
Delaying Effects of Prolactin and Growth Hormone on Aging Processes in Bovine Oocytes Matured In Vitro



Abstract
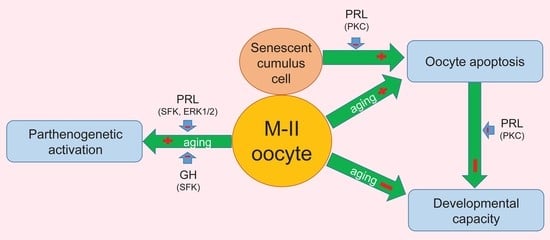
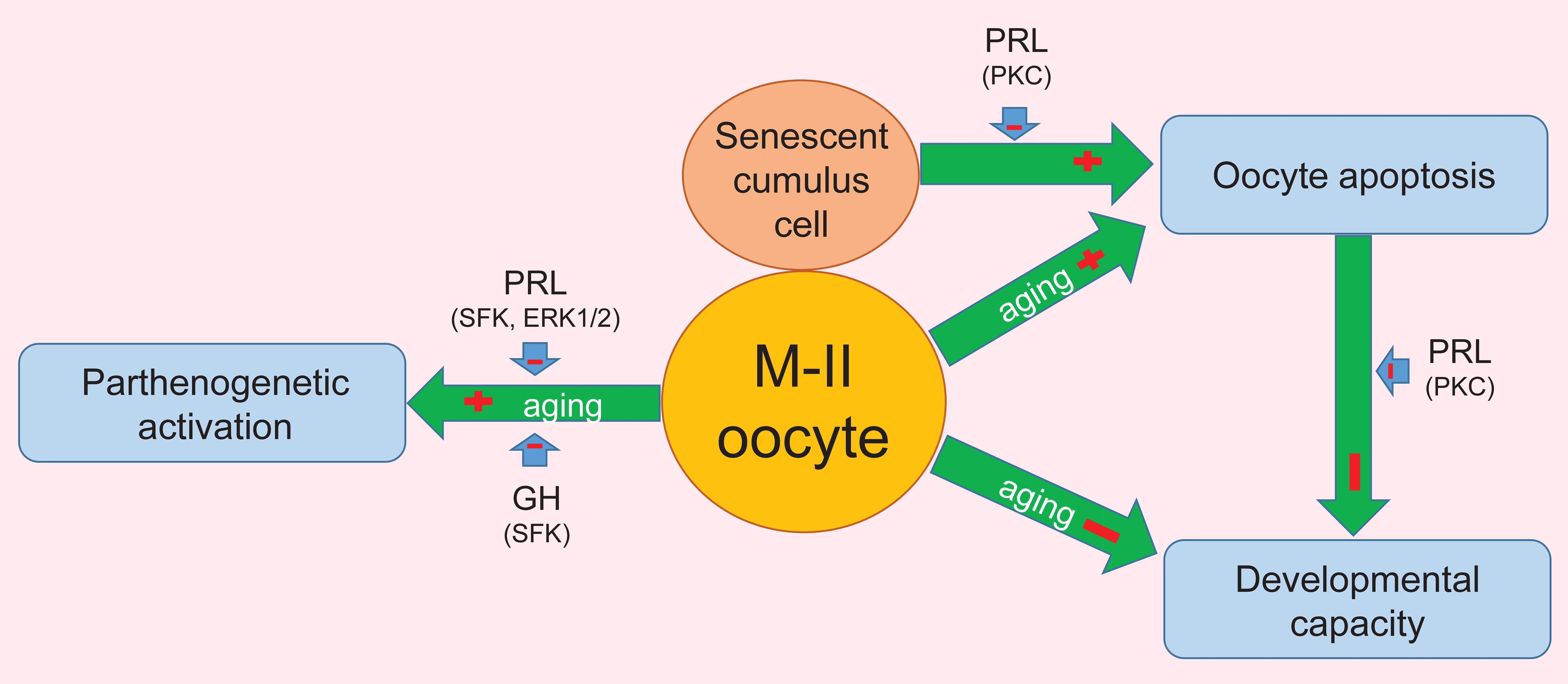
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
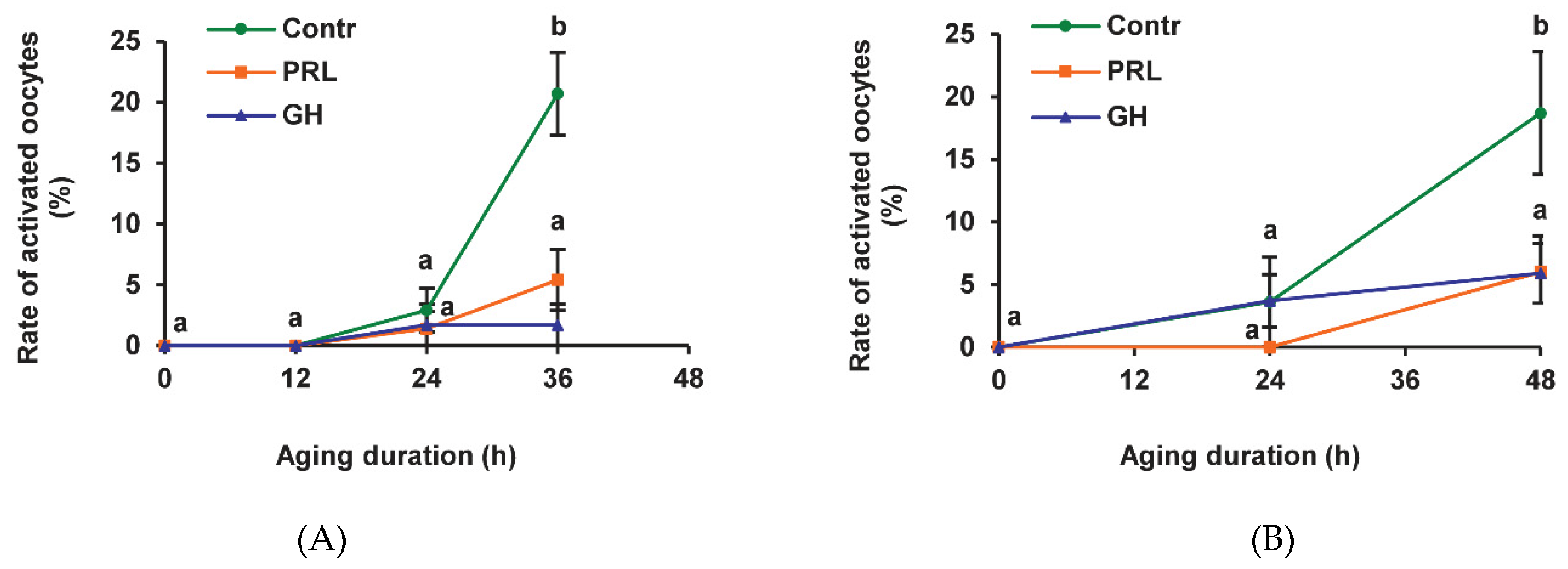
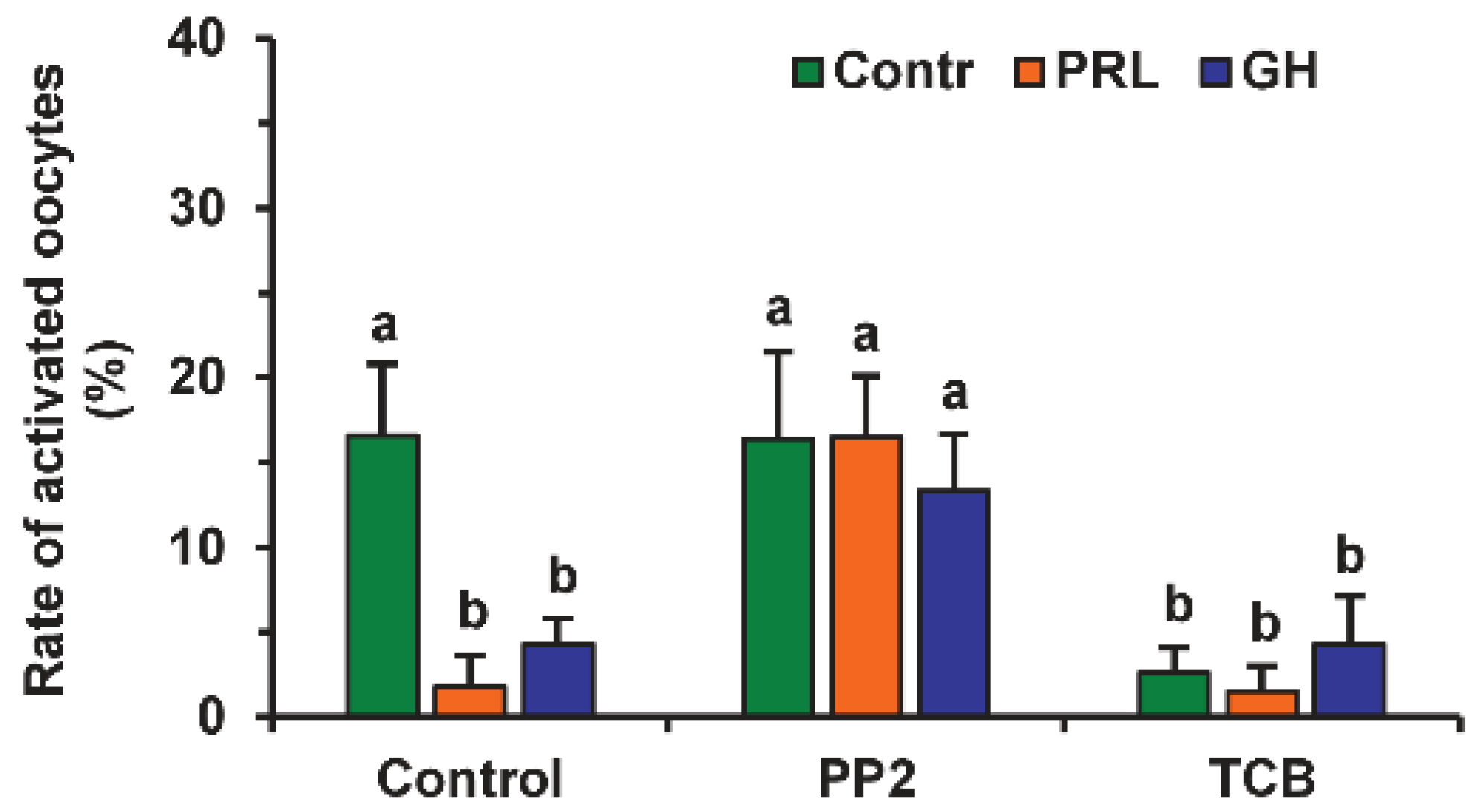
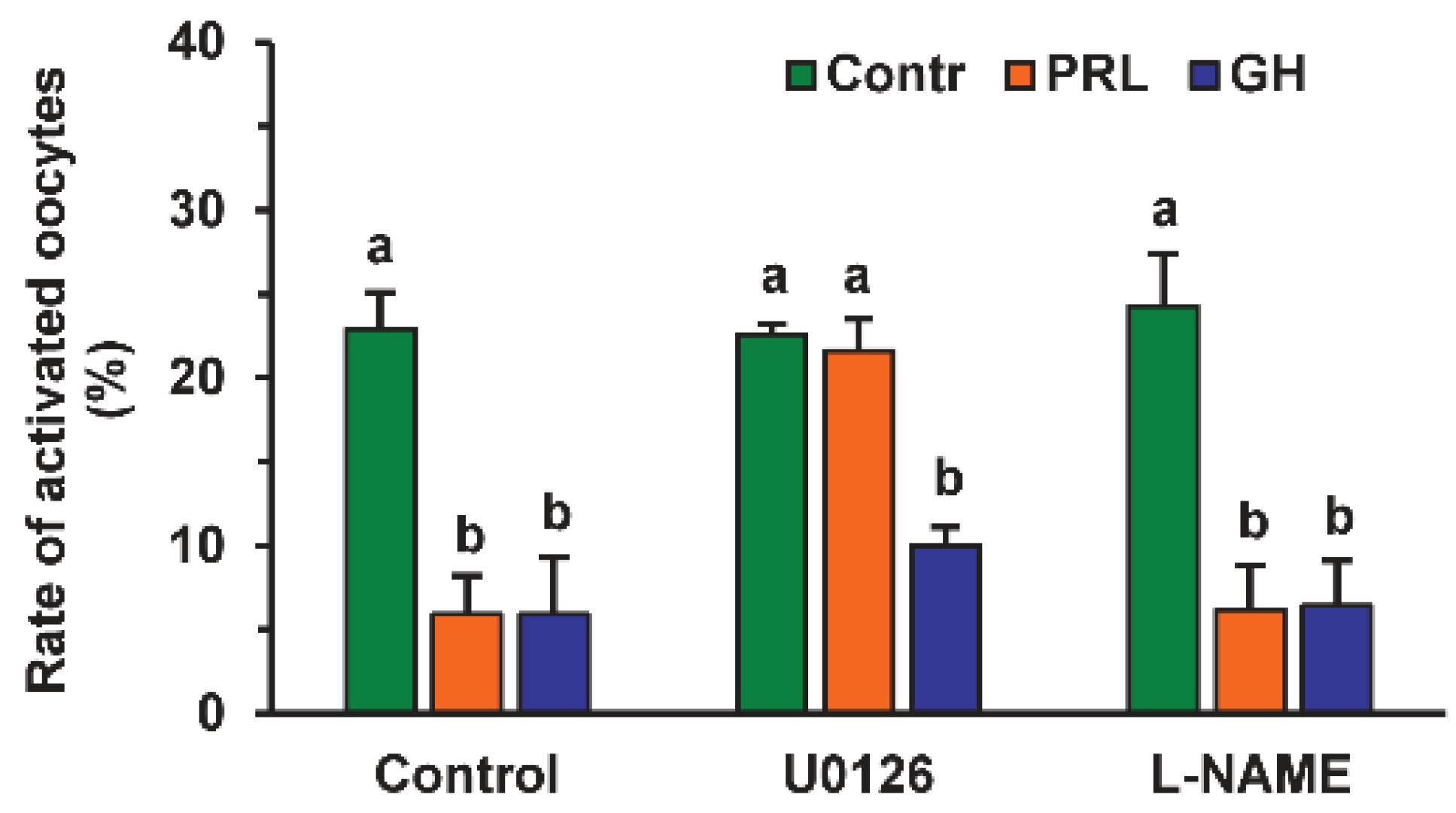
2.1. Effects of PRL and GH on Spontaneous Parthenogenetic Activation (SPA) of Aging Bovine Oocytes
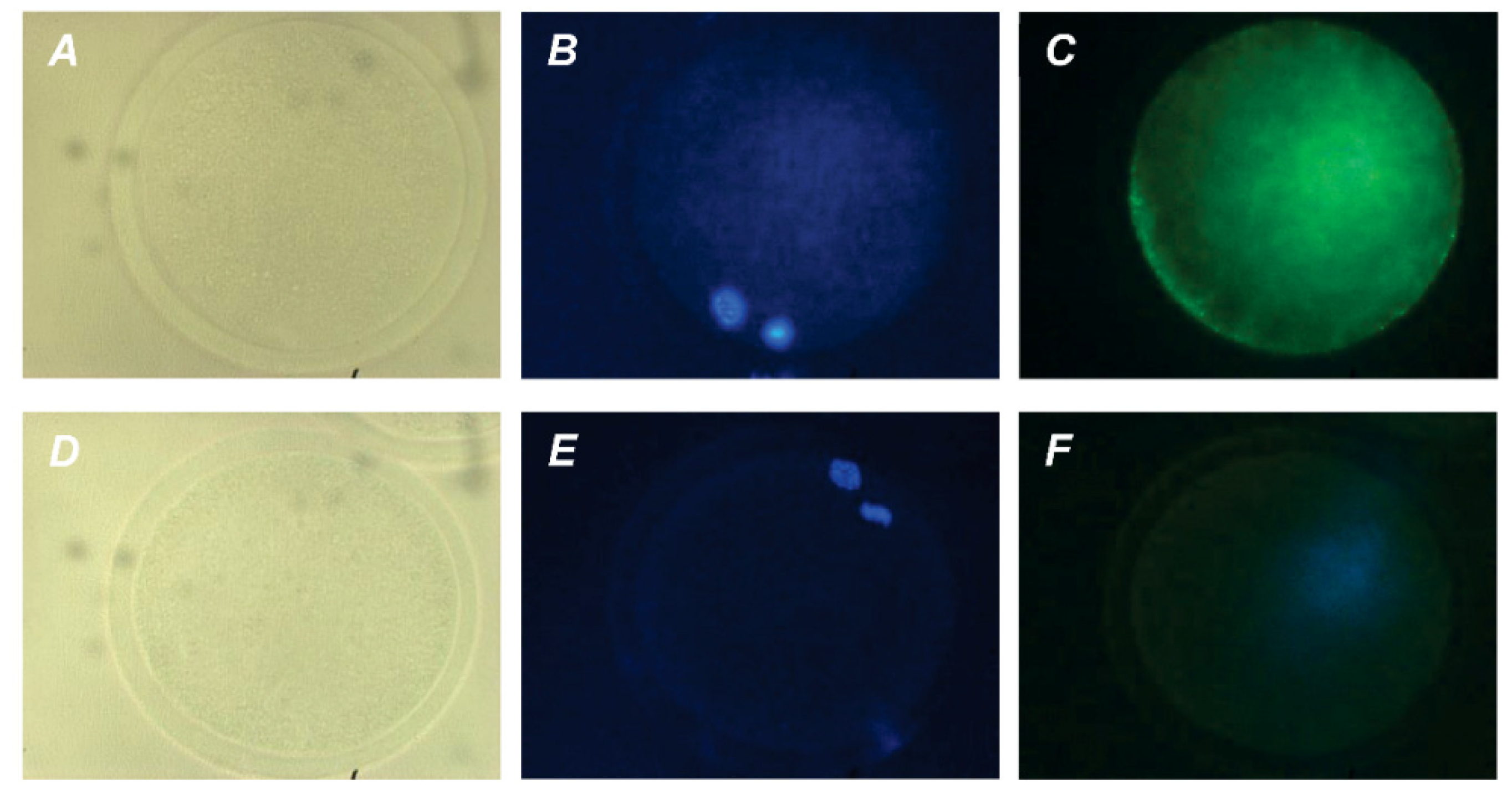
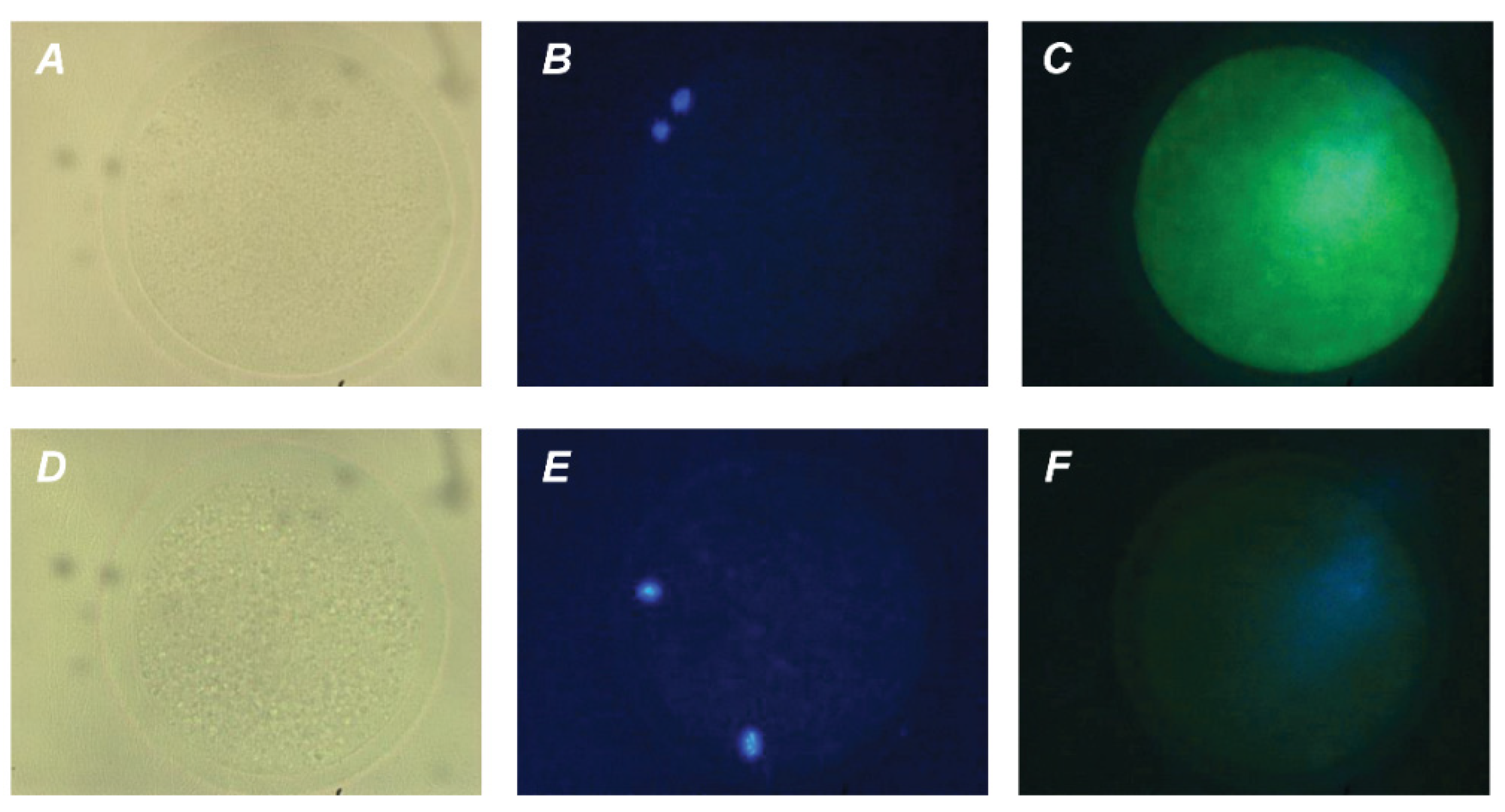
2.2. Localization of PRL and GH Receptors in Matured Bovine Oocytes
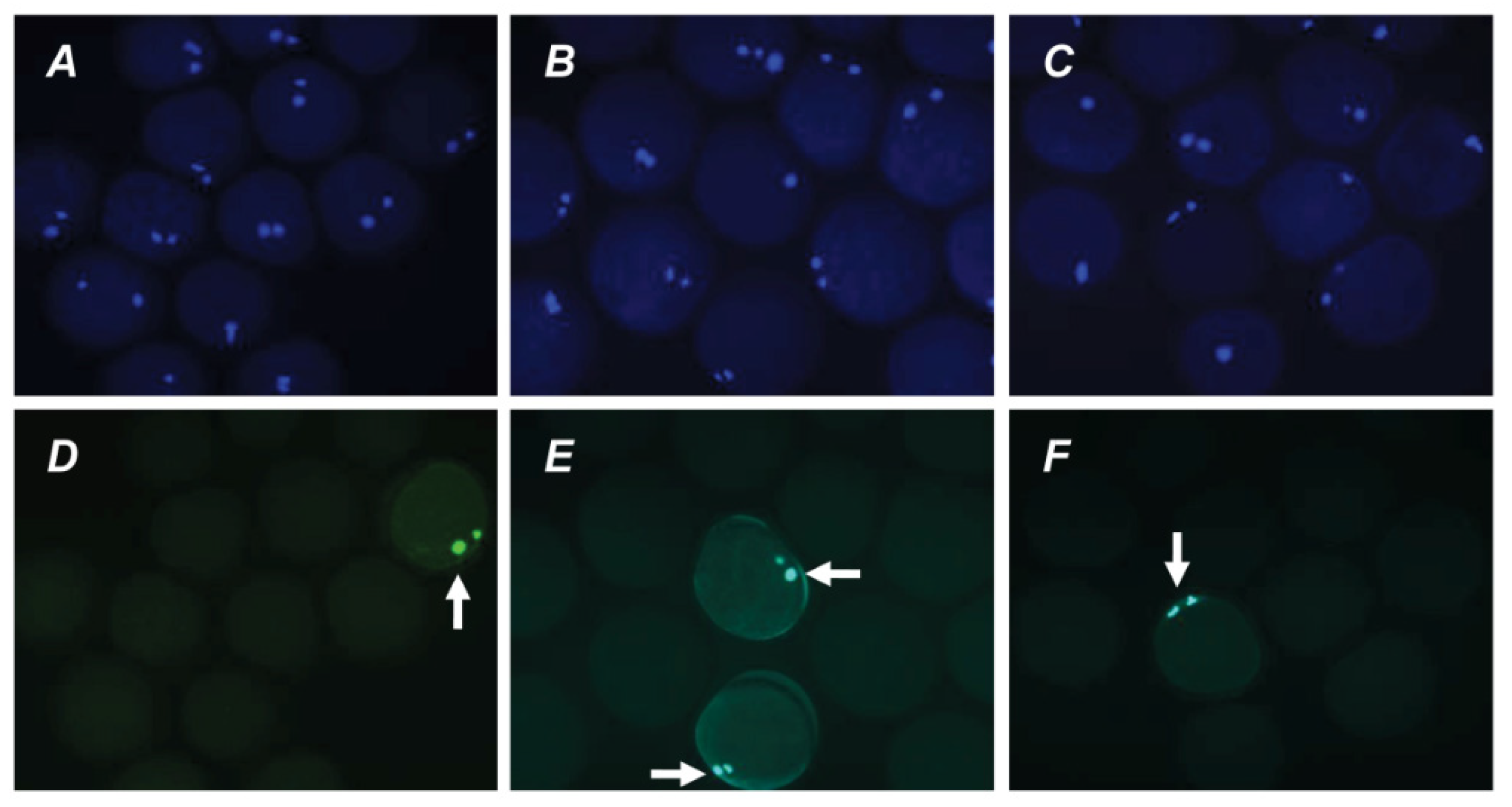
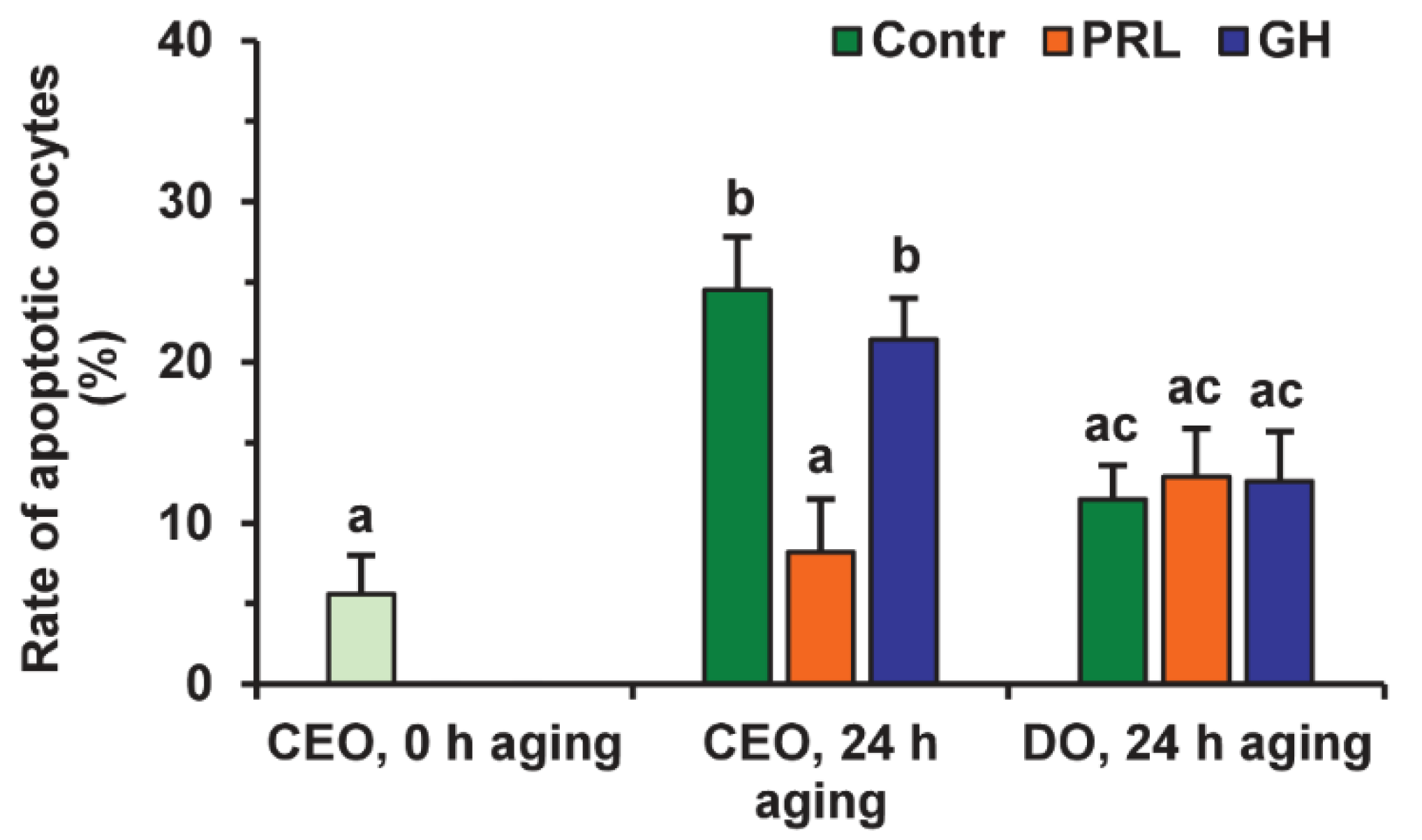
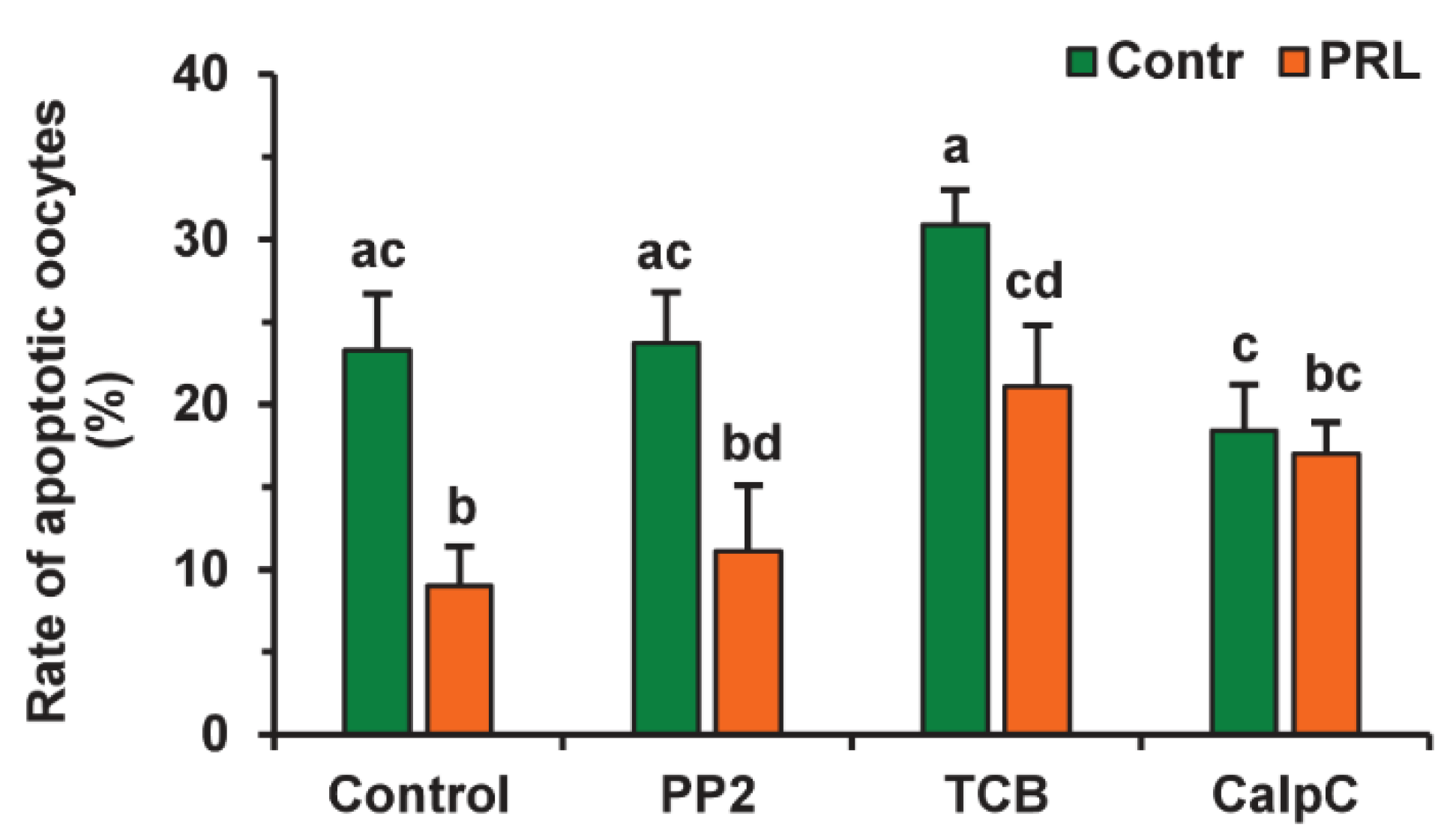
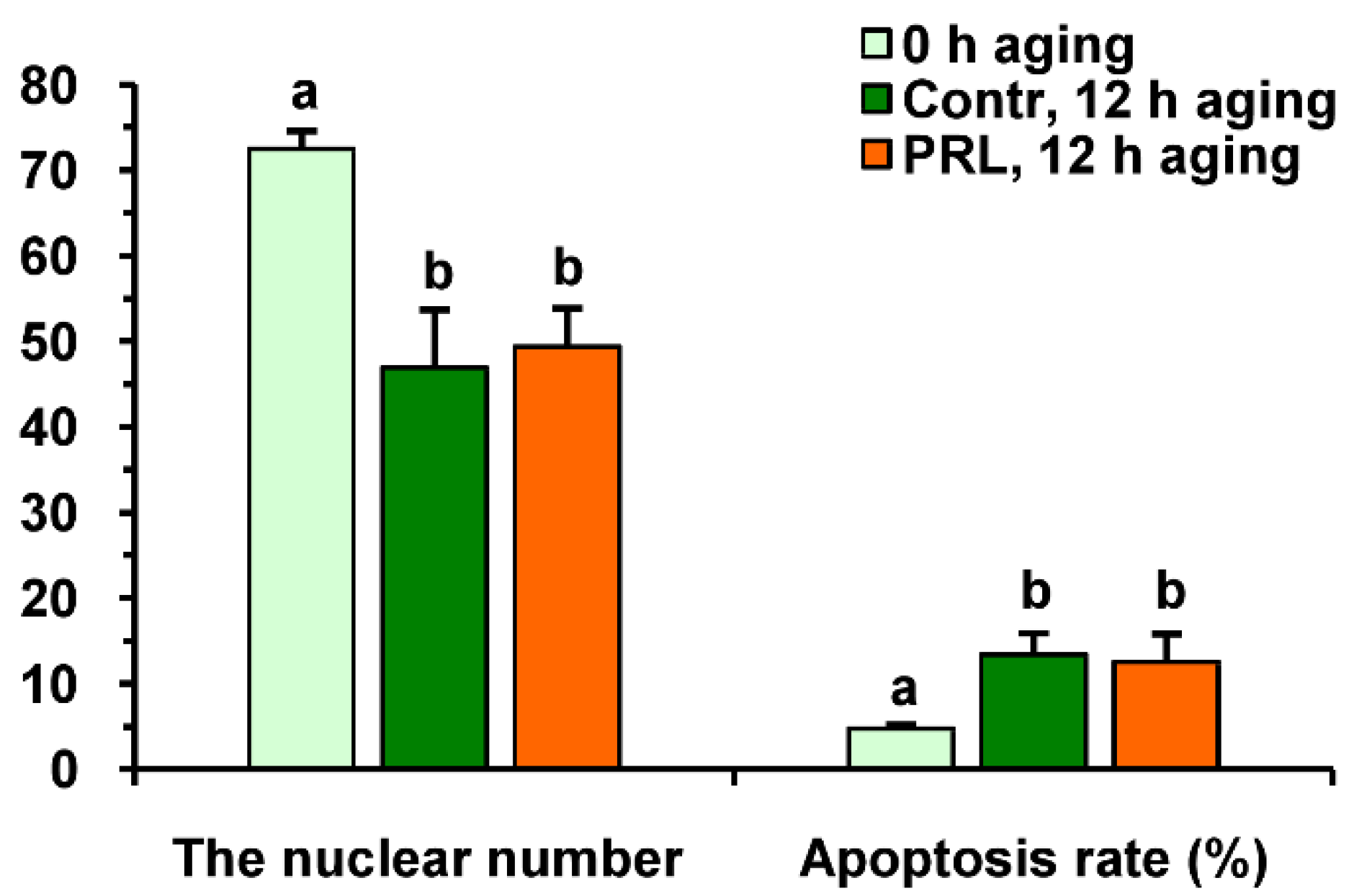
2.3. Effects of PRL on the Apoptosis of Aging Bovine Oocytes
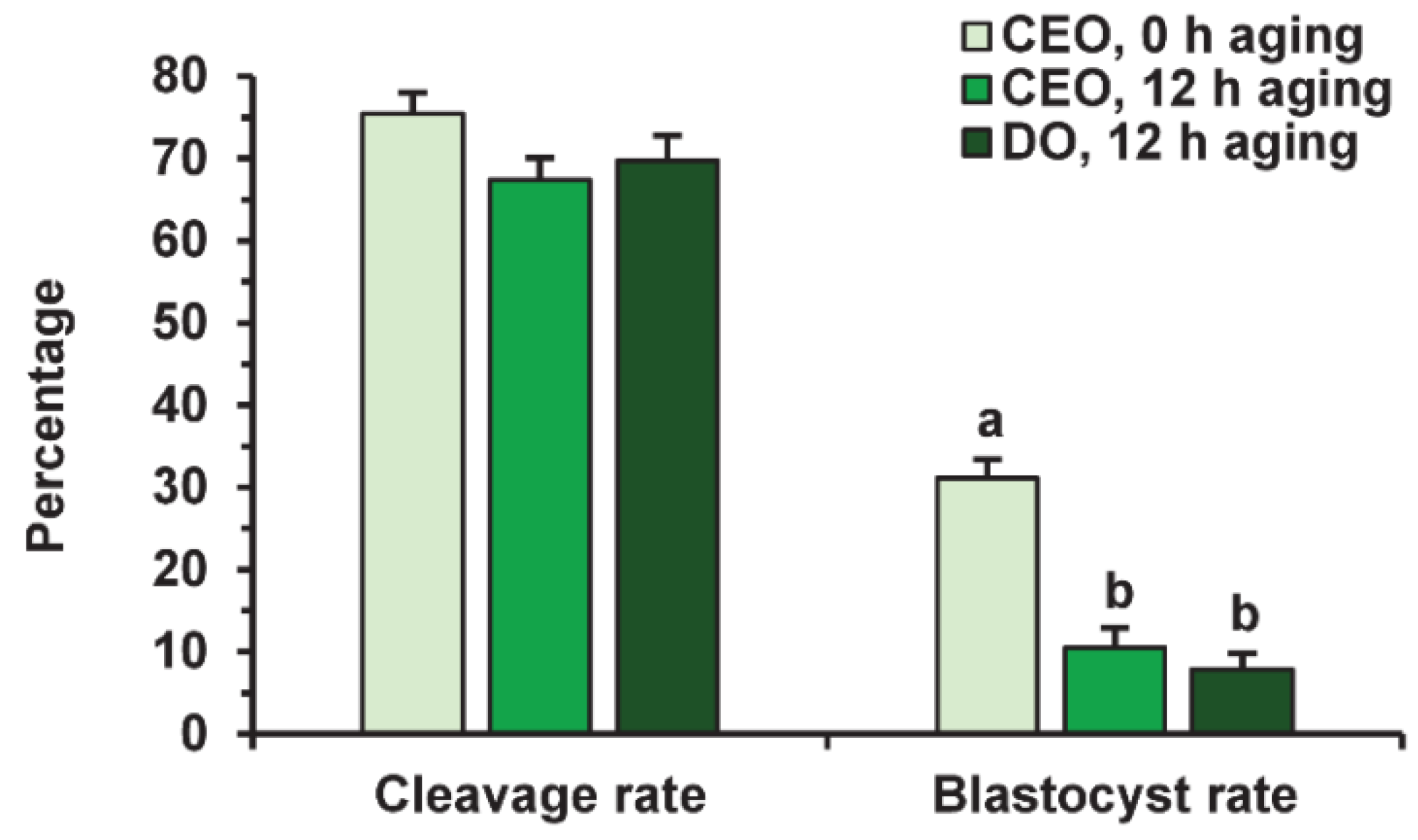
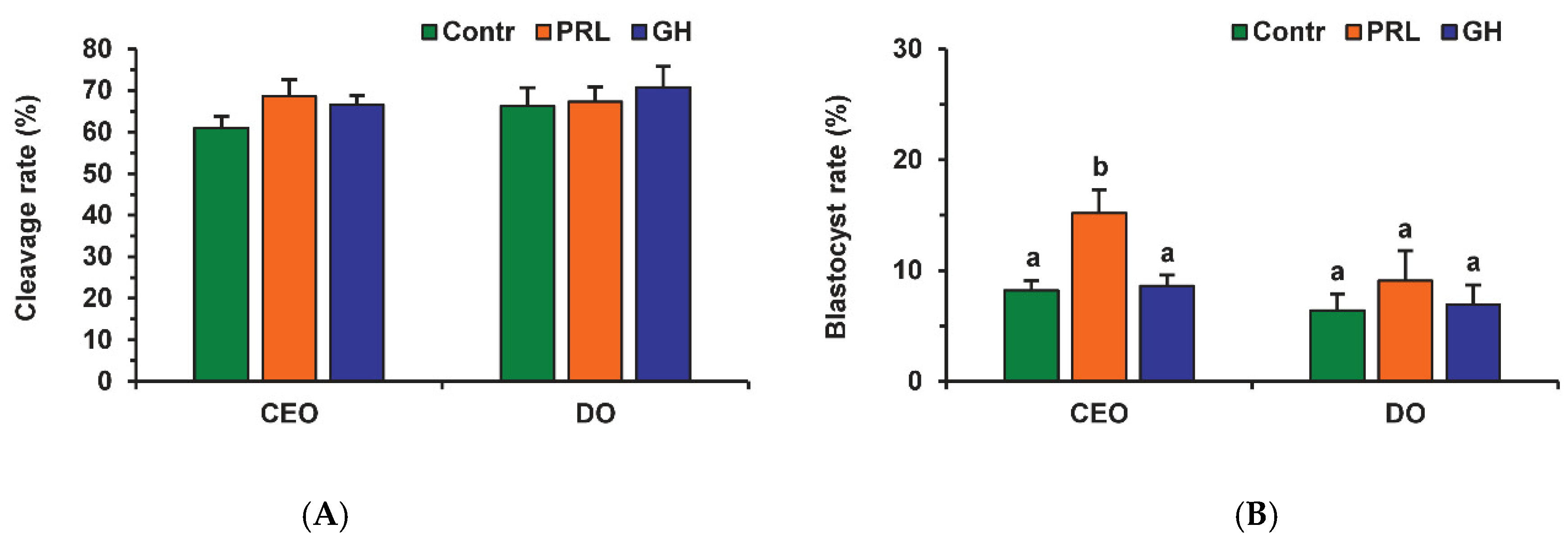
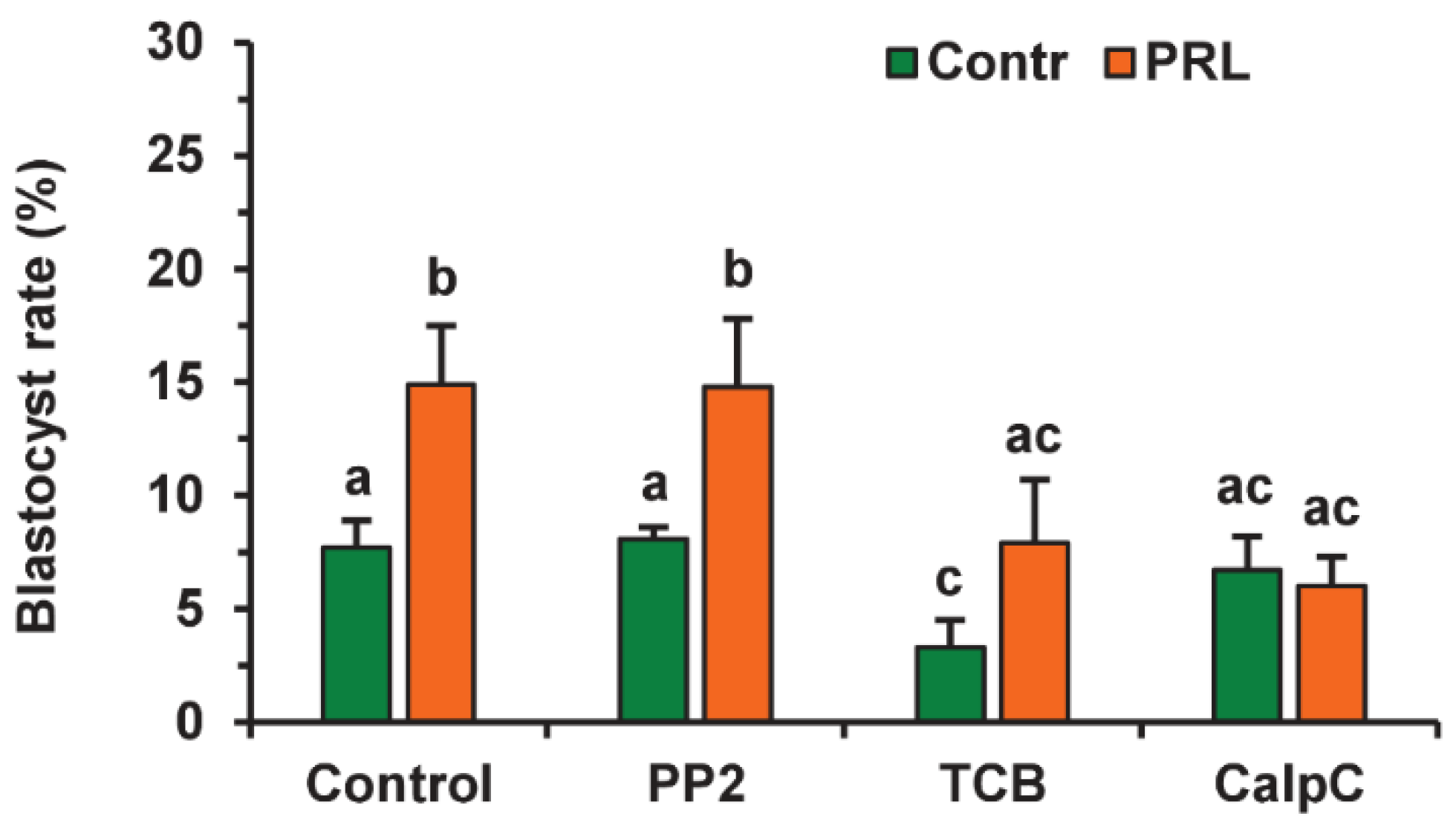
2.4. Effects of PRL on the Developmental Capacity of Aging Bovine Oocytes
3. Discussion
4. Materials and Methods
4.1. Isolation of Oocytes and Conditions of In Vitro Maturation (IVM)
4.2. Prolonged Culture of Aging Oocytes
4.3. Assessment of Oocyte Activation
4.4. Immunofluorescent and Immunocytochemical Detection of Receptors
4.5. In Vitro Fertilization (IVF) and Embryo Culture (IVC)
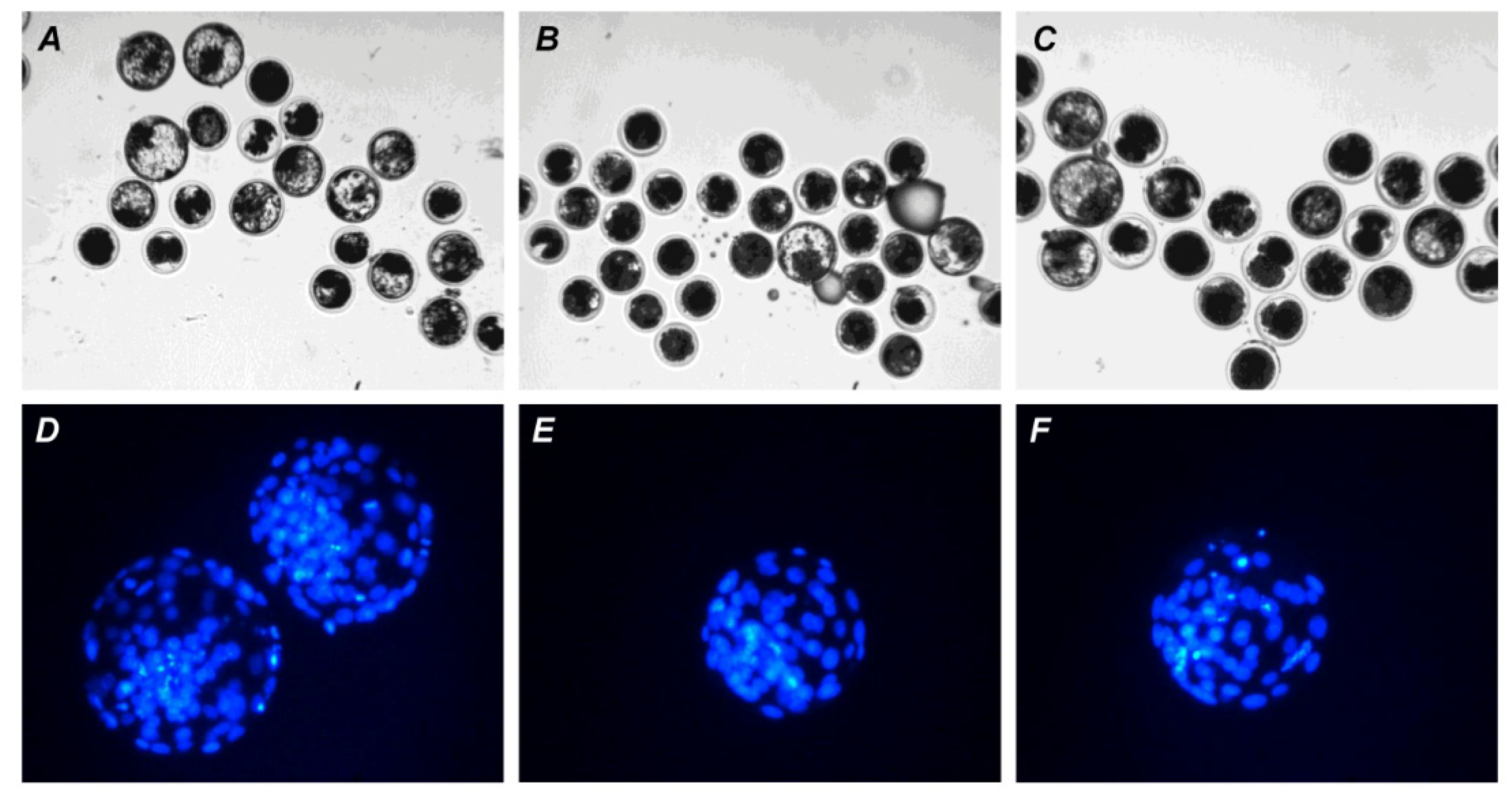
4.6. Terminal Deoxynucleotidyl Transferase-Mediated 2′-Deoxyuridine 5′-Triphosphate (dUTP) Nick-End Labeling (TUNEL) Assay and DAPI Staining
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miao, Y.L.; Kikuchi, K.; Sun, Q.Y.; Schatten, H. Oocyte aging: Cellular and molecular changes, developmental potential and reversal possibility. Hum. Reprod. Update 2009, 15, 573–585. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Igarashi, H.; Amita, M.; Hara, S.; Matsuo, K.; Kurachi, H. Molecular mechanism of poor embryo development in postovulatory aged oocytes: Mini review. J. Obstet. Gynaecol. Res. 2013, 39, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Tiwari, M.; Koch, B.; Chaube, S.K. Morphological, cellular and molecular changes during postovulatory egg aging in mammals. J. Biomed. Sci. 2015, 22, 36. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, A.J.; Weinberg, C.R.; Baird, D.D. Post-ovulatory ageing of the human oocyte and embryo failure. Hum. Reprod. 1998, 13, 394–397. [Google Scholar] [CrossRef]
