
In Vitro and In Vivo Activity of AS101 against Carbapenem-Resistant Acinetobacter baumannii

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Antibacterial Efficacy of AS101 against CRAB Clinical Isolates
2.2. Characterization of Antibacterial Activity of AS101
2.3. Antibacterial Mechanism of AS101
2.4. Effect on Bacterial Morphology
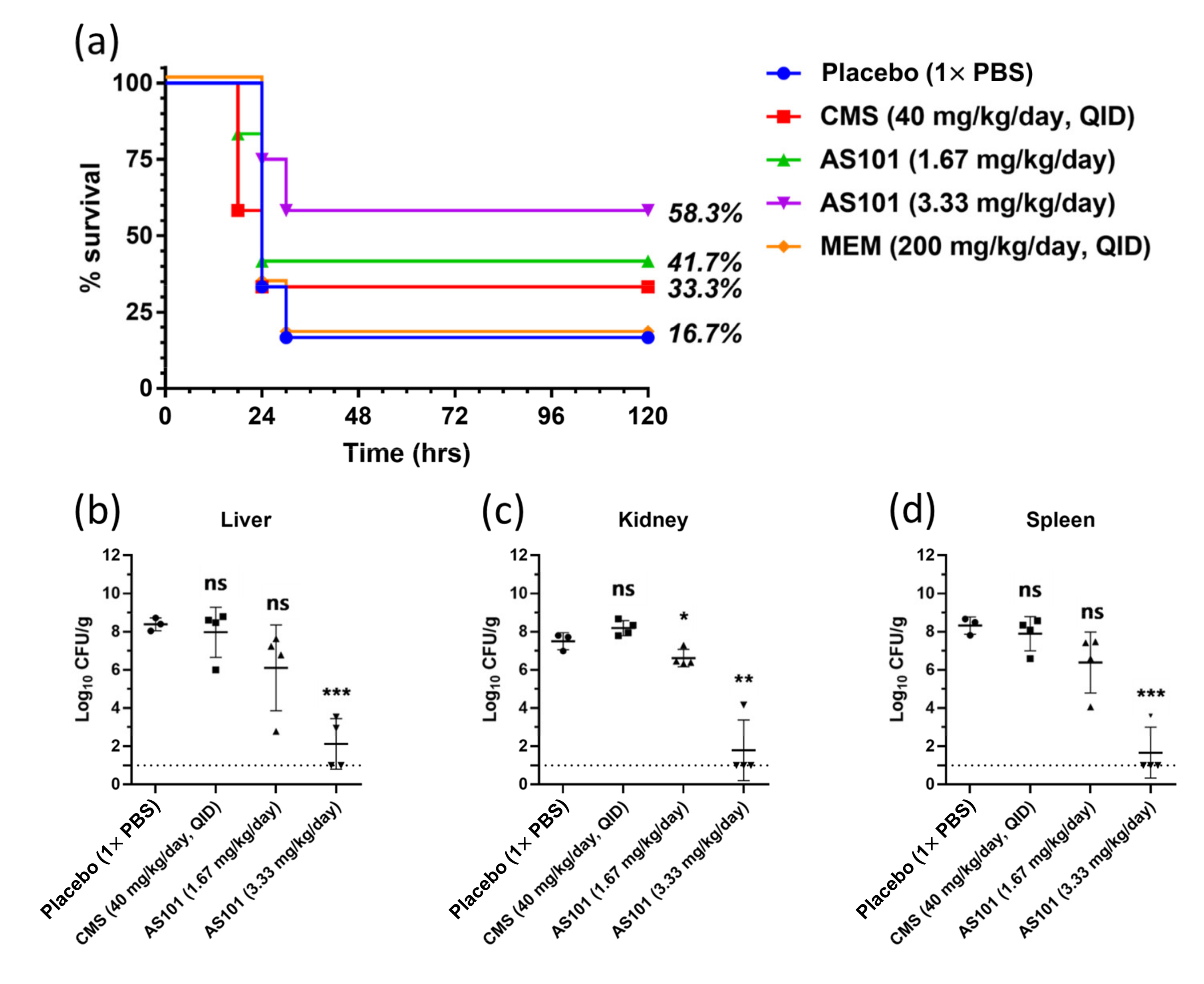
2.5. In Vivo Administration Test
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Susceptibility Testing
4.3. Antibacterial Activity of AS101
4.4. Detection of Carbapenemase
4.5. Time-Kill Kinetics Assay
4.6. Pharmacological Manipulation
4.7. Reactive Oxygen Species (ROS) Generation Test
4.8. Electron Microscopy
4.9. Animal Model
4.10. Bacterial Infection and Survival
4.11. Organ Bacterial Load
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ali, H.M.; Salem, M.Z.; El-Shikh, M.S.; Abdel-Megeed, A.; Alogaibi, Y.A.; Talea, I.A. Investigation of the Virulence Factors and Molecular Characterization of the Clonal Relations of Multidrug-Resistant Acinetobacter baumannii Isolates. J. AOAC Int. 2017, 100, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.; Alvi, I.A.; Rehman, S.U. Insight into Acinetobacter baumannii: Pathogenesis, global resistance, mechanisms of resistance, treatment options, and alternative modalities. Infect. Drug Resist. 2018, 11, 1249–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labarca, J.A.; Salles, M.J.; Seas, C.; Guzman-Blanco, M. Carbapenem resistance in Pseudomonas aeruginosa and Acinetobacter baumannii in the nosocomial setting in Latin America. Crit. Rev. Microbiol. 2016, 42, 276–292. [Google Scholar] [PubMed]
- Sievert, D.M.; Ricks, P.; Edwards, J.R.; Schneider, A.; Patel, J.; Srinivasan, A.; Kallen, A.; Limbago, B.; Fridkin, S.; National Healthcare Safety Network (NHSN) Team and Participating NHSN Facilities. Antimicrobial-Resistant Pathogens Associated with Healthcare-Associated Infections Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–2010. Infect. Control Hosp. Epidemiol. 2013, 34, 1–14. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Antimicrobial resistance in the EU/EEA (EARS-Net)—Annual Epidemiological Report 2019; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2020. [Google Scholar]
- Towse, A.; Hoyle, C.K.; Goodall, J.; Hirsch, M.; Mestre-Ferrandiz, J.; Rex, J. Time for a change in how new antibiotics are reimbursed: Development of an insurance framework for funding new antibiotics based on a policy of risk mitigation. Health Policy 2017, 121, 1025–1030. [Google Scholar] [CrossRef]
- Luepke, K.H.; Suda, K.J.; Boucher, H.; Russo, R.L.; Bonney, M.W.; Hunt, T.D.; Mohr, J.F., 3rd. Past, present, and future of antibacterial rconomics: Increasing bacterial resistance, limited antibiotic pipeline, and societal implications. Pharmacotherapy 2017, 37, 71–84. [Google Scholar] [CrossRef]
- Cruz-Muñiz, M.Y.; López-Jacome, L.E.; Hernández-Durán, M.; Franco-Cendejas, R.; Licona-Limón, P.; Ramos-Balderas, J.L.; Martínez-Vázquez, M.; Belmont-Díaz, J.A.; Wood, T.K.; García-Contreras, R. Repurposing the anticancer drug mitomycin C for the treatment of persistent Acinetobacter baumannii infections. Int. J. Antimicrob. Agents 2017, 49, 88–92. [Google Scholar] [CrossRef]
