
Dickkopf Proteins and Their Role in Cancer: A Family of Wnt Antagonists with a Dual Role

 , , ,
, , ,  ,
,
Abstract
:1. Overview of the Wnt Pathway
2. The Dickkopf (DKK) Family and Its Role in Cancer
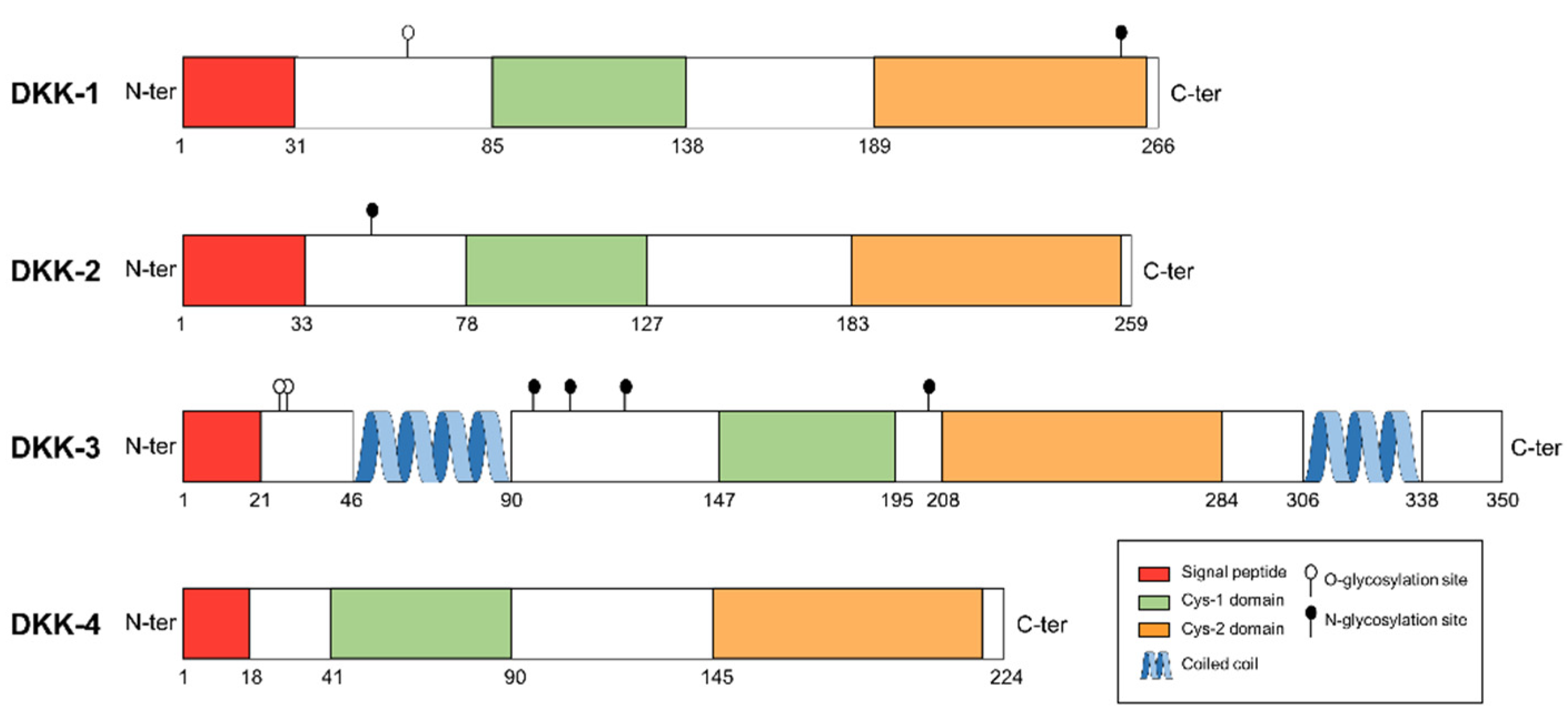
2.1. Dickkopf-1 (DKK-1)
2.2. Dickkopf-2 (DKK-2)
2.3. Dickkopf-3 (DKK-3)
2.4. Dickkopf-4 (DKK-4)
3. DKK Proteins as Diagnostic and/or Prognostic Factor in Cancer
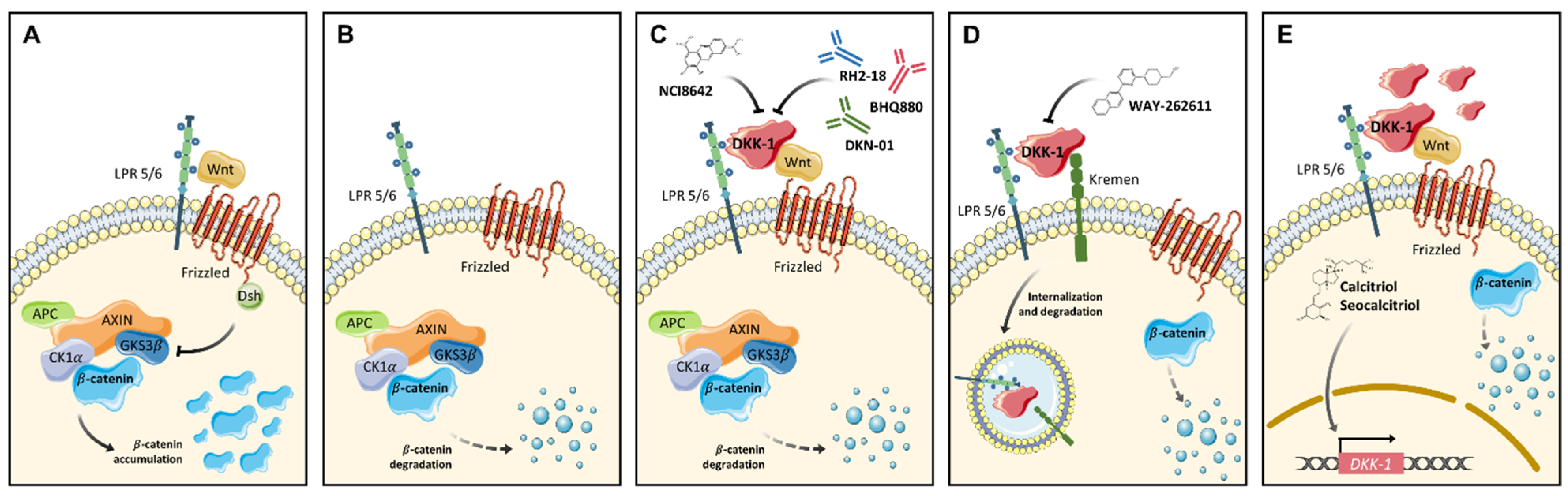
4. DKK Family as Emerging Molecular Targets for Anti-Cancer Drugs
4.1. Small Molecules
4.2. Monoclonal Antibodies
4.3. Gene and Immunotherapy
4.4. Activators of Wnt Antagonist Proteins
5. Clinical Trials for Wnt Antagonist Proteins in Cancer
6. Final Remarks
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiese, K.E.; Nusse, R.; van Amerongen, R. Wnt signalling: Conquering complexity. Development 2018, 145, dev165902. [Google Scholar] [CrossRef] [Green Version]
- El-Sahli, S.; Xie, Y.; Wang, L.; Liu, S. Wnt signaling in cancer metabolism and immunity. Cancers 2019, 11, 904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, L.F.; Kaur, P.; Bunnag, N.; Suresh, J.; Sung, I.C.H.; Tan, Q.H.; Gruber, J.; Tolwinski, N.S. WNT Signaling in Disease. Cells 2019, 8, 826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, S.; Hua, F.; Hu, Z.W. The regulation of β-catenin activity and function in cancer: Therapeutic opportunities. Oncotarget 2017, 8, 33972–33989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadigan, K.M.; Ramakrishnan, A.B. Wnt target genes and where to find them. F1000Research 2017, 6, 746. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, S.Y.; Jun, Y.; Kim, J.Y.; Nam, J.S. Roles of Wnt target genes in the journey of cancer stem cells. Int. J. Mol. Sci. 2017, 18, 1604. [Google Scholar] [CrossRef] [Green Version]
- De, A. Wnt/Ca 2 signaling pathway: A brief overview. Acta Biochim. Biophys. Sin. 2011, 43, 745–756. [Google Scholar] [CrossRef]
- Wang, M.; Marco, P.D.; Capra, V.; Kibar, Z. Update on the Role of the Non-Canonical Wnt/Planar Cell Polarity Pathway in Neural Tube Defects. Cells 2019, 8, 1198. [Google Scholar] [CrossRef] [Green Version]
- Haseeb, M.; Pirzada, R.H.; Ain, Q.U.; Choi, S. Wnt Signaling in the Regulation of Immune Cell and Cancer Therapeutics. Cells 2019, 8, 1380. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Nakamura, T.; Matsumoto, K. The functions and possible significance of Kremen as the gatekeeper of Wnt signalling in development and pathology: Molecular Medicine. J. Cell. Mol. Med. 2008, 12, 391–408. [Google Scholar] [CrossRef]
- Hartung, N.; Benary, U.; Wolf, J.; Kofahl, B. Paracrine and autocrine regulation of gene expression by Wnt-inhibitor Dickkopf in wild-type and mutant hepatocytes. BMC Syst. Biol. 2017, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Niehrs, C. Function and biological roles of the Dickkopf family of Wnt modulators. Oncogene 2006, 25, 7469–7481. [Google Scholar] [CrossRef] [Green Version]
- Pandur, P.; Läsche, M.; Eisenberg, L.M.; Kühl, M. Wnt-11 activation of a non-canonical Wnt signalling pathway is required for cardiogenesis. Nature 2002, 418, 636–641. [Google Scholar] [CrossRef]
- Rawson, J.B.; Manno, M.; Mrkonjic, M.; Daftary, D.; Dicks, E.; Buchanan, D.D.; Younghusband, H.B.; Parfrey, P.S.; Young, J.P.; Pollett, A.; et al. Promoter methylation of Wnt antagonists DKK1 and SFRP1 is associated with opposing tumor subtypes in two large populations of colorectal cancer patients. Carcinogenesis 2011, 32, 741–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.C.; Zhang, X.L.; Yang, B.; Geng, J.; Peng, B.; Zheng, J.H. Decreased expression of Dkk1 and Dkk3 in human clear cell renal cell carcinoma. Mol. Med. Rep. 2014, 9, 2367–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; He, H.; Lv, R.; Zhang, M.; Huang, H.; An, Z.; Li, S. Preliminary mechanism on the methylation modification of Dkk-1 and Dkk-3 in hepatocellular carcinoma. Tumor Biol. 2015, 36, 1245–1250. [Google Scholar] [CrossRef]
- Sato, H.; Suzuki, H.; Toyota, M.; Nojima, M.; Maruyama, R.; Sasaki, S.; Takagi, H.; Sogabe, Y.; Sasaki, Y.; Idogawa, M.; et al. Frequent epigenetic inactivation of DICKKOPF family genes in human gastrointestinal tumors. Carcinogenesis 2007, 28, 2459–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.A.; Yoo, S.H.; Lee, H.H.; Sun, D.S.; Won, H.S.; Kim, O.; Ko, Y.H. Prognostic value of Dickkopf-1 and ß-catenin expression in advanced gastric cancer. BMC Cancer 2018, 18, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rachner, T.D.; Thiele, S.; Göbel, A.; Browne, A.; Fuessel, S.; Erdmann, K.; Wirth, M.P.; Fröhner, M.; Todenhöfer, T.; Muders, M.H.; et al. High serum levels of Dickkopf-1 are associated with a poor prognosis in prostate cancer patients. BMC Cancer 2014, 14, 649. [Google Scholar] [CrossRef]
- Aguilera, Ó.; González-Sancho, J.M.; Zazo, S.; Rincón, R.; Fernández, A.F.; Tapia, O.; Canals, F.; Morte, B.; Calvanese, V.; Orgaz, J.L.; et al. Nuclear DICKKOPF-1 as a biomarker of chemoresistance and poor clinical outcome in colorectal cancer. Oncotarget 2015, 6, 5903–5917. [Google Scholar] [CrossRef] [Green Version]
