
High-Throughput Phenotypic Assay to Screen for Anthelmintic Activity on Haemonchus contortus

 , ,
, ,  , and
, and 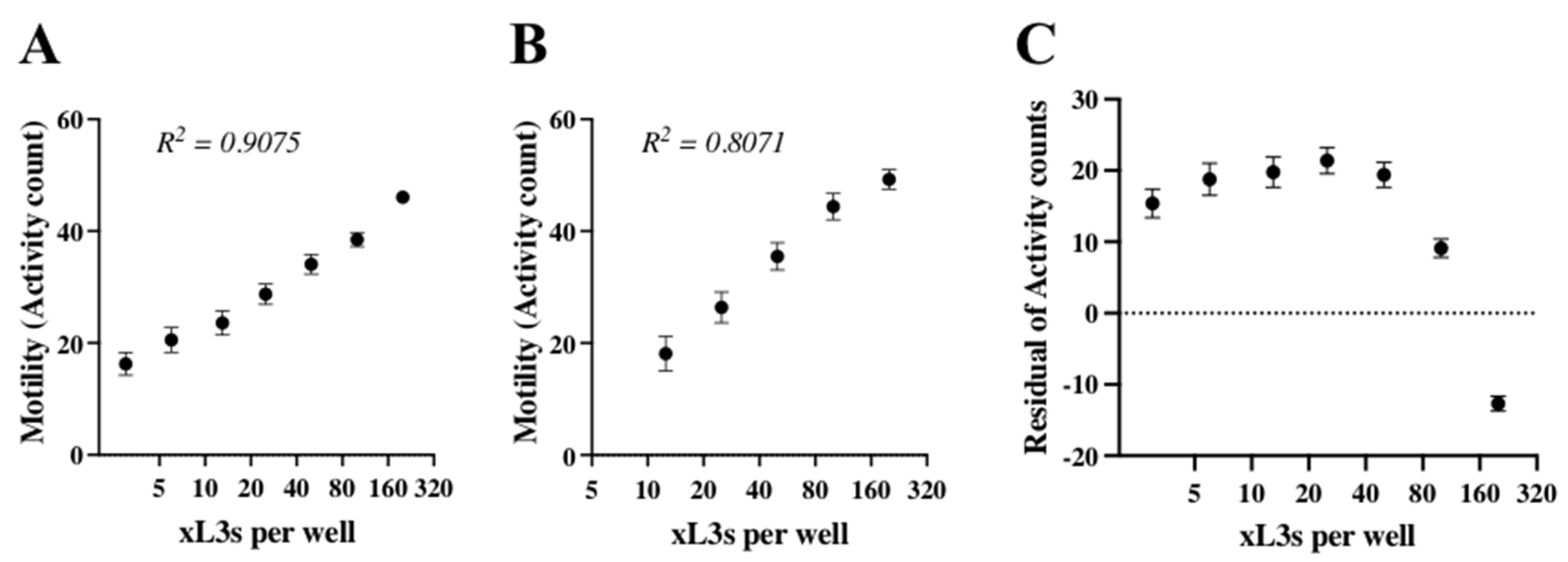{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Selection of the Acquisition Algorithm in WMicrotracker ONE Instrument (Phylumtech, Argentina) for the HTS Assay
2.2. Optimisation of xL3s Density, Assessment of Physical Stimulation and Time for the Measurement of Motility in the Assay
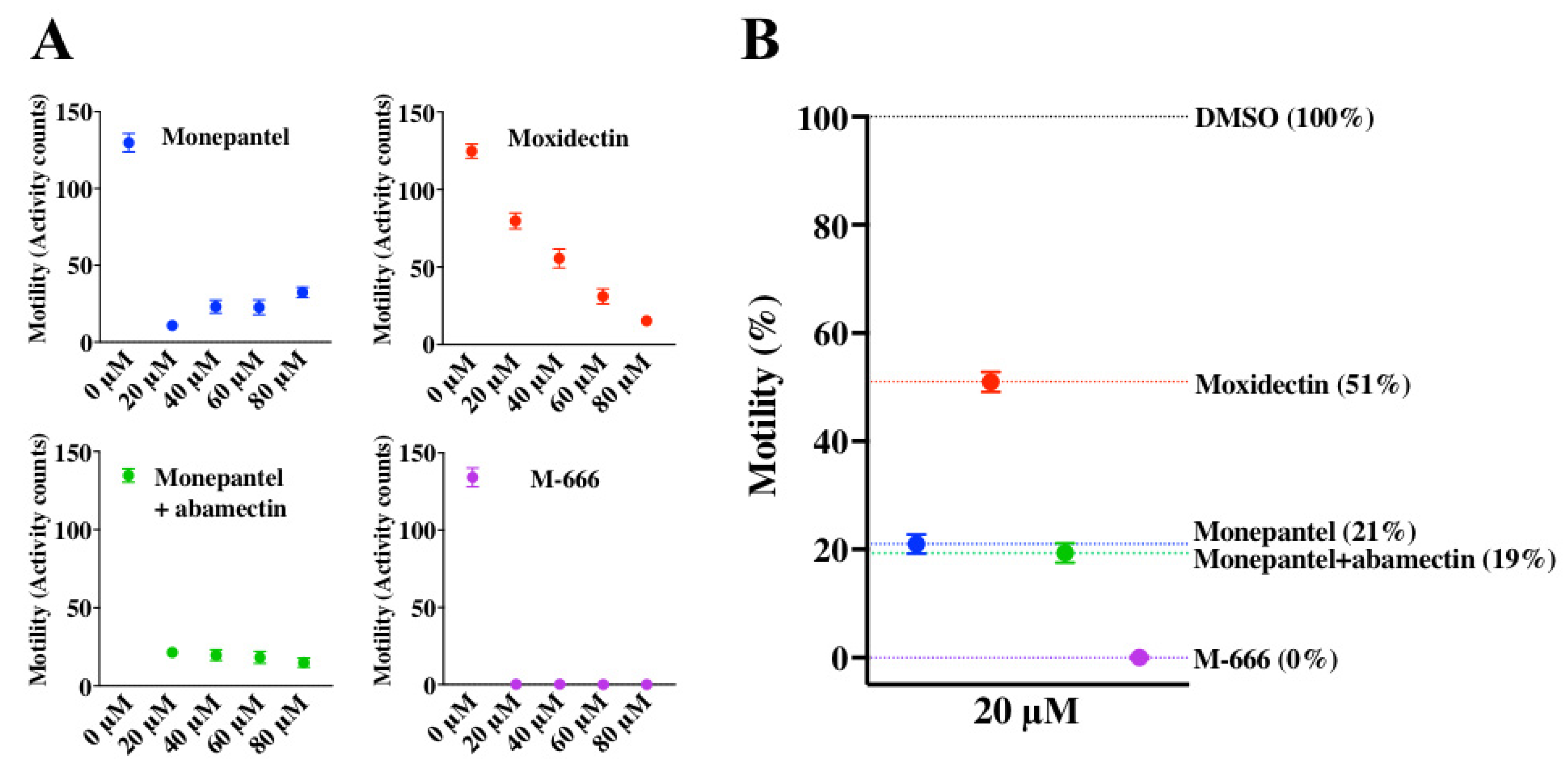
2.3. Establishing Compound Concentration, Hit-Identification Cut-off and IC50 Values for Reference Compounds
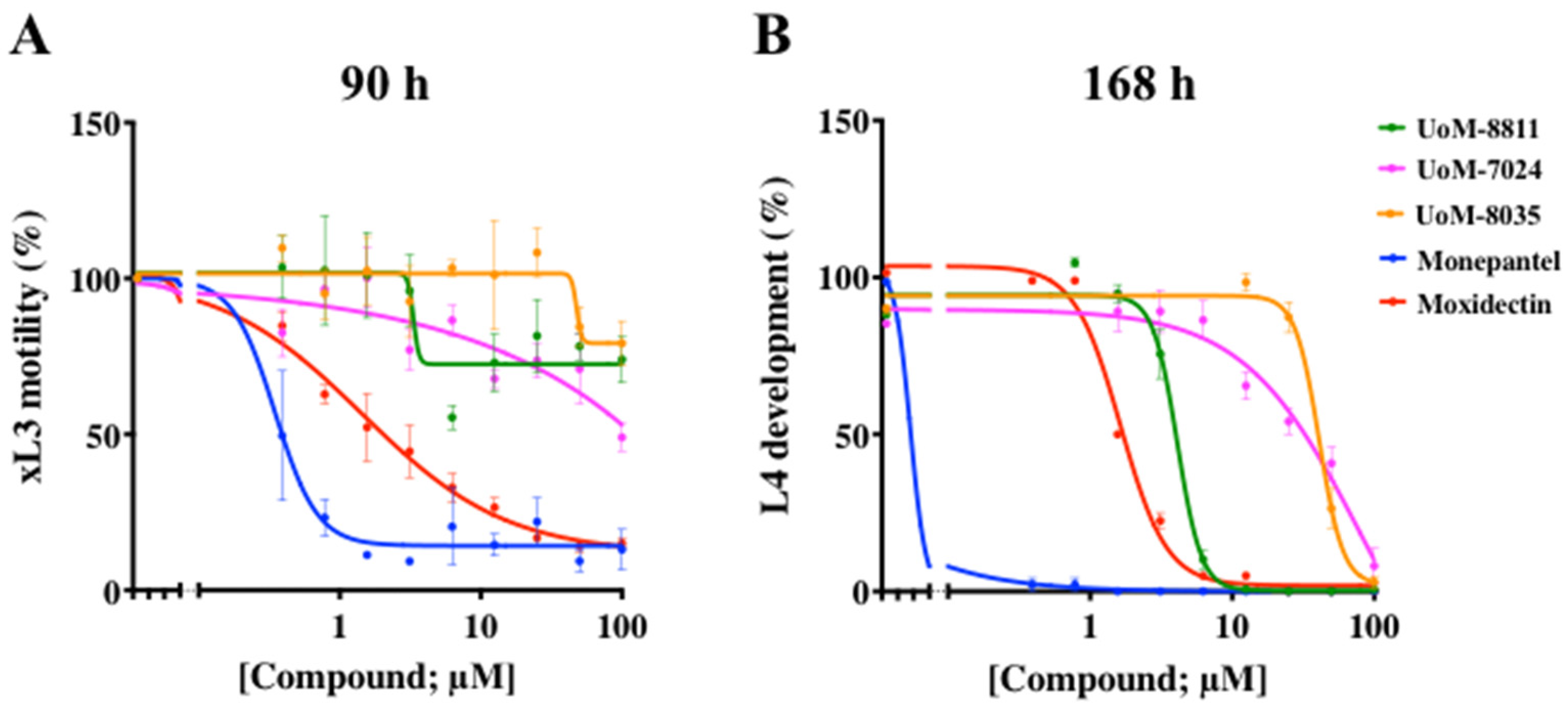
2.4. Screening Results, Dose Responses and Phenotypes Detected
3. Discussion
4. Materials and Methods
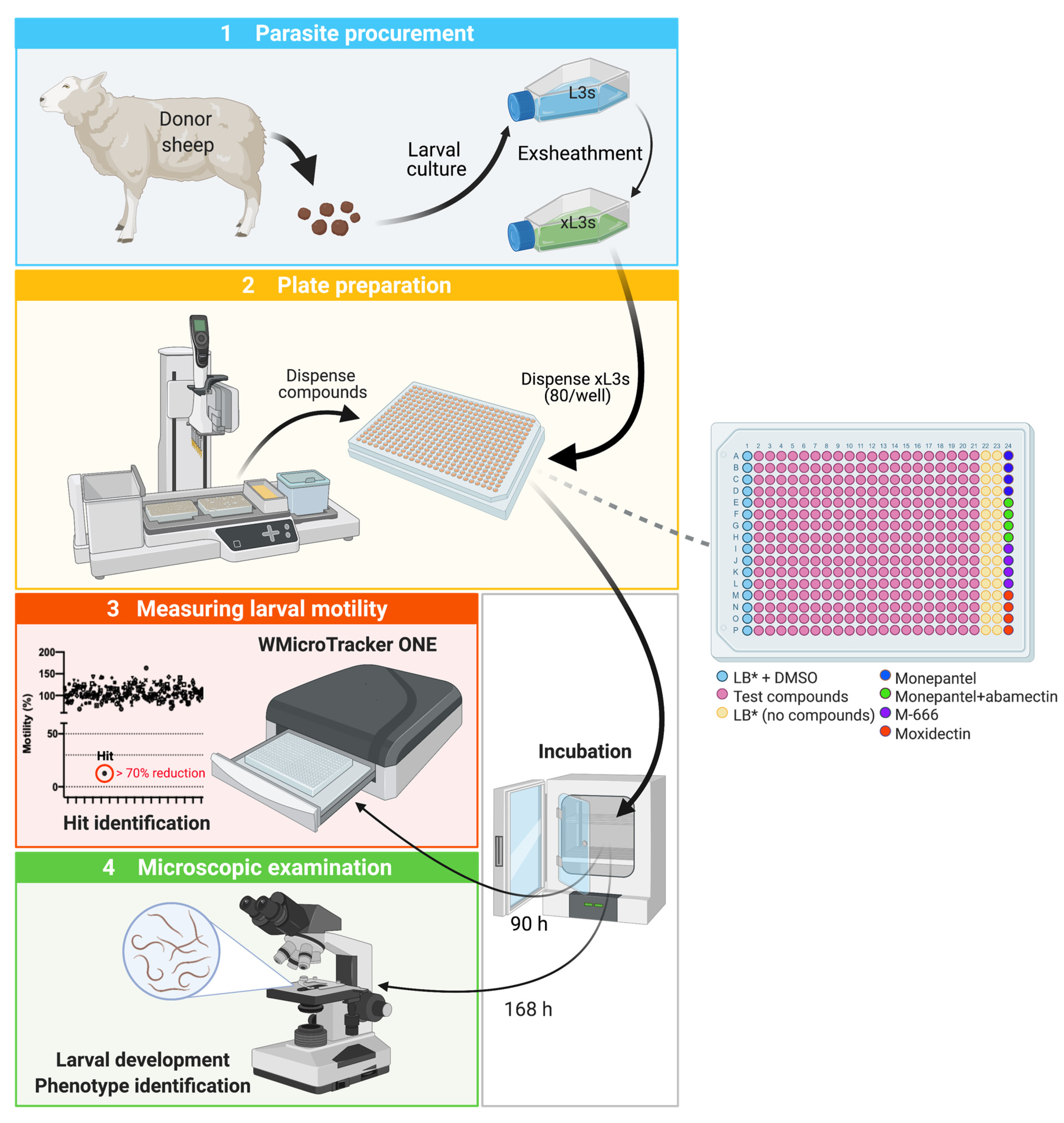
4.1. Procurement, Storage and Preparation of H. Contortus Larvae for Screening
4.2. High Throughput Screening of Compounds for xL3 Motility Reduction at 90 h
4.3. Assessing Larval Development and Phenotypes at 168 h
4.4. Dose-Response Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geary, T.G.; Sakanari, J.A.; Caffrey, C.R. Anthelmintic drug discovery: Into the future. J. Parasitol. 2015, 101, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Kotze, A.C.; Prichard, R.K. Anthelmintic resistance in Haemonchus contortus: History, mechanisms and diagnosis. Adv. Parasitol. 2016, 93, 397–428. [Google Scholar]
- Diemert, D.J.; Bottazzi, M.E.; Plieskatt, J.; Hotez, P.J.; Bethony, J.M. Lessons along the critical path: Developing vaccines against human helminths. Trends Parasitol. 2018, 34, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Stutzer, C.; Richards, S.A.; Ferreira, M.; Baron, S.; Maritz-Olivier, C. Metazoan parasite vaccines: Present status and future prospects. Front. Cell Infect. Microbiol. 2018, 8, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacks, D.L.; Peters, N.C.; Bethony, J.M. Vaccines against parasites. In The Vaccine Book, 2nd ed.; Bloom, B.R., Lambert, P.-H., Eds.; Academic Press: London, UK, 2016; pp. 331–360. ISBN 9780128054000. [Google Scholar]