- Kwan, B.W.; Chowdhury, N.; Wood, T.K. Combatting bacterial infections by killing persister cells with mitomycin C. Environ. Microbiol. 2015, 17, 4406–4414. [Google Scholar] [CrossRef]
- Sredni, B.; Caspi, R.; Klein, A.; Kalechman, Y.; Danziger, Y.; BenYa’Akov, M.; Tamari, T.; Shalit, F.; Albeck, M. A new immunomodulating compound (AS-101) with potential therapeutic application. Nature 1987, 330, 173–176. [Google Scholar] [CrossRef]
- Strassmann, G.; Kambayashi, T.; Jacob, C.O.; Sredni, D. The immunomodulator AS-101 inhibits IL-10 release and augments TNF alpha and IL-1 alpha release by mouse and human mononuclear phagocytes. Cell Immunol. 1997, 176, 180–185. [Google Scholar] [CrossRef]
- Kalechman, Y.; Gafter, U.; Gal, R.; Rushkin, G.; Yan, D.; Albeck, M.; Sredni, B. Anti-IL-10 Therapeutic Strategy Using the Immunomodulator AS101 in Protecting Mice from Sepsis-Induced Death: Dependence on Timing of Immunomodulating Intervention. J. Immunol. 2002, 169, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Indenbaum, V.; Bin, H.; Makarovsky, D.; Weil, M.; Shulman, L.M.; Albeck, M.; Sredni, B.; Mendelson, E. In vitro and in vivo activity of AS101 against West Nile virus (WNV). Virus Res. 2012, 166, 68–76. [Google Scholar] [CrossRef]
- Sredni, B.; Albeck, M.; Kazimirsky, G.; Shalit, F. The immunomodulator AS101 administered orally as a chemoprotective and radioprotective agent. Int. J. Immunopharmacol. 1992, 14, 613–619. [Google Scholar] [CrossRef]
- Vonsover, A.; Loya, S.; Sredni, B.; Albeck, M.; Gotlieb-Stematsky, T.; Araf, O.; Hizi, A. Inhibition of the Reverse Transcriptase Activity and Replication of Human Immunodeficiency Virus Type 1 by AS 101 In Vitro. AIDS Res. Hum. Retrovir. 1992, 8, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Halpert, G.; Sredni, B. The effect of the novel tellurium compound AS101 on autoimmune diseases. Autoimmun. Rev. 2014, 13, 1230–1235. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt-Bin, H.; Kalechman, Y.; Vonsover, A.; Xu, R.H.; Da, J.P.; Shalit, F.; Huberman, M.; Klein, A.; Strassmann, G.; Albeck, M.; et al. The immunomodulator AS101 restores T(H1) type of response suppressed by Babesia rodhaini in BALB/c mice. Cell Immunol. 1998, 184, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Daniel-Hoffmann, M.; Albeck, M.; Sredni, B.; Nitzan, Y. A potential antimicrobial treatment against ESBL-producing Klebsiella pneumoniae using the tellurium compound AS101. Arch. Microbiol. 2009, 191, 631–638. [Google Scholar] [CrossRef]
- Daniel-Hoffmann, M.; Sredni, B.; Nitzan, Y. Bactericidal activity of the organo-tellurium compound AS101 against Enterobacter cloacae. J. Antimicrob. Chemother. 2012, 67, 2165–2172. [Google Scholar] [CrossRef]
- Yang, T.-Y.; Kao, H.-Y.; Lu, P.-L.; Chen, P.-Y.; Wang, S.-C.; Wang, L.-C.; Hsieh, Y.-J.; Tseng, S.-P. Evaluation of the Organotellurium Compound AS101 for Treating Colistin- and Carbapenem-Resistant Klebsiella pneumoniae. Pharmaceuticals 2021, 14, 795. [Google Scholar] [CrossRef]