- Erdal, H.; Utku, Ö.G.; Karatay, E.; Çelik, B.; Elbeg, Ş.; Doǧan, I. Combination of DKK1 and AFP improves diagnostic accuracy of hepatocellular carcinoma compared with either marker alone. Turk. J. Gastroenterol. 2016, 27, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Glinka, A.; Wu, W.; Delius, H.; Monaghan, A.P.; Blumenstock, C.; Niehrs, C. Dickkopf-1 is a member of a new family of secreted proteins and functions in head induction. Nature 1998, 391, 357–362. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Adamska, M.; Meisler, M.H. Hypomorphic expression of Dkk1 in the doublebridge mouse: Dose dependence and compensatory interactions with Lrp6. Development 2004, 131, 2543–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bafico, A.; Liu, G.; Yaniv, A.; Gazit, A.; Aaronson, S.A. Novel mechanism of Wnt signalling inhibition mediated by Dickkopf-1 interaction with LRP6/Arrow. Nat. Cell Biol. 2001, 3, 683–686. [Google Scholar] [CrossRef] [PubMed]
- Matoba, K.; Mihara, E.; Tamura-Kawakami, K.; Miyazaki, N.; Maeda, S.; Hirai, H.; Thompson, S.; Iwasaki, K.; Takagi, J. Conformational Freedom of the LRP6 Ectodomain Is Regulated by N-glycosylation and the Binding of the Wnt Antagonist Dkk1. Cell Rep. 2017, 18, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Baetta, R.; Banfi, C. Dkk (Dickkopf) proteins emerging new players in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1330–1342. [Google Scholar] [CrossRef]
- Shi, L.S.; Huang, G.; Yu, B.; Wen, X.Q. Clinical significance and prognostic value of serum Dickkopf-1 concentrations in patients with lung cancer. Clin. Chem. 2009, 55, 1656–1664. [Google Scholar] [CrossRef]
- Qiao, R.; Zhong, R.; Chang, Q.; Teng, J.; Pei, J.; Han, B.; Chu, T. Serum dickkopf-1 as a clinical and prognostic factor in nonsmall cell lung cancer patients with bone metastases. Oncotarget 2017, 8, 79469–79479. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhou, H.; Zhang, X.; Ma, X.; Liu, Z.; Liu, X. Elevated levels of dickkopf-1 are associated with β-catenin accumulation and poor prognosis in patients with chondrosarcoma. PLoS ONE 2014, 9, e105414. [Google Scholar] [CrossRef]
- Zarea, M.; Mohammadian Bajgiran, A.; Sedaghati, F.; Hatami, N.; Taheriazam, A.; Yahaghi, E.; Shakeri, M. Diagnostic investigations of DKK-1 and PDCD5 expression levels as independent prognostic markers of human chondrosarcoma. IUBMB Life 2016, 68, 597–601. [Google Scholar] [CrossRef] [Green Version]
- Koppen, A.; Ait-Aissa, R.; Hopman, S.; Koster, J.; Haneveld, F.; Versteeg, R.; Valentijn, L.J. Dickkopf-1 is down-regulated by MYCN and inhibits neuroblastoma cell proliferation. Cancer Lett. 2007, 256, 218–228. [Google Scholar] [CrossRef]
- Granchi, D.; Corrias, M.V.; Garaventa, A.; Baglìo, S.R.; Cangemi, G.; Carlini, B.; Paolucci, P.; Giunti, A.; Baldini, N. Neuroblastoma and bone metastases: Clinical significance and prognostic value of Dickkopf 1 plasma levels. Bone 2011, 48, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Smolarz, A.J.; Olson, S.; David, O.; Reiser, J.; Kutner, R.; Daw, N.C.; Prockop, D.J.; Horwitz, E.M.; Gregory, C.A. A potential role for Dkk-1 in the pathogenesis of osteosarcoma predicts novel diagnostic and treatment strategies. Br. J. Cancer 2007, 97, 1552–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, S.D.; Trucco, M.; Guzman, W.B.; Hayashi, M.; Loeb, D.M. A monoclonal antibody against the Wnt signaling inhibitor dickkopf-1 inhibits osteosarcoma metastasis in a preclinical model. Oncotarget 2016, 7, 21114–21123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, D.; Agra, N.; Pestaña, Á.; Alonso, J.; González-Sancho, J.M. The EWS/FLI1 oncogenic protein inhibits expression of the Wnt inhibitor DICKKOPF-1 gene and antagonizes β-catenin/TCF-mediated transcription. Carcinogenesis 2010, 31, 394–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaghan, A.P.; Kioschis, P.; Wu, W.; Zuniga, A.; Bock, D.; Poustka, A.; Delius, H.; Niehrs, C. Dickkopf genes are co-ordinately expressed in mesodermal lineages. Mech. Dev. 1999, 87, 45–56. [Google Scholar] [CrossRef]
- Mao, B.; Wu, W.; Li, Y.; Hoppe, D.; Stannek, P.; Glinka, A.; Niehrs, C. LDL-receptor-related protein 6 is a receptor for Dickkopf proteins. Nature 2001, 411, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Glinka, A.; Delius, H.; Niehrs, C. Mutual antagonism between dickkopf1 and dickkopf2 regulates Wnt/β-catenin signalling. Curr. Biol. 2000, 10, 1611–1614. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Q.; Wu, J.; Wang, W.J.; Chen, S.; Zheng, Y.; Yu, X.; Meeth, K.; Sahraei, M.; Bothwell, A.L.M.; Chen, L.; et al. DKK2 imparts tumor immunity evasion through β-catenin-independent suppression of cytotoxic immune-cell activation. Nat. Med. 2018, 24, 262–270. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.-F.; Liu, J.; Deng, Z.-G.; Zeng, L.; Zhou, W.-B. Long Noncoding RNA GAS5 Targeting miR-221-3p/Cyclin-Dependent Kinase Inhibitor 2B Axis Regulates Follicular Thyroid Carcinoma Cell Cycle and Proliferation. Pathobiology 2021, 88, 289–300. [Google Scholar] [CrossRef]