- Matthews, J.B.; Geldhof, P.; Tzelos, T.; Claerebout, E. Progress in the development of subunit vaccines for gastrointestinal nematodes of ruminants. Parasite Immunol. 2016, 38, 744–753. [Google Scholar] [CrossRef] [Green Version]
- Nisbet, A.J.; Meeusen, E.N.; González, J.F.; Piedrafita, D.M. Immunity to Haemonchus contortus and vaccine development. Adv. Parasitol. 2016, 93, 353–396. [Google Scholar] [PubMed]
- Britton, C.; Emery, D.L.; McNeilly, T.N.; Nisbet, A.J.; Stear, M.J. The potential for vaccines against scour worms of small ruminants. Int. J. Parasitol. 2020, 50, 533–553. [Google Scholar] [CrossRef] [PubMed]
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; Van Wyk, J.A. Diagnosis, treatment and management of Haemonchus contortus in small ruminants. Adv. Parasitol. 2016, 93, 181–238. [Google Scholar] [PubMed]
- Geary, T.G. Haemonchus contortus: Applications in drug discovery. Adv. Parasitol. 2016, 93, 429–463. [Google Scholar]
- Jiao, Y.; Preston, S.; Hofmann, A.; Taki, A.C.; Baell, J.B.; Chang, B.C.H.; Jabbar, A.; Gasser, R.B. A perspective on the discovery of selected compounds with anthelmintic activity against the barber’s pole worm—Where to from here? Adv. Parasitol. 2020, 108, 1–45. [Google Scholar]
- SepúlvedaCrespo, D.; Reguera, R.M.; RojoVázquez, F.; Balaña-Fouce, R.; Martínez-Valladares, M. Drug discovery technologies: Caenorhabditis elegans as a model for anthelmintic therapeutics. Med. Res. Rev. 2020, 40, 1715–1753. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.R.; Luciani, G.M.; Musso, G.; Bagg, R.; Yeo, M.; Zhang, Y.; Rajendran, L.; Glavin, J.; Hunter, R.; Redman, E.; et al. Caenorhabditis elegans is a useful model for anthelmintic discovery. Nat. Commun. 2015, 6, 7485. [Google Scholar] [CrossRef]