- Tarín, J.J.; Pérez-Albalá, S.; Pérez-Hoyos, S.; Cano, A. Postovulatory aging of oocytes decreases reproductive fitness and longevity of offspring. Biol. Reprod. 2002, 66, 495–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, F.; Sun, Q.; Li, G.; Yao, Z.; Chen, P.; Wan, C.; Zhong, H.; Zeng, Y. Perinatal and neonatal outcomes of pregnancies after early rescue intracytoplasmic sperm injection in women with primary infertility compared with conventional intracytoplasmic sperm injection: A retrospective 6-year study. BMC Pregnancy Childbirth 2020, 20, 460. [Google Scholar] [CrossRef] [PubMed]
- Cui, W. Oocyte spontaneous activation: An overlooked cellular event that impairs female fertility in mammals. Front. Cell Dev. Biol. 2021, 9, 648057. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Tiwari, M.; Chaube, S.K. Abortive spontaneous egg activation: An emerging biological threat for the existence of mammals. Cell. Reprogram. 2017, 19, 145–149. [Google Scholar] [CrossRef]
- Igarashi, H.; Takahashi, T.; Takahashi, E.; Tezuka, N.; Nakahara, K.; Takahashi, K.; Kurachi, H. Aged mouse oocytes fail to readjust intracellular adenosine triphosphates at fertilization. Biol. Reprod. 2005, 72, 1256–1261. [Google Scholar] [CrossRef]
- Tatone, C.; Carbone, M.C.; Gallo, R.; Delle Monache, S.; Di Cola, M.; Alesse, E.; Amicarelli, F. Age-associated changes in mouse oocytes during postovulatory in vitro culture: Possible role for meiotic kinases and survival factor BCL2. Biol. Reprod. 2006, 74, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Igarashi, H.; Kawagoe, J.; Amita, M.; Hara, S.; Kurachi, H. Poor embryo development in mouse oocytes aged in vitro is associated with impaired calcium homeostasis. Biol. Reprod. 2009, 80, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.J.; Schatten, H.; Zhang, C.L.; Sun, Q.Y. Oocyte ageing and epigenetics. Reproduction 2015, 149, R103–R114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.W.; Lee, S.E.; Park, Y.G.; Kim, W.J.; Park, H.J.; Park, C.O.; Kim, S.H.; Oh, S.H.; Lee, D.G.; Pyeon, D.B.; et al. The antioxidant icariin protects porcine oocytes from age-related damage in vitro. Anim. Biosci. 