- Shin, J.H.; Jeong, J.; Choi, J.; Lim, J.; Dinesh, R.K.; Braverman, J.; Hong, J.Y.; Maher, S.E.; Amezcua Vesely, M.C.; Kim, W.J.; et al. Dickkopf-2 regulates the stem cell marker LGR5 in colorectal cancer via HNF4α1. Iscience 2021, 24, 102411. [Google Scholar] [CrossRef]
- Fraungruber, P.; Kaltofen, T.; Heublein, S.; Kuhn, C.; Mayr, D.; Burges, A.; Mahner, S.; Rathert, P.; Jeschke, U.; Trillsch, F. G Protein-Coupled Estrogen Receptor Correlates With Dkk2 Expression and Has Prognostic Impact in Ovarian Cancer Patients. Front. Endocrinol. 2021, 12, 18. [Google Scholar] [CrossRef]
- Lee, H.M.; Kwon, S.B.; Son, A.; Kim, D.H.; Kim, K.H.; Lim, J.; Kwon, Y.G.; Kang, J.S.; Lee, B.K.; Byun, Y.H.; et al. Stabilization of intrinsically disordered DKK2 protein by fusion to RNA-binding domain. Int. J. Mol. Sci. 2019, 20, 2847. [Google Scholar] [CrossRef] [Green Version]
- Sopko, R.; Mugford, J.W.; Lehmann, A.; Shapiro, R.I.; Rushe, M.; Kulkarni, A.; Worrall, J.; Amatucci, J.; Wen, D.; Pederson, N.E.; et al. Engineering potent long-acting variants of the Wnt inhibitor DKK2. Protein Eng. Des. Sel. 2017, 30, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Shen, T.; Chen, Z.; Qiao, J.; Sun, X.; Xiao, Q. Neutralizing monoclonal antibody against Dickkopf2 impairs lung cancer progression via activating NK cells. Cell Death Discov. 2019, 5, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinho, S.; Niehrs, C. Dkk3 is required for TGF-β signaling during Xenopus mesoderm induction. Differentiation 2007, 75, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Krupnik, V.E.; Sharp, J.D.; Jiang, C.; Robison, K.; Chickering, T.W.; Amaravadi, L.; Brown, D.E.; Guyot, D.; Mays, G.; Leiby, K.; et al. Functional and structural diversity of the human Dickkopf gene family. Gene 1999, 238, 301–313. [Google Scholar] [CrossRef]
- Mao, B.; Wu, W.; Davidson, G.; Marhold, J.; Li, M.; Mechler, B.M.; Dellus, H.; Hoppe, D.; Stannek, P.; Walter, C.; et al. Kremen proteins are Dickkopf receptors that regulate Wnt/β-catenin signalling. Nature 2002, 417, 664–667. [Google Scholar] [CrossRef]
- Yue, W.; Sun, Q.; Dacic, S.; Landreneau, R.J.; Siegfried, J.M.; Yu, J.; Zhang, L. Downregulation of Dkk3 activates β-catenin/TCF-4 signaling in lung cancer. Carcinogenesis 2008, 29, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Hoang, B.H.; Kubo, T.; Healey, J.H.; Yang, R.; Nathan, S.S.; Kolb, E.A.; Mazza, B.A.; Meyers, P.A.; Gorlick, R. Dickkopf 3 Inhibits Invasion and Motility of Saos-2 Osteosarcoma Cells by Modulating the Wnt-β-Catenin Pathway. Cancer Res. 2004, 64, 2734–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eun, J.L.; Bo, B.L.; Han, J.; Eun, Y.C.; Young, M.S.; Park, J.; Kim, D.H. CpG island hypermethylation of E-cadherin (CDH1) and integrin α4 is associated with recurrence of early stage esophageal squamous cell carcinoma. Int. J. Cancer 2008, 123, 2073–2079. [Google Scholar] [CrossRef]
- Tsuji, T.; Nozaki, I.; Miyazaki, M.; Sakaguchi, M.; Pu, H.; Hamazaki, Y.; Iijima, O.; Namba, M. Antiproliferative activity of REIC/Dkk-3 and its significant down-regulation in non-small-cell lung carcinomas. Biochem. Biophys. Res. Commun. 2001, 289, 257–263. [Google Scholar] [CrossRef]
- Abarzua, F.; Sakaguchi, M.; Takaishi, M.; Nasu, Y.; Kurose, K.; Ebara, S.; Miyazaki, M.; Namba, M.; Kumon, H.; Huh, N.H. Adenovirus-mediated overexpression of REIC/Dkk-3 selectively induces apoptosis in human prostate cancer cells through activation of c-Jun-NH 2-kinase. Cancer Res. 2005, 65, 9617–9622. [Google Scholar] [CrossRef] [Green Version]
- Tanimoto, R.; Abarzua, F.; Sakaguchi, M.; Takaishi, M.; Nasu, Y.; Kumon, H.; Huh, N.H. REIC/Dkk-3 as a potential gene therapeutic agent against human testicular cancer. Int. J. Mol. Med. 2007, 19, 363–368. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Kataoka, K.; Abarzua, F.; Tanimoto, R.; Watanabe, M.; Murata, H.; Than, S.S.; Kurose, K.; Kashiwakura, Y.; Ochiai, K.; et al. Overexpression of REIC/Dkk-3 in normal fibroblasts suppresses tumor growth via induction of interleukin-7. J. Biol. Chem. 2009, 284, 14236–14244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, K.; Hirata, H.; Majid, S.; Chen, Y.; Zaman, M.S.; Tabatabai, Z.L.; Hinoda, Y.; Dahiya, R. Wnt antagonist DICKKOPF-3 (Dkk-3) induces apoptosis in human renal cell carcinoma. Mol. Carcinog. 2011, 50, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Mizobuchi, Y.; Matsuzaki, K.; Kuwayama, K.; Kitazato, K.; Mure, H.; Kageji, T.; Nagahiro, S. REIC/Dkk-3 induces cell death in human malignant glioma. Neuro. Oncol. 