
Analysis of Boswellic Acid Contents and Related Pharmacological Activities of Frankincense-Based Remedies That Modulate Inflammation


Abstract
:1. Introduction
2. Results
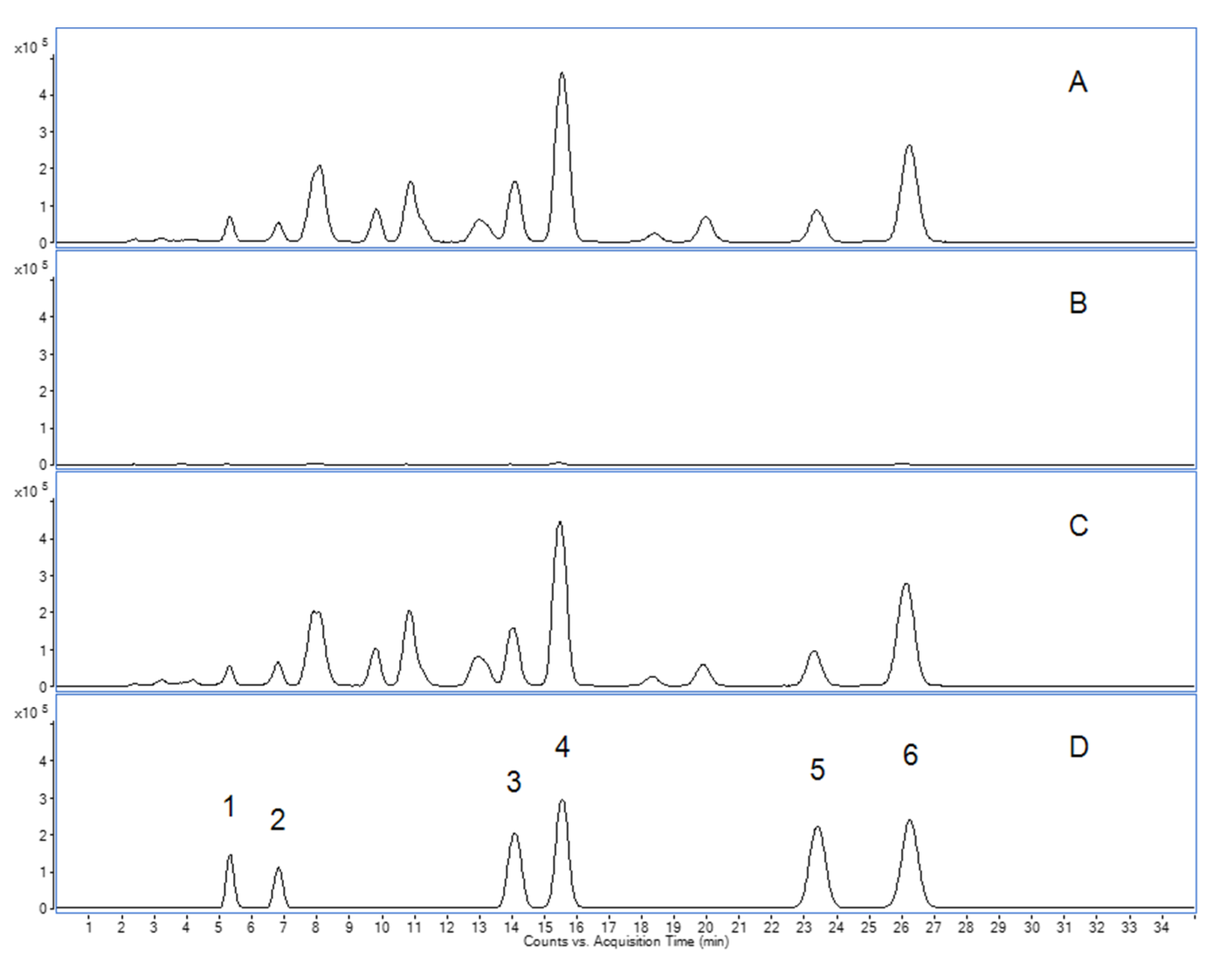
2.1. Analytical Assessment of Boswellic Acids in Frankincense-Based Remedies
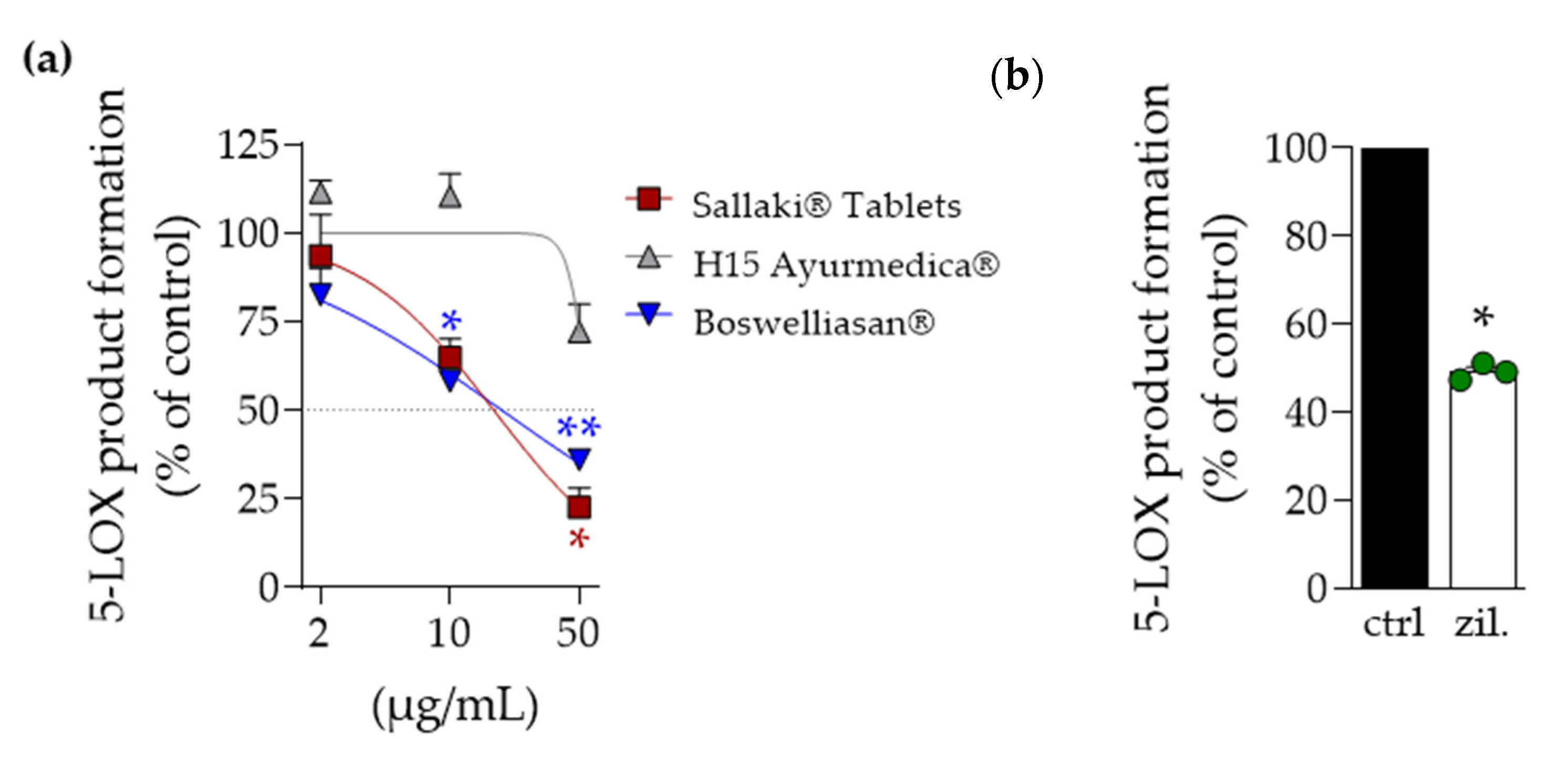
2.2. Inhibition of 5-Lipoxygenase and Microsomal Prostaglandin E2 Synthase-1 by Frankincense-Based Remedies
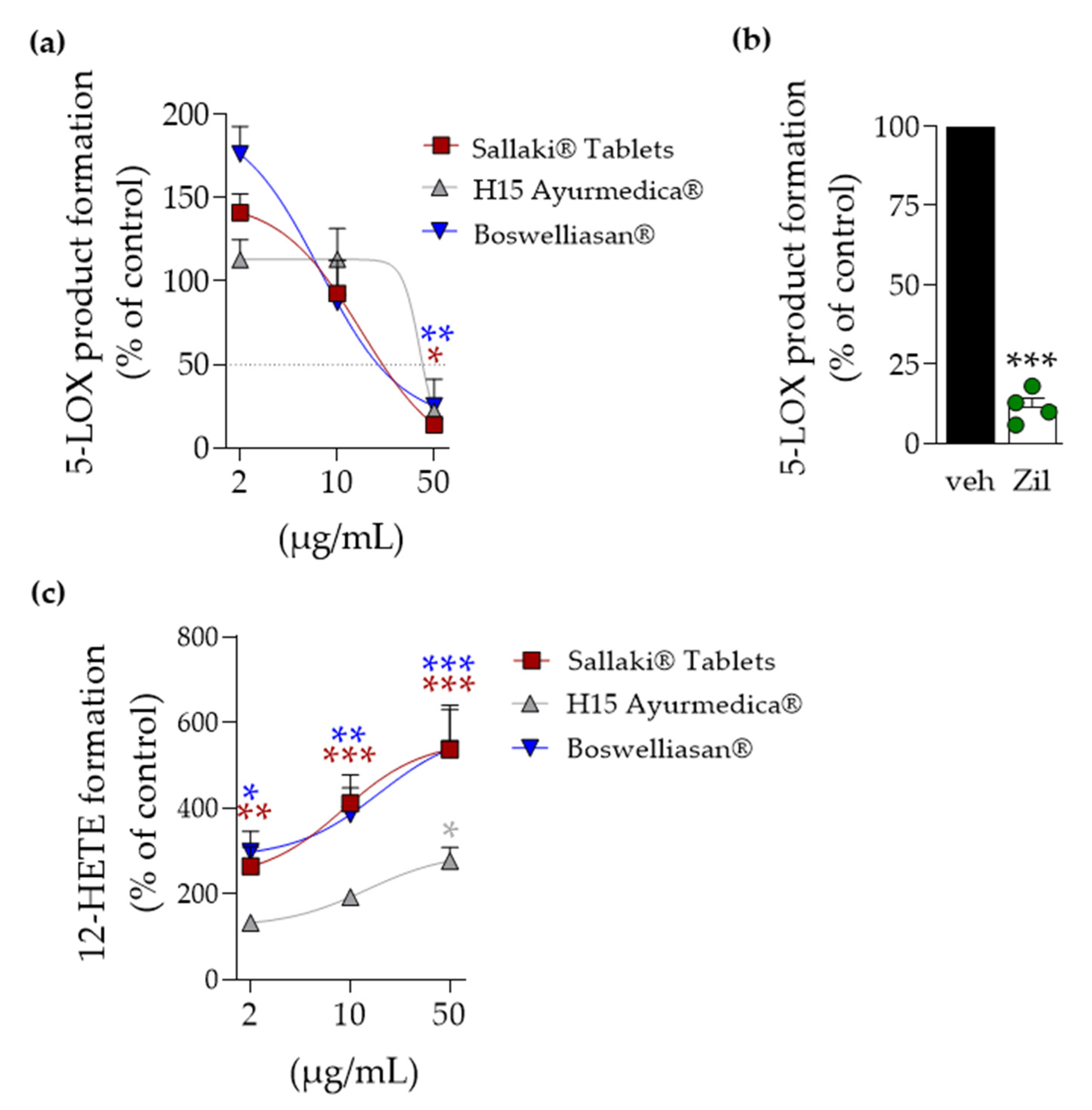
2.3. Modulation of Lipid Mediator Production in Exotoxin-Stimulated Neutrophils by Frankincense-Based Remedies
2.4. Effects of Frankincense-Based Remedies on Pro-Inflammatory Cytokine Secretion
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Standard Preparation
4.3. Sample Preparation
4.4. LC-MS/MS Method
4.5. Expression, Purification, and Activity Assay of Human Recombinant 5-LOX
4.6. Induction of mPGES-1 in A549 Cells, Isolation of Microsomes, and Determination of mPGES-1 Activity
4.7. Isolation of Human Leucocytes
4.8. Preparation of Exotoxin-Containing Staphylococcus Aureus-Conditioned Medium (SACM)
4.9. Analysis of LOX Product Formation in Human Neutrophils
4.10. Determination of Cytokine Levels
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Yasiry, A.R.; Kiczorowska, B. Frankincense--therapeutic properties. Postepy Hig. Med. Dosw. 2016, 70, 380–391. [Google Scholar] [CrossRef]
- Ernst, E. Frankincense: Systematic review. BMJ 2008, 337, a2813. [Google Scholar] [CrossRef] [Green Version]
- Moussaieff, A.; Mechoulam, R. Boswellia resin: From religious ceremonies to medical uses, a review of in-vitro, in-vivo and clinical trials. J. Pharm Pharmacol 2009, 61, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Tawab, M.; Werz, O.; Schubert-Zsilavecz, M. Boswellia serrata: An overall assessment of in vitro, preclinical, pharmacokinetic and clinical data. Clin. Pharmacokinet 2011, 50, 349–369. [Google Scholar] [CrossRef]
- Ammon, H.P. Boswellic acids in chronic inflammatory diseases. Planta Med. 2006, 72, 1100–1116. [Google Scholar] [CrossRef] [Green Version]