2021, 34, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Petr, J.; Krejčová, M.; Rajmon, R.; Jílek, F. Activation of protein kinase C suppresses fragmentation of pig oocytes aged in vitro. Animal 2011, 5, 565–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecconi, S.; Rossi, G.; Deldar, H.; Cellini, V.; Patacchiola, F.; Carta, G.; Macchiarelli, G.; Canipari, R. Post-ovulatory ageing of mouse oocytes affects the distribution of specific spindle-associated proteins and Akt expression levels. Reprod. Fertil. Dev. 2014, 26, 562–569. [Google Scholar] [CrossRef]
- McGinnis, L.K.; Pelech, S.; Kinsey, W.H. Post-ovulatory aging of oocytes disrupts kinase signaling pathways and lysosome biogenesis. Mol. Reprod. Dev. 2014, 81, 928–945. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.Q.; Wang, J.; Xiao, B.; Lin, F.H.; Zhu, J.; Sun, G.Y.; Luo, M.J.; Tan, J.H. Cumulus cell-released tumor necrosis factor (TNF)-α promotes post-ovulatory aging of mouse oocytes. Aging 2018, 10, 1745–1757. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Ahmed, S.M.; El-Gammal, Z.; Shouman, S.; Ahmed, A.; Mansour, R.; El-Badri, N. Oocyte aging: The role of cellular and environmental factors and impact on female fertility. Adv. Exp. Med. Biol. 2020, 1247, 109–123. [Google Scholar] [CrossRef]
- Lord, T.; Nixon, B.; Jones, K.T.; Aitken, R.J. Melatonin prevents postovulatory oocyte aging in the mouse and extends the window for optimal fertilization in vitro. Biol. Reprod. 2013, 88, 67. [Google Scholar] [CrossRef]
- Wang, T.; Gao, Y.Y.; Chen, L.; Nie, Z.W.; Cheng, W.; Liu, X.; Schatten, H.; Zhang, X.; Miao, Y.L. Melatonin prevents postovulatory oocyte aging and promotes subsequent embryonic development in the pig. Aging 2017, 9, 1552–1564. [Google Scholar] [CrossRef] [Green Version]
- Shu, Y.M.; Zeng, H.T.; Ren, Z.; Zhuang, G.L.; Liang, X.Y.; Shen, H.W.; Yao, S.Z.; Ke, P.Q.; Wang, N.N. Effects of cilostamide and forskolin on the meiotic resumption and embryonic development of immature human oocytes. Hum. Reprod. 2008, 23, 504–513. [Google Scholar] [CrossRef] [Green Version]