- Partridge, F.A.; Brown, A.E.; Buckingham, S.D.; Willis, N.J.; Wynne, G.M.; Forman, R.; Else, K.J.; Morrison, A.A.; Matthews, J.B.; Russell, A.J.; et al. An automated high-throughput system for phenotypic screening of chemical libraries on C. elegans and parasitic nematodes. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 8–21. [Google Scholar] [CrossRef]
- Gasser, R.B.; von Samson-Himmelstjerna, G. Haemonchus contortus and haemonchosis—past, present and future trends. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2016; Volume 93, ISBN 9780128103951. [Google Scholar]
- Lanusse, C.E.; Alvarez, L.I.; Lifschitz, A.L. Gaining insights into the pharmacology of anthelmintics using Haemonchus contortus as a model nematode. Adv. Parasitol. 2016, 93, 465–518. [Google Scholar] [PubMed]
- Veglia, F. The Anatomy and Life-History of the Haemonchus contortus (Rud.). Rep. Dir. Vet. Res. 1915, 3–4, 347–500. [Google Scholar]
- Doyle, S.R.; Tracey, A.; Laing, R.; Holroyd, N.; Bartley, D.; Bazant, W.; Beasley, H.; Beech, R.; Britton, C.; Brooks, K.; et al. Extensive genomic and transcriptomic variation defines the chromosome-scale assembly of Haemonchus contortus, a model gastrointestinal worm. Commun. Biol. 2020, 3, 656. [Google Scholar] [CrossRef]
- Ma, G.; Wang, T.; Korhonen, P.K.; Hofmann, A.; Sternberg, P.W.; Young, N.D.; Gasser, R.B. Elucidating the molecular and developmental biology of parasitic nematodes: Moving to a multiomics paradigm. Adv. Parasitol. 2020, 108, 175–229. [Google Scholar] [PubMed]
- Preston, S.; Gasser, R.B. Working towards new drugs against parasitic worms in a public-development partnership. Trends Parasitol. 2018, 34, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Rohwer, A.; Lutz, J.; Chassaing, C.; Uphoff, M.; Heckeroth, A.R.; Selzer, P.M. Identification and profiling of nematicidal compounds in veterinary parasitology. In Parasitic Helminths: Targets, Screens, Drugs and Vaccines; Caffrey, C.R., Ed.; Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 2012; pp. 135–157. ISBN 9783527652969. [Google Scholar]