- Efferth, T.; Oesch, F. Anti-inflammatory and anti-cancer activities of frankincense: Targets, treatments and toxicities. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Kunnumakkara, A.B.; Banik, K.; Bordoloi, D.; Harsha, C.; Sailo, B.L.; Padmavathi, G.; Roy, N.K.; Gupta, S.C.; Aggarwal, B.B. Googling the Guggul (Commiphora and Boswellia) for Prevention of Chronic Diseases. Front. Pharmacol 2018, 9, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, N.K.; Parama, D.; Banik, K.; Bordoloi, D.; Devi, A.K.; Thakur, K.K.; Padmavathi, G.; Shakibaei, M.; Fan, L.; Sethi, G.; et al. An Update on Pharmacological Potential of Boswellic Acids against Chronic Diseases. Int J. Mol. Sci 2019, 20, 4101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Tawab, M. Do We Need Plant Food Supplements? A Critical Examination of Quality, Safety, Efficacy, and Necessity for a New Regulatory Framework. Planta Med. 2018, 84, 372–393. [Google Scholar] [CrossRef] [Green Version]
- Cohen, P.A.; Venhuis, B.J. Adulterated sexual enhancement supplements: More than mojo. JAMA Intern. Med. 2013, 173, 1169–1170. [Google Scholar] [CrossRef]
- Rocha, T.; Amaral, J.S.; Oliveira, M. Adulteration of Dietary Supplements by the Illegal Addition of Synthetic Drugs: A Review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Meins, J.; Artaria, C.; Riva, A.; Morazzoni, P.; Schubert-Zsilavecz, M.; Abdel-Tawab, M. Survey on the Quality of the Top-Selling European and American Botanical Dietary Supplements Containing Boswellic Acids. Planta Med. 2016, 82, 573–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmiech, M.; Lang, S.J.; Ulrich, J.; Werner, K.; Rashan, L.J.; Syrovets, T.; Simmet, T. Comparative Investigation of Frankincense Nutraceuticals: Correlation of Boswellic and Lupeolic Acid Contents with Cytokine Release Inhibition and Toxicity against Triple-Negative Breast Cancer Cells. Nutrients 2019, 11, 2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaible, A.M.; Traber, H.; Temml, V.; Noha, S.M.; Filosa, R.; Peduto, A.; Weinigel, C.; Barz, D.; Schuster, D.; Werz, O. Potent inhibition of human 5-lipoxygenase and microsomal prostaglandin E(2) synthase-1 by the anti-carcinogenic and anti-inflammatory agent embelin. Biochem. Pharmacol. 