- Sasseville, M.; Albuz, F.K.; Côté, N.; Guillemette, C.; Gilchrist, R.B.; Richard, F.J. Characterization of novel phosphodiesterases in the bovine ovarian follicle. Biol. Reprod. 2009, 81, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.R.; Schoevers, E.J.; Roelen, B.A. Usefulness of bovine and porcine IVM/IVF models for reproductive toxicology. Reprod. Biol. Endocrinol. 2014, 12, 117. [Google Scholar] [CrossRef] [Green Version]
- Lebedeva, I.Y.; Singina, G.N.; Lopukhov, A.V.; Zinovieva, N.A. Dynamics of morphofunctional changes in aging bovine ova during prolonged culture in vitro. Cell Tissue Biol. 2014, 8, 258–266. [Google Scholar] [CrossRef]
- Jiang, W.J.; Yao, X.R.; Zhao, Y.H.; Gao, Q.S.; Jin, Q.G.; Li, Y.H.; Yan, A.G.; Xu, Y.N. L-carnitine prevents bovine oocyte aging and promotes subsequent embryonic development. J. Reprod. Dev. 2019, 65, 499–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebedeva, I.Y.; Singina, G.N.; Lopukhov, A.V.; Shedova, E.N.; Zinovieva, N.A. Prolactin and growth hormone affect metaphase-II chromosomes in aging oocytes via cumulus cells using similar signaling pathways. Front. Genet. 2015, 6, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebedeva, I.Y.; Singina, G.N.; Shedova, E.N.; Lopukhov, A.V.; Zinov’eva, N.A. The role of different NO synthase isoforms in the regulatory effect of prolactin and growth hormone on the state of chromosomes in mature oocytes aging in vitro. Cell Tissue Biol. 2019, 13, 219–227. [Google Scholar] [CrossRef]
- Huang, Y.; Kim, S.O.; Jiang, J.; Frank, S.J. Growth hormone-induced phosphorylation of epidermal growth factor (EGF) receptor in 3T3-F442A cells. Modulation of EGF-induced trafficking and signaling. J. Biol. Chem. 2003, 278, 18902–18913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutzman, J.H.; Rugowski, D.E.; Schroeder, M.D.; Watters, J.J.; Schuler, L.A. Multiple kinase cascades mediate prolactin signals to activating protein-1 in breast cancer cells. Mol. Endocrinol. 2004, 18, 3064–3075. [Google Scholar] [CrossRef] [Green Version]