- Paveley, R.A.; Bickle, Q.D. Automated imaging and other developments in whole-organism anthelmintic screening. Parasite Immunol. 2013, 35, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.J.; Le Jambre, L.F. Larval paralysis as an in vitro assay of levamisole and morantel tartrate resistance in Ostertagia. Vet. Res. Commun. 1979, 3, 159–164. [Google Scholar] [CrossRef]
- Kotze, A.C.; Le Jambre, L.F.; O’Grady, J. A modified larval migration assay for detection of resistance to macrocyclic lactones in Haemonchus contortus, and drug screening with Trichostrongylidae parasites. Vet. Parasitol. 2006, 137, 294–305. [Google Scholar] [CrossRef]
- Demeler, J.; Küttler, U.; El-Abdellati, A.; Stafford, K.; Rydzik, A.; Varady, M.; Kenyon, F.; Coles, G.; Höglund, J.; Jackson, F.; et al. Standardization of the larval migration inhibition test for the detection of resistance to ivermectin in gastrointestinal nematodes of ruminants. Vet. Parasitol. 2010, 174, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Paveley, R.A.; Mansour, N.R.; Hallyburton, I.; Bleicher, L.S.; Benn, A.E.; Mikic, I.; Guidi, A.; Gilbert, I.H.; Hopkins, A.L.; Bickle, Q.D. Whole organism high-content screening by label-free, image-based Bayesian classification for parasitic diseases. PLoS Negl. Trop. Dis. 2012, 6, e1762. [Google Scholar] [CrossRef] [PubMed]