- World Health Organisation. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery and Development of New Antibiotics; WHO Press: Geneva, Switzerland, 2017; p. 7. Available online: https://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf (accessed on 18 August 2021).
- Isler, B.; Doi, Y.; Bonomo, R.A.; Paterson, D.L. New Treatment Options against Carbapenem-Resistant Acinetobacter baumannii Infections. Antimicrob. Agents Chemother. 2019, 63, e01110-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, J.M.; Turton, J.F.; Kaufmann, M.E.; Glover, J.; Woodford, N.; Warner, M.; Palepou, M.-F.; Pike, R.; Pitt, T.L.; Patel, B.C.; et al. Occurrence of Carbapenem-Resistant Acinetobacter baumannii Clones at Multiple Hospitals in London and Southeast England. J. Clin. Microbiol. 2006, 44, 3623–3627. [Google Scholar] [CrossRef] [Green Version]
- Peleg, A.Y.; Franklin, C.; Bell, J.M.; Spelman, D.W. Emergence of Carbapenem Resistance in Acinetobacter baumannii Recovered from Blood Cultures in Australia. Infect. Control Hosp. Epidemiol. 2006, 27, 759–761. [Google Scholar] [CrossRef]
- Hu, Y.-F. Emergence of carbapenem-resistant Acinetobacter baumannii ST787 in clinical isolates from blood in a tertiary teaching hospital in Northern Taiwan. J. Microbiol. Immunol. Infect. 2017, 50, 640–645. [Google Scholar] [CrossRef]
- Carlson-Banning, K.M.; Chou, A.; Liu, Z.; Hamill, R.J.; Song, Y.; Zechiedrich, L. Toward Repurposing Ciclopirox as an Antibiotic against Drug-Resistant Acinetobacter baumannii, Escherichia coli, and Klebsiella pneumoniae. PLoS ONE 2013, 8, e69646. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, A.; Kakar, P.N.; Arora, D.; Das, S.; Govil, P. Colistin and polymyxin B: A re-emergence. Indian J. Crit. Care Med. 2009, 13, 49–53. [Google Scholar] [CrossRef]
- Shen, B.; Jensen, R.G.; Bohnert, H.J. Mannitol Protects against Oxidation by Hydroxyl Radicals. Plant Physiol. 1997, 115, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Zeng, J.; Wang, X.; Drlica, K.; Zhao, X. Post-stress bacterial cell death mediated by reactive oxygen species. Proc. Natl. Acad. Sci. USA 2019, 116, 10064–10071. [Google Scholar] [CrossRef] [Green Version]
- Kohanski, M.A.; Tharakan, A.; London, N.R.; Lane, A.P.; Ramanathan, M. Bactericidal antibiotics promote oxidative damage and programmed cell death in sinonasal epithelial cells. Int. Forum Allergy Rhinol. 2017, 7, 359–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwyer, D.J.; Kohanski, M.A.; Hayete, B.; Collins, J.J. Gyrase inhibitors induce an oxidative damage cellular death pathway in Escherichia coli. Mol. Syst. Biol. 2007, 3, 91. [Google Scholar] [CrossRef] [PubMed]
- Endres, B.; Bassères, E.; Rashid, T.; Chang, L.; Alam, M.J.; Garey, K.W. A Protocol to Characterize the Morphological Changes of Clostridium difficile in Response to Antibiotic Treatment. J. Vis. Exp. 2017, 55383. [Google Scholar] [CrossRef] [PubMed]
- Mehendiratta, M.; Bishen, K.A.; Boaz, K.; Mathias, Y. Ghost cells: A journey in the dark…. Dent. Res. J. 2012, 9, S1–S8. [Google Scholar]