2008, 10, 244–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, K.; Watanabe, M.; Sakaguchi, M.; Ogasawara, Y.; Ochiai, K.; Nasu, Y.; Doihara, H.; Kashiwakura, Y.; Huh, N.H.; Kumon, H.; et al. REIC/Dkk-3 overexpression downregulates P-glycoprotein in multidrug-resistant MCF7/ADR cells and induces apoptosis in breast cancer. Cancer Gene Ther. 2009, 16, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Fujii, M.; Katase, N.; Lefeuvre, M.; Gunduz, M.; Buery, R.R.; Tamamura, R.; Tsujigiwa, H.; Nagatsuka, H. Dickkopf (Dkk)-3 and β-catenin expressions increased in the transition from normal oral mucosal to oral squamous cell carcinoma. J. Mol. Histol. 2011, 42, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Meister, M.; Papatriantafyllou, M.; Nordström, V.; Kumar, V.; Ludwig, J.; Lui, K.O.; Boyd, A.S.; Popovic, Z.V.; Fleming, T.H.; Moldenhauer, G.; et al. Dickkopf-3, a tissue-derived modulator of local T-cell responses. Front. Immunol. 2015, 6, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, N.; Ranftl, R.; Chicherova, I.; Slaven, N.D.; Moeendarbary, E.; Farrugia, A.J.; Lam, M.; Semiannikova, M.; Westergaard, M.C.W.; Tchou, J.; et al. Dickkopf-3 links HSF1 and YAP/TAZ signalling to control aggressive behaviours in cancer-associated fibroblasts. Nat. Commun. 2019, 10, 130. [Google Scholar] [CrossRef]
- Zitt, M.; Untergasser, G.; Amberger, A.; Moser, P.; Stadlmann, S.; Zitt, M.; Müller, H.M.; Mühlmann, G.; Perathoner, A.; Margreiter, R.; et al. Dickkopf-3 as a new potential marker for neoangiogenesis in colorectal cancer: Expression in cancer tissue and adjacent non-cancerous tissue. Dis. Markers 2008, 24, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, G.; Steurer, M.; Zimmermann, M.; Hermann, M.; Kern, J.; Amberger, A.; Gastl, G.; Gunsilius, E. The Dickkopf-homolog 3 is expressed in tumor endothelial cells and supports capillary formation. Int. J. Cancer 2008, 122, 1539–1547. [Google Scholar] [CrossRef]
- Zenzmaier, C.; Untergasser, G.; Hermann, M.; Dirnhofer, S.; Sampson, N.; Berger, P. Dysregulation of Dkk-3 expression in benign and malignant prostatic tissue. Prostate 2008, 68, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Bazzi, H.; Fantauzzo, K.A.; Richardson, G.D.; Jahoda, C.A.B.; Christiano, A.M. The Wnt inhibitor, Dickkopf 4, is induced by canonical Wnt signaling during ectodermal appendage morphogenesis. Dev. Biol. 2007, 305, 498–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pendás-Franco, N.; García, J.M.; Peña, C.; Valle, N.; Pálmer, H.G.; Heinäniemi, M.; Carlberg, C.; Jiménez, B.; Bonilla, F.; Muñoz, A.; et al. DICKKOPF-4 is induced by TCF/β-catenin and upregulated in human colon cancer, promotes tumour cell invasion and angiogenesis and is repressed by 1α,25-dihydroxyvitamin D3. Oncogene 2008, 27, 4467–4477. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Yao, Z.; Li, L.; Huang, J. Role of DKK4 in tumorigenesis and tumor progression. Int. J. Biol. Sci. 2018, 14, 616–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatima, S.; Lee, N.P.; Tsang, F.H.; Kolligs, F.T.; Ng, I.O.L.; Poon, R.T.P.; Fan, S.T.; Luk, J.M. Dickkopf 4 (DKK4) acts on Wnt/Β-catenin pathway by influencing Β-catenin in hepatocellular carcinoma. Oncogene 2012, 31, 4233–4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, H.; Hinoda, Y.; Majid, S.; Chen, Y.; Zaman, M.S.; Ueno, K.; Nakajima, K.; Tabatabai, Z.L.; Ishii, N.; Dahiya, R. DICKKOPF-4 activates the noncanonical c-Jun-NH2 kinase signaling pathway while inhibiting the Wnt-canonical pathway in human renal cell carcinoma. Cancer 2011, 117, 1649–1660. [Google Scholar] [CrossRef]
- Ebert, M.P.A.; Tänzer, M.; Balluff, B.; Burgermeister, E.; Kretzschmar, A.K.; Hughes, D.J.; Tetzner, R.; Lofton-Day, C.; Rosenberg, R.; Reinacher-Schick, A.C.; et al. TFAP2E–DKK4 and Chemoresistance in Colorectal Cancer. N. Engl. J. Med. 2012, 366, 44–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Shen, J.; Hu, N.; Xu, X.; Li, J. DKK4 enhances resistance to chemotherapeutics 5-Fu and YN968D1 in colorectal cancer cells. Oncol. Lett. 2017, 13, 587–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Liu, Y.; Li, W.; Li, A.; Sun, Q. DKK4-knockdown enhances chemosensitivity of A549/DTX cells to docetaxel. Acta Biochim. Biophys. Sin. 2017, 49, 899–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eskander, R.N.; Ali, S.; Dellinger, T.; Lankes, H.A.; Randall, L.M.; Ramirez, N.C.; Monk, B.J.; Walker, J.L.; Eisenhauer, E.; Hoang, B.H. Expression Patterns of the Wnt Pathway Inhibitors Dickkopf3 and Secreted Frizzled-Related Proteins 1 and 4 in Endometrial Endometrioid Adenocarcinoma: An NRG Oncology/Gynecologic Oncology Group Study. Int. J. Gynecol. Cancer 2016, 26, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Mabille, C.; Ruyssen-Witrand, A.; Degboe, Y.; Gennero, I.; Loiseau, H.A.; Roussel, M.; Hebraud, B.; Nigon, D.; Attal, M.; Laroche, M. DKK1 and sclerostin are early markers of relapse in multiple myeloma. Bone 2018, 113, 114–117. [Google Scholar] [CrossRef]