- Cecconi, S.; Mauro, A.; Cellini, V.; Patacchiola, F. The role of Akt signalling in the mammalian ovary. Int. J. Dev. Biol. 2012, 56, 809–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kheilová, K.; Petr, J.; Žalmanová, T.; Kučerová-Chrpová, V.; Řehák, D. Src family kinases are involved in the meiotic maturation of porcine oocytes. Reprod. Fertil. Dev. 2015, 27, 1097–1105. [Google Scholar] [CrossRef]
- Tiwari, M.; Prasad, S.; Pandey, A.N.; Premkumar, K.V.; Tripathi, A.; Gupta, A.; Chetan, D.R.; Yadav, P.K.; Shrivastav, T.G.; Chaube, S.K. Nitric oxide signaling during meiotic cell cycle regulation in mammalian oocytes. Front. Biosci. 2017, 9, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Lebedeva, I.Y.; Singina, G.N.; Volkova, N.A.; Vejlsted, M.; Zinovieva, N.A.; Schmidt, M. Prolactin affects bovine oocytes through direct and cumulus-mediated pathways. Theriogenology 2014, 82, 1154–1164. [Google Scholar] [CrossRef] [PubMed]
- Kölle, S.; Sinowatz, F.; Boie, G.; Lincoln, D. Developmental changes in the expression of the growth hormone receptor messenger ribonucleic acid and protein in the bovine ovary. Biol. Reprod. 1998, 59, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Lechniak, D.; Cieślak, D.; Sosnowski, J. Cytogenetic analysis of bovine parthenotes after spontaneous activation in vitro. Theriogenology 1998, 49, 779–785. [Google Scholar] [CrossRef]
- Budna, J.; Bryja, A.; Celichowski, P.; Kahan, R.; Kranc, W.; Ciesiółka, S.; Rybska, M.; Borys, S.; Jeseta, M.; Bukowska, D.; et al. Genes of cellular components of morphogenesis in porcine oocytes before and after IVM. Reproduction 2017, 154, 535–545. [Google Scholar] [CrossRef]
- Espada, J.; Martín-Pérez, J. An update on Src family of nonreceptor tyrosine kinases biology. Int. Rev. Cell. Mol. Biol. 2017, 331, 83–122. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, Y.; Lee, C.M.M.; Müller, A.F.; Brooks, A.J. GHR signalling: Receptor activation and degradation mechanisms. Mol. Cell. Endocrinol. 2021, 520, 111075. [Google Scholar] [CrossRef]
- Tomashov-Matar, R.; Levi, M.; Shalgi, R. The involvement of Src family kinases (SFKs) in the events leading to resumption of meiosis. Mol. Cell. Endocrinol. 2008, 282, 56–62. [Google Scholar] [CrossRef]