2013, 86, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Surette, M.E.; Palmantier, R.; Gosselin, J.; Borgeat, P. Lipopolysaccharides prime whole human blood and isolated neutrophils for the increased synthesis of 5-lipoxygenase products by enhancing arachidonic acid availability: Involvement of the CD14 antigen. J. Exp. Med. 1993, 178, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Romp, E.; Arakandy, V.; Fischer, J.; Wolz, C.; Siegmund, A.; Loffler, B.; Tuchscherr, L.; Werz, O.; Garscha, U. Exotoxins from Staphylococcus aureus activate 5-lipoxygenase and induce leukotriene biosynthesis. Cell Mol. Life Sci. 2020, 77, 3841–3858. [Google Scholar] [CrossRef] [PubMed]
- Safayhi, H.; Boden, S.E.; Schweizer, S.; Ammon, H.P. Concentration-dependent potentiating and inhibitory effects of Boswellia extracts on 5-lipoxygenase product formation in stimulated PMNL. Planta Med. 2000, 66, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, N.C.; Gerstmeier, J.; Schexnaydre, E.E.; Borner, F.; Garscha, U.; Neau, D.B.; Werz, O.; Newcomer, M.E. Structural and mechanistic insights into 5-lipoxygenase inhibition by natural products. Nat. Chem. Biol. 2020, 16, 783–790. [Google Scholar] [CrossRef]
- Syrovets, T.; Buchele, B.; Krauss, C.; Laumonnier, Y.; Simmet, T. Acetyl-boswellic acids inhibit lipopolysaccharide-mediated TNF-alpha induction in monocytes by direct interaction with IkappaB kinases. J. Immunol. 2005, 174, 498–506. [Google Scholar] [CrossRef]
- Chiang, N.; Serhan, C.N. Specialized pro-resolving mediator network: An update on production and actions. Essays Biochem. 2020, 64, 443–462. [Google Scholar]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Levy, B.D. Resolvins in inflammation: Emergence of the pro-resolving superfamily of mediators. J. Clin. Investig. 2018, 128, 2657–2669. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.; Bruning, G.; Bergmann, J.; Jauch, J. A thin-layer chromatography method for the identification of three different olibanum resins (Boswellia serrata, Boswellia papyrifera and Boswellia carterii, respectively, Boswellia sacra). Phytochem. Anal. 2012, 23, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Buchele, B.; Simmet, T. Analysis of 12 different pentacyclic triterpenic acids from frankincense in human plasma by high-performance liquid chromatography and photodiode array detection. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2003, 795, 355–362. [Google Scholar] [CrossRef]
- Frank, A.; Unger, M. Analysis of frankincense from various Boswellia species with inhibitory activity on human drug metabolising cytochrome P450 enzymes using liquid chromatography mass spectrometry after automated on-line extraction. J. Chromatogr. A 2006, 1112, 255–262. [Google Scholar] [CrossRef]
- Lai, S.C.; You, R.I.; Chen, T.T.; Chang, Y.; Liu, C.Z.; Chen, H.P.; Wu, C. Rapid Identification of Commercial Frankincense Products by MALDI-TOF Mass Spectrometry. Comb. Chem. High. Throughput Screen. 2021. [Google Scholar] [CrossRef]
- Sharma, N.; Bhardwaj, V.; Singh, S.; Ali, S.A.; Gupta, D.K.; Paul, S.; Satti, N.K.; Chandra, S.; Verma, M.K. Simultaneous quantification of triterpenoic acids by high performance liquid chromatography method in the extracts of gum resin of Boswellia serrata obtained by different extraction techniques. Chem. Cent. J. 2016, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Safayhi, H.; Mack, T.; Sabieraj, J.; Anazodo, M.I.; Subramanian, L.R.; Ammon, H.P. Boswellic acids: Novel, specific, nonredox inhibitors of 5-lipoxygenase. J. Pharmacol Exp. Ther 1992, 261, 1143–1146. [Google Scholar]
- Siemoneit, U.; Koeberle, A.; Rossi, A.; Dehm, F.; Verhoff, M.; Reckel, S.; Maier, T.J.; Jauch, J.; Northoff, H.; Bernhard, F.; et al. Inhibition of microsomal prostaglandin E2 synthase-1 as a molecular basis for the anti-inflammatory actions of boswellic acids from frankincense. Br. J. Pharmacol. 2011, 162, 147–162. [Google Scholar] [CrossRef] [Green Version]
- Bergqvist, F.; Morgenstern, R.; Jakobsson, P.J. A review on mPGES-1 inhibitors: From preclinical studies to clinical applications. Prostaglandins Other Lipid Mediat. 2020, 147, 106383. [Google Scholar] [CrossRef]