- Hall, C.L.; Daignault, S.D.; Shah, R.B.; Pienta, K.J.; Keller, E.T. Dickkopf-1 expression increases early in prostate cancer development and decreases during progression from primary tumor to metastasis. Prostate 2008, 68, 1396–1404. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Chen, G.D.; Fang, F.; Liu, Z.; Lau, S.H.Y.; Zhang, J.F.; Lau, W.Y.; Yang, L.Y. Dickkopf-1: As a diagnostic and prognostic serum marker for early hepatocellular carcinoma. Int. J. Biol. Markers 2013, 28, 286–297. [Google Scholar] [CrossRef]
- Al Shareef, Z.; Kardooni, H.; Murillo-Garzón, V.; Domenici, G.; Stylianakis, E.; Steel, J.H.; Rabano, M.; Gorroño-Etxebarria, I.; Zabalza, I.; Vivanco, M.M.; et al. Protective effect of stromal Dickkopf-3 in prostate cancer: Opposing roles for TGFBI and ECM-1. Oncogene 2018, 37, 5305–5324. [Google Scholar] [CrossRef] [Green Version]
- Kamińska, K.; Białkowska, A.; Kowalewski, J.; Huang, S.; Lewandowska, M.A. Differential gene methylation patterns in cancerous and non-cancerous cells. Oncol. Rep. 2019, 42, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Zenzmaier, C.; Heitz, M.; Klocker, H.; Buck, M.; Gardiner, R.A.; Berger, P. Elevated levels of Dickkopf-related protein 3 in seminal plasma of prostate cancer patients. J. Transl. Med. 2011, 9, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.T.; Ma, Y.Y.; Guo, K.; Xia, Y.J.; Wang, H.J.; Li, L.; He, X.J.; Huang, D.S.; Tao, H.Q. Assessing the diagnostic value of serum dickkopf-related protein 1 levels in cancer detection: A case-control study and meta-analysis. Asian Pac. J. Cancer Prev. 2014, 15, 9077–9083. [Google Scholar] [CrossRef] [Green Version]
- Liang, B.; Zhong, L.; He, Q.; Wang, S.; Pan, Z.; Wang, T.; Zhao, Y. Serum dickkopf-1 as a biomarker in screening gastrointestinal cancers: A systematic review and meta-analysis. Onco. Targets. Ther. 2015, 8, 3115–3122. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.R.; Li, Y.F.; Deng, Z.Q.; Cao, J.Q. Prognostic Significance of Dickkopf-1 in Gastric Cancer Survival: A Meta-Analysis. Genet. Test. Mol. Biomark. 2016, 20, 170–175. [Google Scholar] [CrossRef]
- Li, Z.; Mou, L.; Gao, H.; Zeng, Y.; Tang, X.; Deng, X.; Pu, Z.; Ni, Y.; Zhan, Y.; Rajasekharan, C. Diagnostic accuracy of serum dickkopf-1 protein in diagnosis hepatocellular carcinoma: An updated meta-analysis. Medicine 2019, 98, e16725. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, Y.; Yang, Q. Sensitivity and specificity of Dickkopf-1 protein in serum for diagnosing hepatocellular carcinoma: A meta-analysis. Int. J. Biol. Markers 2014, 29, e403–e410. [Google Scholar] [CrossRef]
- Chen, G.Y.; Zheng, H.C. The clinicopathological and prognostic significances of Dkk3 expression in cancers: A bioinformatics analysis. Cancer Biomark. 2018, 23, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, S.; Zhou, L.; Yu, L.; Zhang, L.-M. A network-pathway based module identification for predicting the prognosis of ovarian cancer patients. J. Ovarian Res. 2016, 9, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Park, K.H.; Shin, S.J.; Lee, K.Y.; Kim, T.I.; Kim, N.K.; Rha, S.Y.; Ahn, J.B. CpG island methylator phenotype and methylation of wnt pathway genes together predict survival in patients with colorectal cancer. Yonsei Med. J. 2018, 59, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Jee, B.A.; Choi, J.H.; Rhee, H.; Yoon, S.; Kwon, S.M.; Nahm, J.H.; Yoo, J.E.; Jeon, Y.; Choi, G.H.; Woo, H.G.; et al. Dynamics of genomic, epigenomic, and transcriptomic aberrations during stepwise hepatocarcinogenesis. Cancer Res. 2019, 79, 5500–5512. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, J.C.; Lundquist IV, J.T.; Gilbert, A.M.; Alon, N.; Bex, F.J.; Bhat, B.M.; Bursavich, M.G.; Coleburn, V.E.; Felix, L.A.; Green, D.M.; et al. (1-(4-(Naphthalen-2-yl)pyrimidin-2-yl)piperidin-4-yl)methanamine: A wingless β-catenin agonist that increases bone formation rate. J. Med. Chem. 2009, 52, 6962–6965. [Google Scholar] [CrossRef]
- Iozzi, S.; Remelli, R.; Lelli, B.; Diamanti, D.; Pileri, S.; Bracci, L.; Roncarati, R.; Caricasole, A.; Bernocco, S. Functional Characterization of a Small-Molecule Inhibitor of the DKK1-LRP6 Interaction. ISRN Mol. Biol. 2012, 2012, 823875. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Yang, X.; Pan, Y.; Qi, Q.; Shen, J.; Fang, H.; Ji, Z. L-securinine inhibits the proliferation of A549 lung cancer cellsand promotes DKK1 promoter methylation. Oncol. Lett. 2017, 14, 4243–4248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glantschnig, H.; Hampton, R.A.; Lu, P.; Zhao, J.Z.; Vitelli, S.; Huang, L.; Haytko, P.; Cusick, T.; Ireland, C.; Jarantow, S.W.; et al. Generation and selection of novel fully human monoclonal antibodies that neutralize Dickkopf-1 (DKK1) inhibitory function in vitro and increase bone mass in vivo. J. Biol. Chem. 2010, 285, 40135–40147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ettenberg, S.; Cong, F.; Shulok, J.; Urlinger, S.; Bardroff, M.; Charlat, O.; Mishina, Y.; Liu, S.; Daley, M.; Li, S.; et al. BHQ880, a novel anti-DKK1 neutralizing antibody, inhibits tumor-induced osteolytic bone disease. Cancer Res. 2008, 68, 3987. [Google Scholar]