- Miyagaki, Y.; Kanemori, Y.; Tanaka, F.; Baba, T. Possible role of p38 MAPK-MNK1-EMI2 cascade in metaphase-II arrest of mouse oocytes. Biol. Reprod. 2014, 91, 45. [Google Scholar] [CrossRef] [PubMed]
- Kujjo, L.L.; Perez, G.I. Ceramide and mitochondrial function in aging oocytes: Joggling a new hypothesis and old players. Reproduction 2012, 143, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebedeva, I.Y.; Singina, G.N.; Shedova, E.N.; Lopukhov, A.V.; Mityashova, O.S. Resistance of bovine oocytes to age-related changes after exposure to luteotropic factors during the second phase of culture till the metaphase II stage. Sel’skokhozyaistvennaya Biol. Agric. Biol. 2018, 53, 1202–1211. [Google Scholar] [CrossRef]
- Lebedeva, I.Y.; Singina, G.N.; Taradajnic, T.E.; Zinovieva, N.A. Protein kinase implication in prolactin signaling in bovine oocyte-cumulus complexes. Cell Tissue Biol. 2011, 5, 560–567. [Google Scholar] [CrossRef]
- Kölle, S.; Stojkovic, M.; Boie, G.; Wolf, E.; Sinowatz, F. Growth hormone-related effects on apoptosis, mitosis, and expression of connexin 43 in bovine in vitro maturation cumulus-oocyte complexes. Biol. Reprod. 2003, 68, 1584–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuranaga, E.; Kanuka, H.; Bannai, M.; Suzuki, M.; Nishihara, M.; Takahashi, M. Fas/Fas ligand system in prolactin-induced apoptosis in rat corpus luteum: Possible role of luteal immune cells. Biochem. Biophys. Res. Commun. 1999, 260, 167–173. [Google Scholar] [CrossRef]
- Vallcaneras, S.S.; Delgado, S.M.; Motta, A.; Telleria, C.M.; Rastrilla, A.M.; Casais, M. Effect of prolactin acting on the coeliac ganglion via the superior ovarian nerve on ovarian function in the postpartum lactating and non-lactating rat. Gen. Comp. Endocrinol. 2013, 184, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Olmos-Ortiz, A.; Déciga-García, M.; Preciado-Martínez, E.; Bermejo-Martínez, L.; Flores-Espinosa, P.; Mancilla-Herrera, I.; Irles, C.; Helguera-Repetto, A.C.; Quesada-Reyna, B.; Goffin, V.; et al. Prolactin decreases LPS-induced inflammatory cytokines by inhibiting TLR-4/NFκB signaling in the human placenta. Mol. Hum. Reprod. 2019, 25, 660–667. [Google Scholar] [CrossRef]