- Smith, W.L.; Urade, Y.; Jakobsson, P.J. Enzymes of the cyclooxygenase pathways of prostanoid biosynthesis. Chem. Rev. 2011, 111, 5821–5865. [Google Scholar] [CrossRef] [Green Version]
- Verhoff, M.; Seitz, S.; Paul, M.; Noha, S.M.; Jauch, J.; Schuster, D.; Werz, O. Tetra- and pentacyclic triterpene acids from the ancient anti-inflammatory remedy frankincense as inhibitors of microsomal prostaglandin E(2) synthase-1. J. Nat. Prod. 2014, 77, 1445–1451. [Google Scholar] [CrossRef]
- Syrovets, T.; Gschwend, J.E.; Buchele, B.; Laumonnier, Y.; Zugmaier, W.; Genze, F.; Simmet, T. Inhibition of IkappaB kinase activity by acetyl-boswellic acids promotes apoptosis in androgen-independent PC-3 prostate cancer cells in vitro and in vivo. J. Biol. Chem. 2005, 280, 6170–6180. [Google Scholar] [CrossRef] [Green Version]
- Catanzaro, D.; Rancan, S.; Orso, G.; Dall’Acqua, S.; Brun, P.; Giron, M.C.; Carrara, M.; Castagliuolo, I.; Ragazzi, E.; Caparrotta, L.; et al. Boswellia serrata Preserves Intestinal Epithelial Barrier from Oxidative and Inflammatory Damage. PLoS ONE 2015, 10, e0125375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Governa, P.; Marchi, M.; Cocetta, V.; De Leo, B.; Saunders, P.T.K.; Catanzaro, D.; Miraldi, E.; Montopoli, M.; Biagi, M. Effects of Boswellia Serrata Roxb. and Curcuma longa L. in an In Vitro Intestinal Inflammation Model Using Immune Cells and Caco-2. Pharmaceuticals 2018, 11, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabas, I.; Glass, C.K. Anti-inflammatory therapy in chronic disease: Challenges and opportunities. Science 2013, 339, 166–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammarberg, T.; Radmark, O. 5-lipoxygenase binds calcium. Biochemistry 1999, 38, 4441–4447. [Google Scholar] [CrossRef]
- Steinhilber, D.; Herrmann, T.; Roth, H.J. Separation of lipoxins and leukotrienes from human granulocytes by high-performance liquid chromatography with a Radial-Pak cartridge after extraction with an octadecyl reversed-phase column. J. Chromatogr. 1989, 493, 361–366. [Google Scholar] [CrossRef]