- Kagey, M.H.; Wu, Y.; Zhang, X.; Sirad, C.A.; Mulligan, S.E.; He, X.; Mirabelli, C.K. Abstract 369: Therapeutic targeting of the Wnt antagonist DKK1 with a humanized monoclonal antibody in oncology indications. Cancer Res. 2017, 77, 369. [Google Scholar] [CrossRef]
- Aguilera, O.; Peña, C.; García, J.M.; Larriba, M.J.; Ordóñez-morán, P.; Navarro, D.; Barbáchano, A.; De Silanes, I.L.; Ballestar, E.; Fraga, M.F.; et al. The Wnt antagonist DICKKOPF-1 gene is induced by 1α,25-dihydroxyvitamin D3 associated to the differentiation of human colon cancer cells. Carcinogenesis 2007, 28, 1877–1884. [Google Scholar] [CrossRef] [PubMed]
- Enochson, L.; Stenberg, J.; Brittberg, M.; Lindahl, A. GDF5 reduces MMP13 expression in human chondrocytes via DKK1 mediated canonical Wnt signaling inhibition. Osteoarthr. Cartil. 2014, 22, 566–577. [Google Scholar] [CrossRef] [Green Version]
- Nierste, B.A.; Glackin, C.A.; Kirshner, J. Dkk-1 and IL-7 in plasma of patients with multiple myeloma prevent differentiation of mesenchymal stem cells into osteoblasts. Am. J. Blood Res. 2014, 4, 73–85. [Google Scholar]
- Choe, J.Y.; Kim, J.H.; Park, K.Y.; Choi, C.H.; Kim, S.K. Activation of dickkopf-1 and focal adhesion kinase pathway by tumour necrosis factor a induces enhanced migration of fibroblast-like synoviocytes in rheumatoid arthritis. Rheumatology 2016, 55, 928–938. [Google Scholar] [CrossRef] [Green Version]
- Chae, W.J.; Ehrlich, A.K.; Chan, P.Y.; Teixeira, A.M.; Henegariu, O.; Hao, L.; Shin, J.H.; Park, J.H.; Tang, W.H.; Kim, S.T.; et al. The Wnt Antagonist Dickkopf-Promotes Pathological Type 2 Cell-Mediated Inflammation. Immunity 2016, 44, 246–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Yi, Q.; Tang, L. The roles of nuclear focal adhesion kinase (FAK) on Cancer: A focused review. J. Exp. Clin. Cancer Res. 2019, 38, 250. [Google Scholar] [CrossRef] [Green Version]
- Priestley, R.S.; Cheung, J.; Murphy, E.J.; Ehebauer, M.T.; Davis, J.B.; Di Daniel, E. A novel high-content imaging-based technique for measuring binding of Dickkopf-1 to low-density lipoprotein receptor-related protein 6. J. Pharmacol. Toxicol. Methods 2019, 95, 47–55. [Google Scholar] [CrossRef]
- Li, X.; Shan, J.; Chang, W.; Kim, I.; Bao, J.; Lee, H.J.; Zhang, X.; Samuel, V.T.; Shulman, G.I.; Liu, D.; et al. Chemical and genetic evidence for the involvement of Wnt antagonist Dickkopf2 in regulation of glucose metabolism. Proc. Natl. Acad. Sci. USA 2012, 109, 11402–11407. [Google Scholar] [CrossRef] [Green Version]
- Thysiadis, S.; Katsamakas, S.; Mpousis, S.; Avramidis, N.; Efthimiopoulos, S.; Sarli, V. Design and synthesis of gallocyanine inhibitors of DKK1/LRP6 interactions for treatment of Alzheimer’s disease. Bioorg. Chem. 2018, 80, 230–244. [Google Scholar] [CrossRef]
- Pozzi, S.; Fulciniti, M.; Yan, H.; Vallet, S.; Eda, H.; Patel, K.; Santo, L.; Cirstea, D.; Hideshima, T.; Schirtzinge, L.; et al. In vivo and in vitro effects of a novel anti-Dkk1 neutralizing antibody in multiple myeloma. Bone 2013, 53, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Fulciniti, M.; Tassone, P.; Hideshima, T.; Vallet, S.; Nanjappa, P.; Ettenberg, S.A.; Shen, Z.; Patel, N.; Tai, Y.T.; Chauhan, D.; et al. Anti-DKK1 mAb (BHQ880) as a potential therapeutic agent for multiple myeloma. Blood 2009, 114, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Glantschnig, H.; Scott, K.; Hampton, R.; Wei, N.; McCracken, P.; Nantermet, P.; Zhao, J.Z.; Vitelli, S.; Huang, L.; Haytko, P.; et al. A rate-limiting role for Dickkopf-1 in bone formation and the remediation of bone loss in mouse and primate models of postmenopausal osteoporosis by an experimental therapeutic antibody. J. Pharmacol. Exp. Ther. 2011, 338, 568–569. [Google Scholar] [CrossRef]
- Yaccoby, S.; Ling, W.; Zhan, F.; Walker, R.; Barlogie, B.; Shaughnessy, J.D. Antibody-based inhibition of DKK1 suppresses tumor-induced bone resorption and multiple myeloma growth in vivo. Blood 2007, 109, 2106–2111. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, Y.; Wei, X.; Zhang, Q. Correlations of DKK1 with pathogenesis and prognosis of human multiple myeloma. Cancer Biomark. 2019, 24, 195–201. [Google Scholar] [CrossRef]
- Bendell, J.C.; Murphy, J.E.; Mahalingam, D.; Halmos, B.; Sirard, C.A.; Landau, S.B.; Ryan, D.P. Phase I study of DKN-01, an anti-DKK1 antibody, in combination with paclitaxel (pac) in patients (pts) with DKK1+ relapsed or refractory esophageal cancer (EC) or gastro-esophageal junction tumors (GEJ). J. Clin. Oncol. 2016, 34, 111. [Google Scholar] [CrossRef]
- Betella, I.; Turbitt, W.J.; Szul, T.; Wu, B.; Martinez, A.; Katre, A.; Wall, J.A.; Norian, L.; Birrer, M.J.; Arend, R. Wnt signaling modulator DKK1 as an immunotherapeutic target in ovarian cancer. Gynecol. Oncol. 2020, 157, 765–774. [Google Scholar] [CrossRef]