- Perks, C.M.; Newcomb, P.V.; Grohmann, M.; Wright, R.J.; Mason, H.D.; Holly, J.M. Prolactin acts as a potent survival factor against C2-ceramide-induced apoptosis in human granulosa cells. Hum. Reprod. 2003, 18, 2672–2677. [Google Scholar] [CrossRef] [Green Version]
- Arav, A.; Zvi, R. Do chilling injury and heat stress share the same mechanism of injury in oocytes? Mol. Cell. Endocrinol. 2008, 282, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Döll, F.; Pfeilschifter, J.; Huwiler, A. Prolactin upregulates sphingosine kinase-1 expression and activity in the human breast cancer cell line MCF7 and triggers enhanced proliferation and migration. Endocr. Relat. Cancer. 2007, 14, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.; Terörde, K.; Dörr, H.G.; Trollmann, R. Recombinant human growth hormone activates neuroprotective growth factors in hypoxic brain injury in neonatal mice. Endocrinology 2021, 162, bqab008. [Google Scholar] [CrossRef] [PubMed]
- Lyons, R.A.; Saridogan, E.; Djahanbakhch, O. The reproductive significance of human Fallopian tube cilia. Hum. Reprod. Update 2006, 12, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Von Wolff, M.; Schneider, S.; Kollmann, Z.; Weiss, B.; Bersinger, N.A. Exogenous gonadotropins do not increase the blood-follicular transportation capacity of extra-ovarian hormones such as prolactin and cortisol. Reprod. Biol. Endocrinol. 2013, 11, 87. [Google Scholar] [CrossRef] [Green Version]
- Artini, P.G.; Tatone, C.; Sperduti, S.; D’Aurora, M.; Franchi, S.; Di Emidio, G.; Ciriminna, R.; Vento, M.; Di Pietro, C.; Stuppia, L.; et al. Cumulus cells surrounding oocytes with high developmental competence exhibit down-regulation of phosphoinositol 1,3 kinase/protein kinase B (PI3K/AKT) signalling genes involved in proliferation and survival. Hum. Reprod. 2017, 32, 2474–2484. [Google Scholar] [CrossRef]
- Tarkowski, A.K. An air-drying method for chromosome preparations from mouse eggs. Cytogenetics 1966, 5, 394–400. [Google Scholar] [CrossRef]
- Kuzmina, T.I.; Lebedeva, I.Y.; Torner, H.; Alm, H.; Denisenko, V.Y. Effects of prolactin on intracellular stored calcium in the course of bovine oocyte maturation in vitro. Theriogenology 1999, 51, 1363–1374. [Google Scholar] [CrossRef]