- Smout, M.J.; Kotze, A.C.; McCarthy, J.S.; Loukas, A. A novel high throughput assay for anthelmintic drug screening and resistance diagnosis by real-time monitoring of parasite motility. PLoS Negl. Trop. Dis. 2010, 4, e885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonetta, S.H.; Golombek, D.A. An automated tracking system for Caenorhabditis elegans locomotor behavior and circadian studies application. J. Neurosci. Methods 2007, 161, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Risi, G.; Aguilera, E.; Ladós, E.; Suárez, G.; Carrera, I.; Álvarez, G.; Salinas, G. Caenorhabditis elegans infrared-based motility assay identified new hits for nematicide drug development. Vet. Sci. 2019, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Preston, S.; Jabbar, A.; Nowell, C.; Joachim, A.; Ruttkowski, B.; Baell, J.; Cardno, T.; Korhonen, P.K.; Piedrafita, D.; Ansell, B.R.; et al. Low cost whole-organism screening of compounds for anthelmintic activity. Int. J. Parasitol. 2015, 45, 333–343. [Google Scholar] [CrossRef] [Green Version]
- Herath, H.M.P.D.; Taki, A.C.; Sleebs, B.E.; Hofmann, A.; Nguyen, N.; Preston, S.; Davis, R.A.; Jabbar, A.; Gasser, R.B. Advances in the discovery and development of anthelmintics by harnessing natural product scaffolds. Adv. Parasitol. 2021, 111, 203–251. [Google Scholar] [PubMed]
- Le, T.G.; Kundu, A.; Ghoshal, A.; Nguyen, N.H.; Preston, S.; Jiao, Y.; Ruan, B.; Xue, L.; Huang, F.; Keiser, J.; et al. Optimization of novel 1-methyl-1 h -pyrazole-5-carboxamides leads to high potency larval development inhibitors of the barber’s pole worm. J. Med. Chem. 2018, 61, 10875–10894. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.C.; Fisher, M.H.; Stapley, E.O.; Albers-Schonberg, G.; Jacob, A.T. Ivermectin: A potent new anthelmintic agent. Science 1983, 221, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, R.; Ducray, P.; Jung, M.; Clover, R.; Rufener, L.; Bouvier, J.; Weber, S.S.; Wenger, A.; Wieland-Berghausen, S.; Goebel, T.; et al. A new class of anthelmintics effective against drug-resistant nematodes. Nature 2008, 452, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Padhi, L.; Leyssen, P.; Liu, M.; Neyts, J.; Luyten, W. Antimicrobial, anthelmintic, and antiviral activity of plants traditionally used for treating infectious disease in the similipal biosphere reserve, Odisha, India. Front. Pharmacol. 