- Shaw, A.R.; Suzuki, M. Immunology of Adenoviral Vectors in Cancer Therapy. Mol. Ther. Methods Clin. Dev. 2019, 15, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Sawahara, H.; Shiraha, H.; Uchida, D.; Kato, H.; Nagahara, T.; Iwamuro, M.; Kataoka, J.; Horiguchi, S.; Watanabe, M.; Sakaguchi, M.; et al. Novel REIC/Dkk-3-encoding adenoviral vector as a promising therapeutic agent for pancreatic cancer. Cancer Gene Ther. 2016, 23, 278–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawahara, H.; Shiraha, H.; Uchida, D.; Kato, H.; Kato, R.; Oyama, A.; Nagahara, T.; Iwamuro, M.; Horiguchi, S.; Tsutsumi, K.; et al. Promising therapeutic efficacy of a novel reduced expression in immortalized cells/dickkopf-3 expressing adenoviral vector for hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2017, 32, 1769–1777. [Google Scholar] [CrossRef]
- Veeck, J.; Dahl, E. Targeting the Wnt pathway in cancer: The emerging role of Dickkopf-3. Biochim. Biophys. Acta Rev. Cancer 2012, 1825, 18–28. [Google Scholar] [CrossRef]
- Qian, J.; Xie, J.; Hong, S.; Yang, J.; Zhang, L.; Han, X.; Wang, M.; Zhan, F.; Shaughnessy, J.D.; Epstein, J.; et al. Dickkopf-1 (DKK1) is a widely expressed and potent tumor-associated antigen in multiple myeloma. Blood 2007, 110, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Meng, S.; Jin, Y.; Zhang, W.; Li, Z.; Wang, F.; Wang-Johanning, F.; Wei, Y.; Liu, H.; Tu, H.; et al. A novel multi-epitope vaccine from MMSA-1 and DKK1 for multiple myeloma immunotherapy. Br. J. Haematol. 2017, 178, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Zheng, Y.; Zheng, C.; Wang, L.; Qin, H.; Hong, S.; Li, H.; Lu, Y.; He, J.; Yang, J.; et al. Active vaccination with Dickkopf-1 induces protective and therapeutic antitumor immunity in murine multiple myeloma. Blood 2012, 119, 161–169. [Google Scholar] [CrossRef]
- Liu, T.T.; Wu, Y.; Niu, T. Human DKK1 and human HSP70 fusion DNA vaccine induces an effective anti-tumor efficacy in murine multiple myeloma. Oncotarget 2018, 9, 178–191. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Zheng, C.; Wang, Q.; Bi, E.; Yang, M.; Hou, J.; Fu, W.; Yi, Q.; Qian, J. Identification of an immunogenic DKK1 long peptide for immunotherapy of human multiple myeloma. Haematologica 2021, 106, 838–846. [Google Scholar] [CrossRef] [Green Version]
- Evans, T.R.J.; Colston, K.W.; Lofts, F.J.; Cunningham, D.; Anthoney, D.A.; Gogas, H.; De Bono, J.S.; Hamberg, K.J.; Skov, T.; Mansi, J.L. A phase II trial of the vitamin D analogue seocalcitol (EB1089) in patients with inoperable pancreatic cancer. Br. J. Cancer 2002, 86, 680–685. [Google Scholar] [CrossRef]
- Iyer, S.P.; Beck, J.T.; Stewart, A.K.; Shah, J.; Kelly, K.R.; Isaacs, R.; Bilic, S.; Sen, S.; Munshi, N.C. A Phase IB multicentre dose-determination study of BHQ880 in combination with anti-myeloma therapy and zoledronic acid in patients with relapsed or refractory multiple myeloma and prior skeletal-related events. Br. J. Haematol. 2014, 167, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Kurozumi, K.; Fujii, K.; Shimazu, Y.; Tomita, Y.; Sasaki, T.; Yasuhara, T.; Hishikawa, T.; Kameda, M.; Kumon, H.; Date, I. Study protocol of a Phase I/IIa clinical trial of Ad-SGE-REIC for treatment of recurrent malignant glioma. Future Oncol. 2020, 16, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Oyama, A.; Shiraha, H.; Uchida, D.; Iwamuro, M.; Kato, H.; Takaki, A.; Ikeda, F.; Onishi, H.; Yasunaka, T.; Takeuchi, Y.; et al. A Phase I/Ib trial of Ad-REIC in liver cancer: Study protocol. Future Oncol. 2019, 15, 3547–3554. [Google Scholar] [CrossRef]
- Kumon, H.; Ariyoshi, Y.; Sasaki, K.; Sadahira, T.; Araki, M.; Ebara, S.; Yanai, H.; Watanabe, M.; Nasu, Y. Adenovirus vector carrying REIC/DKK-3 gene: Neoadjuvant intraprostatic injection for high-risk localized prostate cancer undergoing radical prostatectomy. Cancer Gene Ther. 2016, 23, 400–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, T.; Kurozumi, K.; Shimazu, Y.; Ichikawa, T.; Ishida, J.; Otani, Y.; Shimizu, T.; Tomita, Y.; Sakaguchi, M.; Watanabe, M.; et al. A super gene expression system enhances the anti-glioma effects of adenovirus-mediated REIC/Dkk-3 gene therapy. Sci. Rep. 2016, 6, 33319. [Google Scholar] [CrossRef] [Green Version]
- Nowak-Sliwinska, P.; Scapozza, L.; i Altaba, A.R. Drug repurposing in oncology: Compounds, pathways, phenotypes and computational approaches for colorectal cancer. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 434–454. [Google Scholar] [CrossRef]
- Marcinowska-Suchowierska, E.; Kupisz-Urbańska, M.; Łukaszkiewicz, J.; Płudowski, P.; Jones, G. Vitamin D Toxicity-A Clinical Perspective. Front. Endocrinol. 2018, 9, 550. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.M.; Shin, E.A. Exploring vitamin D metabolism and function in cancer. Exp. Mol. Med. 2018, 50, 20. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, A.V.; Trump, D.L.; Johnson, C.S.; Feldman, D. The role of vitamin D in cancer prevention and treatment. Endocrinol. Metab. Clin. N. Am. 2010, 39, 401–418. [Google Scholar] [CrossRef] [Green Version]