- Tang, H.-J.; Chuang, Y.-C.; Ko, W.-C.; Chen, C.-C.; Shieh, J.-M.; Chen, C.-H.; Lee, N.-Y.; Chiang, S.-R. Comparative evaluation of intratracheal colistimethate sodium, imipenem, and meropenem in BALB/c mice with carbapenem-resistant Acinetobacter baumannii pneumonia. Int. J. Infect. Dis. 2012, 16, e34–e40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergen, P.J.; Li, J.; Rayner, C.R.; Nation, R.L. Colistin Methane sulfonate Is an Inactive Prodrug of Colistin against Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2006, 50, 1953–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Bou, G.; Oliver, A.; Martinez-Beltran, J. OXA-24, a novel class D beta-lactamase with carbapenemase activity in an Acinetobacter baumannii clinical strain. Antimicrob. Agents Chemother. 2000, 44, 1556–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, J.; Lane, C.R.; Romanes, F.; Da Silva, A.G.; Easton, M.; Cronin, K.; Waters, M.J.; Tomita, T.; Stevens, K.; Schultz, M.; et al. Translating genomics into practice for real-time surveillance and response to carbapenemase-producing Enterobacteriaceae: Evidence from a complex multi-institutional KPC outbreak. PeerJ 2018, 6, e4210. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.Y.; Hung, W.W.; Lin, L.; Hung, W.C.; Tseng, S.P. mecA-related structure in methicillin-resistant coagulase-negative staphylococci from street food in Taiwan. Sci. Rep. 2017, 7, 42205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.C.; Huang, Y.T.; Chen, C.S.; Chen, Y.W.; Huang, Y.T.; Su, J.C.; Teng, L.J.; Shiau, C.W.; Chiu, H.C. In vitro and in vivo activity of a novel sorafenib derivative SC5005 against MRSA. J. Antimicrob. Chemother. 2016, 71, 449–459. [Google Scholar] [CrossRef]
- Alhanout, K.; Malesinki, S.; Vidal, N.; Peyrot, V.; Rolain, J.-M.; Brunel, J.M. New insights into the antibacterial mechanism of action of squalamine. J. Antimicrob. Chemother. 2010, 65, 1688–1693. [Google Scholar] [CrossRef]
- Yang, T.-Y.; Hsieh, Y.-J.; Lu, P.-L.; Lin, L.; Wang, L.-C.; Wang, H.-Y.; Tsai, T.-H.; Shih, C.-J.; Tseng, S.-P. In vitro and in vivo assessments of inspired Ag/80S bioactive nanocomposites against carbapenem-resistant Klebsiella pneumoniae. Mater. Sci. Eng. C 2021, 125, 112093. [Google Scholar] [CrossRef]
- Zahller, J.; Stewart, P. Transmission Electron Microscopic Study of Antibiotic Action on Klebsiella pneumoniae Biofilm. Antimicrob. Agents Chemother. 2002, 46, 2679–2683. [Google Scholar] [CrossRef] [Green Version]
- King, A.M.; Reid-Yu, S.A.; Wang, W.; King, D.T.; de Pascale, G.; Strynadka, N.C.; Walsh, T.R.; Coombes, B.K.; Wright, G.D. Aspergillomarasmine A overcomes metallo-beta-lactamase antibiotic resistance. Nature 2014, 510, 503–506. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.-C.; Wang, X.; Qin, W.-T.; Qiu, X.-F.; Sun, B.-W. Exogenous carbon monoxide suppresses Escherichia coli vitality and improves survival in an Escherichia coli-induced murine sepsis model. Acta Pharmacol. Sin. 2014, 35, 1566–1576. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | MIC of AS101 | Susceptibility to Antimicrobial Agents 1 | Gene Detection of Carbapenemase | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CL | GM | DOX | CAZ | TIC | LEV | MEM | |||