2017, 8, 658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Landuyt, B.; Klaassen, H.; Geldhof, P.; Luyten, W. Screening of a drug repurposing library with a nematode motility assay identifies promising anthelmintic hits against Cooperia oncophora and other ruminant parasites. Vet. Parasitol. 2018, 265, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Kipanga, P.; Mai, A.H.; Dhondt, I.; Braeckman, B.P.; Borggraeve, W.D.; Luyten, W. Bioassay-guided isolation of three anthelmintic compounds from Warburgia ugandensis Sprague subspecies ugandensis, and the mechanism of action of polygodial. Int. J. Parasitol. 2018, 48, 833–844. [Google Scholar] [CrossRef]
- Puyvelde, L.V.; Liu, M.; Veryser, C.; Borggraeve, W.M.D.; Mungarulire, J.; Mukazayire, M.J.; Luyten, W. Active principles of Tetradenia riparia. IV. anthelmintic activity of 8(14),15-sandaracopimaradiene-7α,18-diol. J. Ethnopharmacol. 2018, 216, 229–232. [Google Scholar] [CrossRef]
- Bichara, D.; Calcaterra, N.B.; Arranz, S.; Armas, P.; Simonetta, S.H. Set-up of an infrared fast behavioral assay using zebrafish (Danio rerio) larvae, and its application in compound biotoxicity screening. J. Appl. Toxicol. 2014, 34, 214–219. [Google Scholar] [CrossRef]
- Clare, R.H.; Bardelle, C.; Harper, P.; Hong, W.D.; Börjesson, U.; Johnston, K.L.; Collier, M.; Myhill, L.; Cassidy, A.; Plant, D.; et al. Industrial scale high-throughput screening delivers multiple fast acting macrofilaricides. Nat. Commun. 