
Nitrative Stress and Auditory Dysfunction



1
Institute of Environmental Health Sciences, Wayne State University, Detroit, MI 48202, USA
2
Department of Pharmacology, Wayne State University, Detroit, MI 48202, USA
*
Author to whom correspondence should be addressed.
Pharmaceuticals 2022, 15(6), 649; https://0-doi-org.brum.beds.ac.uk/10.3390/ph15060649
Submission received: 21 April 2022
/
Revised: 12 May 2022
/
Accepted: 20 May 2022
/
Published: 24 May 2022
(This article belongs to the Special Issue Drug-induced Ototoxicity: Mechanisms and Otoprotective Strategies)
Abstract
:Nitrative stress is increasingly recognized as a critical mediator of apoptotic cell death in many pathological conditions. The accumulation of nitric oxide along with superoxide radicals leads to the generation of peroxynitrite that can eventually result in the nitration of susceptible proteins. Nitrotyrosine is widely used as a biomarker of nitrative stress and indicates oxidative damage to proteins. Ototoxic insults, such as exposure to noise and ototoxic drugs, enhance the generation of 3-nitrotyrosine in different cell types in the cochlea. Nitrated proteins can disrupt critical signaling pathways and eventually lead to apoptosis and loss of sensory receptor cells in the cochlea. Accumulating evidence shows that selective targeting of nitrative stress attenuates cellular damage. Anti-nitrative compounds, such as peroxynitrite decomposition catalysts and inducible nitric oxide synthase inhibitors, prevent nitrative stress-mediated auditory damage. However, the role of nitrative stress in acquired hearing loss and its potential significance as a promising interventional target is yet to be fully characterized. This review provides an overview of nitrative stress mechanisms, the induction of nitrative stress in the auditory tissue after ototoxic insults, and the therapeutic value of targeting nitrative stress for mitigating auditory dysfunction.
1. Introduction
Nitrative stress is a cellular stress phenomenon that is caused by the increased production and accumulation of reactive nitrogen species (RNS) intermediates and the inability of cells and tissues to remove these reactive products. Nitric oxide (NO), a key regulator of redox signaling, plays a central role in a number of physiological processes [1]. NO was first reported as a biological messenger molecule in 1987 when Bob Furchgott proposed that endothelium-derived relaxing factor (EDRF) might be NO [2]. The generation of NO is regulated by nitric oxide synthase (NOS) enzymes. Endogenously, three types of NOS exist, namely, the l-arginine-dependent neuronal nitric oxide synthase (nNOS), inducible nitric oxide synthase (iNOS), and endothelial nitric oxide synthase (eNOS) [1,2]. NO signaling is critical for vasodilation, bronchodilation, neurotransmission, inhibition of phagocyte and platelet aggregation, and inhibition of microbial activity [3,4,5,6,7]. The reaction of NO with partially reduced oxygen species or reactive oxygen species (ROS) can generate RNS intermediates such as peroxynitrite (OONO−), nitrogen dioxide (NO2), and dinitrogen trioxide (N2O3) [4]. NO and NO-derived RNS such as NO2 and N2O3 are implicated in neuronal cell death and apoptosis [3,4,5,6], while OONO− causes lipid peroxidation and oxidation of protein-associated thiol groups [6,7]. Additionally, these RNS intermediates can induce posttranslational modifications, namely, 3-nitrotyrosination and nitrosylation, both of which have significant pathophysiological consequences [7]. The purpose of this review is to provide an overview of cochlear nitrative stress mechanisms with special emphasis on nitration and nitrosylation of cochlear proteins and their role in auditory dysfunction.