- Pace, S.; Pergola, C.; Dehm, F.; Rossi, A.; Gerstmeier, J.; Troisi, F.; Pein, H.; Schaible, A.M.; Weinigel, C.; Rummler, S.; et al. Androgen-mediated sex bias impairs efficiency of leukotriene biosynthesis inhibitors in males. J. Clin. Investig 2017, 127, 3167–3176. [Google Scholar] [CrossRef] [Green Version]
- Werner, M.; Jordan, P.M.; Romp, E.; Czapka, A.; Rao, Z.; Kretzer, C.; Koeberle, A.; Garscha, U.; Pace, S.; Claesson, H.E.; et al. Targeting biosynthetic networks of the proinflammatory and proresolving lipid metabolome. FASEB J. 2019, 33, 6140–6153. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Batch | Dosage Form | Declared Extract Content | KBA | AKBA | αBA | β-BA | AαBA | AβBA | ∑ of BA |
|---|---|---|---|---|---|---|---|---|---|---|
| Sallaki® tablets | AB18024 | tablets | 400 | 9.34 | 7.88 | 20.73 | 52.65 | 7.77 | 25.72 | 124.07 |
| H15 Ayurmedica® | 171 | oily capsules | 400 | 0.30 | 0.31 | 0.40 | 0.27 | 0.33 | 0.32 | 1.92 |
| BOSWELLIASAN® | HBPC11 | hard gelatin capsule | 300 | 5.12 | 7.51 | 13.58 | 36.40 | 6.45 | 21.16 | 90.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Börner, F.; Werner, M.; Ertelt, J.; Meins, J.; Abdel-Tawab, M.; Werz, O. Analysis of Boswellic Acid Contents and Related Pharmacological Activities of Frankincense-Based Remedies That Modulate Inflammation. Pharmaceuticals 2021, 14, 660. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14070660
Börner F, Werner M, Ertelt J, Meins J, Abdel-Tawab M, Werz O. Analysis of Boswellic Acid Contents and Related Pharmacological Activities of Frankincense-Based Remedies That Modulate Inflammation. Pharmaceuticals. 2021; 14(7):660. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14070660
Chicago/Turabian StyleBörner, Friedemann, Markus Werner, Johannes Ertelt, Jürgen Meins, Mona Abdel-Tawab, and Oliver Werz. 2021. "Analysis of Boswellic Acid Contents and Related Pharmacological Activities of Frankincense-Based Remedies That Modulate Inflammation" Pharmaceuticals 14, no. 7: 660. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14070660