2019, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, E.M.; Korhonen, P.K.; Campbell, B.E.; Young, N.D.; Jex, A.R.; Jabbar, A.; Hall, R.S.; Mondal, A.; Howe, A.C.; Pell, J.; et al. The genome and developmental transcriptome of the strongylid nematode Haemonchus contortus. Genome Biol. 2013, 14, R89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolaou, S.; Hartman, D.; Presidente, P.J.A.; Newton, S.E.; Gasser, R.B. HcSTK, a Caenorhabditis elegans PAR-1 homologue from the parasitic nematode, Haemonchus contortus. Int. J. Parasitol. 2002, 32, 749–758. [Google Scholar] [CrossRef]
- Zhang, J.H.; Chung, T.D.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Sommerville, R.I. The development of Haemonchus contortus to the fourth stage in vitro. J. Parasitol. 1966, 52, 127–136. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Ending the Neglect to Attain the Sustainable Development Goals—A Road Map for Neglected Tropical Diseases 2021–2030; WHO: Geneva, Switzerland, 2020; p. 55. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taki, A.C.; Byrne, J.J.; Wang, T.; Sleebs, B.E.; Nguyen, N.; Hall, R.S.; Korhonen, P.K.; Chang, B.C.H.; Jackson, P.; Jabbar, A.; et al. High-Throughput Phenotypic Assay to Screen for Anthelmintic Activity on Haemonchus contortus. Pharmaceuticals 2021, 14, 616. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14070616
Taki AC, Byrne JJ, Wang T, Sleebs BE, Nguyen N, Hall RS, Korhonen PK, Chang BCH, Jackson P, Jabbar A, et al. High-Throughput Phenotypic Assay to Screen for Anthelmintic Activity on Haemonchus contortus. Pharmaceuticals. 2021; 14(7):616. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14070616
Chicago/Turabian StyleTaki, Aya C., Joseph J. Byrne, Tao Wang, Brad E. Sleebs, Nghi Nguyen, Ross S. Hall, Pasi K. Korhonen, Bill C.H. Chang, Paul Jackson, Abdul Jabbar, and et al. 2021. "High-Throughput Phenotypic Assay to Screen for Anthelmintic Activity on Haemonchus contortus" Pharmaceuticals 14, no. 7: 616. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14070616