2. Nitrative Stress-Induced Post-Translational Modifications
Protein nitration: Protein tyrosine nitration is a selective process where only a few proteins become nitrated on only one or few of the tyrosine residues [7,8]. Nitrotyrosine (3-NT) is formed by the substitution of a nitro group in place of hydrogen on carbon 3 in the phenolic ring of a tyrosine residue [8,9]. In vivo nitration mostly depends on superoxide (O2−) and NO as they react to form ONOO−. The accumulation of ONOO− leads to the formation of peroxynitrous acid (ONOOH), which homolyzes to nitrite (NO2) and hydroxyl (OH) radicals or, in the presence of CO2, forms the nitroso-peroxocarboxylate adduct, which homolyzes to NO2 and CO3− [9,10]. These one-electron oxidants (CO3−, OH, NO2) and Compound I of peroxidases can oxidize the phenolic ring of tyrosine to yield the tyrosyl radical (Tyr). The addition of NO2 to Tyr in a radical–radical termination reaction results in tyrosine nitration (Figure 1).
In addition, alternate pathways exist for nitration, especially those involving certain transition metal-containing proteins such as prostaglandin, endoperoxide H synthase, and MnSOD [8]. The analysis of different protein mixtures suggests that neither the abundance nor the number of tyrosine residues per protein molecules determine which protein is nitrated. However, some common features exist for proteins that are nitrated on tyrosine residues. These include (i) the presence of more acidic residues in the vicinity of target tyrosine (glutamic or aspartic residues), (ii) a small number of cysteine or methionine residues in the neighborhood of tyrosine residues, and (iii) the presence of loop- or turn-inducing amino acids such as proline or glycine. In basal conditions, the level of protein-bound 3-NT is low; however, in several pathological conditions, elevated levels of 3-NT are observed and used as a biomarker for these conditions [5,6,7,11,12,13,14].
Protein nitrosylation: S-Nitrosylation occurs by covalent attachment of a NO group to cysteine residue of specific proteins [15]. S-Nitrosylation of proteins can occur via (i) an oxidative pathway with NO in a higher oxidation status, (ii) a radical-mediated pathway with ·NO and thiol (RS·) radicals, (iii) metal-catalyzed RSNO formation in the presence of transition metals, and (iv) trans-nitrosylation [15] (Figure 1). The affinity of cysteine residues to NO can vary in different proteins. Although there are no general rules for determining which cysteine residues are susceptible to nitrosylation, analysis of NO transfer in proteins such as hemoglobin suggests the involvement of an acid–base motif in protein S-nitrosylation and denitrosylation [16,17]. The acid–base motif comprises flanking acidic (D, E) and basic (R, H, K) residues in the vicinity of reactive thiol cysteine sites. Hence, these can suppress or favor, respectively, the formation of nucleophilic thiolate through electrostatic interactions. Furthermore, this motif has been shown to be predictive in a number of cases. Additionally, the low pKa of cysteine also plays a role in S-nitrosylation [9]. It has been shown that the interaction between Cys thiols and aromatic side-chains in its vicinity promotes the formation of a thiolate anion, which enhances the potential for NO modification. S-Nitrosylated cysteines are also found in hydrophobic pockets of proteins [9], which can sequester or stabilize radicals to form S-nitrosylating species. To further understand and predict nitrosylated sites with accuracy computational studies are being employed. Unlike protein tyrosine nitration, protein S-nitrosylation is a less stable and easily reversible post-translational modification [18].
2.1. Pathophysiological Consequences of Nitrative Stress
Post-translational modifications determine the function, interaction, and signaling of proteins that eventually control the functional state of a cell. Both nitration and nitrosylation of proteins can interfere with signaling processes [15,19,20,21,22,23,24,25]. Although the nitration of tyrosine occurs at relatively low levels compared to tyrosine phosphorylation, it is more stable and capable of causing vital changes in biological function as it can modulate phosphorylation cascades. Furthermore, the addition of 3-NT can alter protein function because the binding of a nitro group on tyrosine reduces its pKa value from 10 to 7.2, which impacts its pH [9,26]. The development of pathological manifestations associated with protein nitration depends on the functional role of the specific proteins, whose nitration results in either loss or gain of function. Nitration of proteins such as MnSOD, glutathione reductase, prostacyclin synthase, and tyrosine hydroxylase leads to decreased activity, while cytochrome c, fibrinogen, glutathione S-transferase 1, and α-synuclein exhibit increased activity when nitrated [25,26,27,28,29,30,31]. Some proteins such as α1-chymotrypsin and transferrin do not show any change in function upon nitration [32]. Moreover, nitrated proteins are often targets for proteolytic degradation by the 20S proteosome. Although removal of modified proteins with altered function is essential, synthesis of new proteins is also critical for reestablishing homeostasis. However, if the rate of synthesis of a specific protein does not correspond with its rate of degradation, especially in proteins with long half-lives whose gene regulation may be less sensitive for changes, nitration can result in pathological phenotypes depending on the protein’s functional activity. Hence, establishing the identity of nitrated proteins is of paramount importance to envisage its functional implications.