- Garland, C.F.; Garland, F.C.; Gorham, E.D.; Lipkin, M.; Newmark, H.; Mohr, S.B.; Holick, M.F. The role of vitamin D in cancer prevention. Am. J. Public Health 2006, 96, 252–261. [Google Scholar] [CrossRef]
- Young, M.R.I.; Xiong, Y. Influence of vitamin D on cancer risk and treatment: Why the variability? Trends Cancer Res. 2018, 13, 43–53. [Google Scholar] [PubMed]
- Pendás-Franco, N.; Aguilera, Ó.; Pereira, F.; González-Sancho, J.M.; Muñoz, A. Vitamin D and Wnt/β-catenin Pathway in Colon Cancer: Role and regulation of DICKKOPF genes. Anticancer Res. 2008, 28, 2613–2623. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Name | Human Gene(s) | Protein(s) Inhibited | Main Cellular Location |
|---|---|---|---|
| WIF1 | WIF1 | Wnt | Extracellular |
| Cerberus | CER1 | Wnt | Extracellular |
| Tiki family | TIKI (1–2) | Wnt | Transmembrane |
| sFRP family | sFRP (1–5) | Wnt and Fz | Extracellular |
| APCDD1 | APCDD1 | Wnt and LRP | Transmembrane |
| DKK family | DKK (1–4) | LRP | Extracellular |
| SOST/Sclerostin | SOST | LRP | Extracellular |
| Wise | SOSTDC1 | LRP | Extracellular |
| MESD | MESD | LRP | Endoplasmic reticulum |
| Waif | TPBG | LRP | Transmembrane |
| IGFBP-4 | IGFBP4 | LRP and Fz | Extracellular |
| Shisa family | SHISA (2–9) | Fz | Transmembrane |
| Function | Chemical Structure | Compound Name | Target | Mechanism of Action | Disease (Pre-Clinical) |
|---|---|---|---|---|---|
| inhibitor | small molecules | WAY-262611 | DKK-1 | Inhibits DKK-1 impeding DKK1-LRP5/6 interaction [89] | Rheumatoid arthritis Leishmaniosis AML MM |
| NCI8642 | DKK-1 and DKK-2 | Binds to LRP6 disrupting DKK1-LRP6 interaction [90] | Alzheimer | ||
| L-securinine | DKK-1 | Promotes DKK-1 promoter methylation inhibiting DKK-1 expression [91] | Lung cancer | ||
| monoclonal antibodies | RH2-18 | DKK-1 | Binds to DKK-1 Cys-2 domain [92] | Osteoporosis | |
| BHQ880 | DKK-1 | Binds to DKK-1 Cys-2 domain [93] | Osteosarcoma MM | ||
| DKN-01 | DKK-1 | Binds to DKK-1 Cys-2 domain [94] | Solid tumors | ||
| 5F8 | DKK-2 | Blocks the binding of DKK2 to LRP5 [39] | colorectal cancer | ||
| activator | vitamin D3 | Calcitriol | DKK-1 | Induce the transcription of DKK-1 gene [95] | Osteoporosis Solid tumors |
| vitamin D3 analogue | Seocalcitol | DKK-1 | Induce the transcription of DKK-1 gene [95] | Hepatocellular carcinoma |
| Compound/Strategy | Target | Phase | Tumor | ID |
|---|---|---|---|---|
| DKN-01 | DKK-1 | I | Esophagogastric Malignancies | NCT02013154 |
| MM, solid tumors, NSCLC | NCT01457417 | |||
| Relapsed MM | NCT01711671 | |||
| Intra- or extra-Hepatic biliary system | NCT02375880 | |||
| Ib/IIa | Prostate | NCT03837353 | ||
| I/II | Hepatocellular Carcinoma | NCT03645980 | ||
| II | Endometrial, uterine and ovarian | NCT03395080 | ||
| Endometrial and Ovarian | NCT03395080 | |||
| BTC and EGC | NCT03818997 | |||
| BHQ880 | DKK-1 | I/II | Relapsed MM | NCT00741377 |
| II | MM | NCT01337752 | ||
| Smouldering MM | NCT01302886 | |||
| Calcitriol (DN-101) | DKK-1 | I | Solid tumors | NCT01588522 |
| CNS | NCT00008086 | |||
| Prostate | NCT00004928 | |||
| Prostate | NCT00010231 | |||
| I/II | Carcinoma, NSCLS | NCT00066885 | ||
| NSCLC | NCT00794547 | |||
| Metastatic Melanoma | NCT00301067 | |||
| II | CCA | NCT01039181 | ||
| CRPC | NCT03261336 | |||
| AIPC and NSCLC | NCT00285675 | |||
| Prostate | NCT00004043 | |||
| Prostate | NCT00524589 | |||
| Prostate (stage IV) | NCT00017576 | |||
| Prostate (metastatic) | NCT00182741 | |||
| Pancreas | NCT00238199 | |||
| Pancreas | NCT00536770 | |||
| Adenocarcinoma | NCT00084864 | |||
| BCC | NCT01358045 | |||
| II/III | Prostate cancer | NCT00043576 | ||
| Seocalcitol | DKK-1 | III | Hepatocellular Carcinoma | NCT00051532 |
| Dendritic cell vaccine | DKK-1 | Early I | Stable and Smouldering MM | NCT03591614 |
| Ad-REIC | DKK-3 | I | Prostate | NCT01197209 |
| I/IIa | Prostate | NCT01931046 | ||
| I/IIa | Glioma | [122] | ||
| I/Ib | Liver | [123] | ||
| II | Mesothelioma | NCT04013334 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giralt, I.; Gallo-Oller, G.; Navarro, N.; Zarzosa, P.; Pons, G.; Magdaleno, A.; Segura, M.F.; Sánchez de Toledo, J.; Moreno, L.; Gallego, S.; et al. Dickkopf Proteins and Their Role in Cancer: A Family of Wnt Antagonists with a Dual Role. Pharmaceuticals 2021, 14, 810. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080810
Giralt I, Gallo-Oller G, Navarro N, Zarzosa P, Pons G, Magdaleno A, Segura MF, Sánchez de Toledo J, Moreno L, Gallego S, et al. Dickkopf Proteins and Their Role in Cancer: A Family of Wnt Antagonists with a Dual Role. Pharmaceuticals. 2021; 14(8):810. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080810
Chicago/Turabian StyleGiralt, Irina, Gabriel Gallo-Oller, Natalia Navarro, Patricia Zarzosa, Guillem Pons, Ainara Magdaleno, Miguel F. Segura, José Sánchez de Toledo, Lucas Moreno, Soledad Gallego, and et al. 2021. "Dickkopf Proteins and Their Role in Cancer: A Family of Wnt Antagonists with a Dual Role" Pharmaceuticals 14, no. 8: 810. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080810