- Homa, S.T. Effects of cyclic AMP on the spontaneous meiotic maturation of cumulus-free bovine oocytes cultured in chemically defined medium. J. Exp. Zool. 1988, 248, 222–231. [Google Scholar] [CrossRef]
- Parrish, J.J.; Susko-Parrish, J.L.; Leibfried-Rutledge, M.L.; Critser, E.S.; Eyestone, W.H.; First, N.L. Bovine in vitro fertilization with frozen-thawed semen. Theriogenology 1986, 25, 591–600. [Google Scholar] [CrossRef]
- Rosenkrans, C.F., Jr.; First, N.L. Effect of free amino acids and vitamins on cleavage and developmental rate of bovine zygotes in vitro. J. Anim. Sci. 1994, 72, 434–437. [Google Scholar] [CrossRef]
- Paula-Lopes, F.F.; Boelhauve, M.; Habermann, F.A.; Sinowatz, F.; Wolf, E. Leptin promotes meiotic progression and developmental capacity of bovine oocytes via cumulus cell-independent and -dependent mechanisms. Biol. Reprod. 2007, 76, 532–541. [Google Scholar] [CrossRef] [Green Version]
- Min, S.H.; Song, B.S.; Yeon, J.Y.; Kim, J.W.; Bae, J.H.; Park, S.Y.; Lee, Y.H.; Chang, K.T.; Koo, D.B. A cathepsin B inhibitor, E-64, improves the preimplantation development of bovine somatic cell nuclear transfer embryos. J. Reprod. Dev. 2014, 60, 21–27. [Google Scholar] [CrossRef] [Green Version]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singina, G.N.; Shedova, E.N.; Lopukhov, A.V.; Mityashova, O.S.; Lebedeva, I.Y. Delaying Effects of Prolactin and Growth Hormone on Aging Processes in Bovine Oocytes Matured In Vitro. Pharmaceuticals 2021, 14, 684. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14070684
Singina GN, Shedova EN, Lopukhov AV, Mityashova OS, Lebedeva IY. Delaying Effects of Prolactin and Growth Hormone on Aging Processes in Bovine Oocytes Matured In Vitro. Pharmaceuticals. 2021; 14(7):684. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14070684
Chicago/Turabian StyleSingina, Galina N., Ekaterina N. Shedova, Alexander V. Lopukhov, Olga S. Mityashova, and Irina Y. Lebedeva. 2021. "Delaying Effects of Prolactin and Growth Hormone on Aging Processes in Bovine Oocytes Matured In Vitro" Pharmaceuticals 14, no. 7: 684. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14070684