| TSP-AB-1 | 8 | S | R | R | R | R | R | R | blaOXA-24, blaOXA-69 |
| TSP-AB-2 | 2 | S | R | S | R | R | R | R | blaIMP-1, blaOXA-69 |
| TSP-AB-3 | 2 | S | R | R | R | R | R | R | blaOXA-24, blaOXA-69 |
| TSP-AB-4 | 8 | S | R | R | R | R | R | R | blaOXA-24, blaOXA-69 |
| TSP-AB-5 | 2 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-6 | 8 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-7 | 2 | S | S | S | R | R | I | R | blaOXA-24, blaOXA-69 |
| TSP-AB-8 | 8 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-9 | 2 | S | R | R | R | R | R | R | blaOXA-24, blaOXA-69 |
| TSP-AB-10 | 1 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-11 | 1 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-12 | 16 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-13 | 32 | S | R | R | R | R | R | R | blaVIM-2, blaOXA-69 |
| TSP-AB-14 | 0.5 | S | R | R | R | R | R | R | - |
| TSP-AB-15 | 4 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-16 | 4 | S | S | R | R | R | R | R | blaOXA-24, blaOXA-69 |
| TSP-AB-17 | 2 | S | R | R | R | R | R | R | - |
| TSP-AB-18 | 0.5 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-19 | 1 | S | R | R | R | R | R | R | - |
| TSP-AB-20 | 1 | S | R | R | R | R | R | R | - |
| TSP-AB-21 | 2 | S | I | R | R | R | R | R | blaOXA-24, blaOXA-69 |
| TSP-AB-22 | 2 | S | R | R | R | R | R | R | blaOXA-24, blaOXA-69 |
| TSP-AB-23 | 1 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-24 | 8 | S | I | R | R | R | R | R | blaOXA-69 |
| TSP-AB-25 | 1 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-26 | 2 | S | R | R | R | R | R | R | blaOXA-69 |
| TSP-AB-27 | 8 | S | S | R | R | R | R | R | blaOXA-69 |
| Resistant rate | N/A 2 | 0% | 81.5% | 92.6% | 100% | 100% | 96.3% | 100% | N/A |
| 0/27 | 22/27 | 25/27 | 27/27 | 27/27 | 26/27 | 27/27 | |||
| Treatment | Function | MIC of AS101 Mean ± SD (μg/mL) 1 |
|---|---|---|
| Untreated | - | 2 ± 0 |
| 1 mM EDTA | Alteration of outer-membrane permeability | 0.21 ± 0.07 |
| 10 mM Ca2+ | Alteration of outer-membrane charge | 4 ± 0 |
| 10 mM Mg2+ | Alteration of outer-membrane charge | 4 ± 0 |
| 500 mM Mannitol | Reactive oxygen species (ROS) scavenger | 24 ± 9.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.-Y.; Tseng, S.-P.; Dlamini, H.N.; Lu, P.-L.; Lin, L.; Wang, L.-C.; Hung, W.-C. In Vitro and In Vivo Activity of AS101 against Carbapenem-Resistant Acinetobacter baumannii. Pharmaceuticals 2021, 14, 823. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080823
Yang T-Y, Tseng S-P, Dlamini HN, Lu P-L, Lin L, Wang L-C, Hung W-C. In Vitro and In Vivo Activity of AS101 against Carbapenem-Resistant Acinetobacter baumannii. Pharmaceuticals. 2021; 14(8):823. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080823
Chicago/Turabian StyleYang, Tsung-Ying, Sung-Pin Tseng, Heather Nokulunga Dlamini, Po-Liang Lu, Lin Lin, Liang-Chun Wang, and Wei-Chun Hung. 2021. "In Vitro and In Vivo Activity of AS101 against Carbapenem-Resistant Acinetobacter baumannii" Pharmaceuticals 14, no. 8: 823. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080823