Morales, and José E. Piñero. 2021. "Acrylonitrile Derivatives against Trypanosoma cruzi: In Vitro Activity and Programmed Cell Death Study" Pharmaceuticals 14, no. 6: 552. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060552