S-Nitrosylation, another redox-based regulator of protein function, participates in a wide range of biological processes related to normal cellular function, as well as various pathophysiological conditions. Addition of S-nitrosothiols can affect protein activity such as translocation and protein function by modulating crosstalk between different molecules or by directly impacting the structure of the protein [16,18]. Furthermore, major protein post-translational modifications (acetylation, ubiquitination, sumoylation, etc.) are also affected and regulated by S-nitrosylation crosstalk. Protein S-nitrosylation can signal either cell death or cell survival according to the functional characteristics of its target protein. S-Nitrosylated proteins have been reported to regulate signaling pathways associated with neurodegeneration, apoptosis, cellular trafficking, DNA repair, muscle contractility, circulation, and cardioprotection [18,33,34,35,36]. S-Nitrosylation is easily reversible, which allows it to serve as an on/off switch to precisely modify protein function in response to cellular signals. Denitrosylation occurs through enzyme-mediated reactions or nonenzymatically by changes in the redox environment of the protein. Depending on the levels of cellular oxidative stress, this reversible S-nitrosylation, which plays a crucial role in NO cell signaling, can progress to an irreversible sulfonic acid modification resulting in cellular toxicity [18]. Protein S-nitrosylation plays a central role in regulating stress-induced apoptosis as it can signal either a pro- or an antiapoptotic response, according to the characteristics of its substrate protein [4]. Such stimulatory and inhibitory signaling allows S-nitrosylation to regulate both mitochondrial and nuclear programs of apoptosis and fine-tune the cellular responses to apoptotic stimuli [5,33].
2.2. Nitrative Stress and Otopathology
Nitrative stress has been reported in different types of auditory dysfunction such as noise-induced, age-related, and drug-induced hearing loss [37,38,39,40,41,42,43,44,45]. Although both protein nitration and S-nitrosylation can lead to cellular apoptosis, there are very few studies that reported the association of S-nitrosylation with cell death in the auditory system. The induction of nitrative stress is determined by measuring the changes in related biomarkers in the inner ear. Several studies have reported the association of nitrative stress with auditory dysfunction; however, very few studies have conducted an in-depth investigation of the associated signaling mechanism. This section briefly describes the otopathological conditions in which nitrative stress plays a role in causing auditory dysfunction. A list of studies that detected nitrative stress in the inner ear and/or analyzed their association with auditory dysfunction is provided in Table 1.
The link between noise-induced hearing loss (NIHL) and nitrative stress is well documented [14,21,22,23,45,46]. Studies on rodents (BALB/c mice or CBA/J mice or Wistar rats) indicated that ototoxic insults with either noise or ototoxic reagents increased the generation of RNS, and pretreatment with anti-nitrative agents protected against cochlear injury [43,47,48,49]. Both ROS and RNS were implicated in the loss of outer hair cells (OHCs) after noise exposure [23,39,44,50]. NO accumulation in the mitochondria of OHC during NIHL was reported to interfere with mitochondrial function in the guinea pig’s cochlea [41]. Noise exposure was also associated with an increase in the expression of endothelial nitric oxide synthase (eNOS) and induction of inducible nitric oxide synthase (iNOS) in the cochlea [50]. Furthermore, noise exposure increased the levels of 3-NT, a biomarker of nitrative stress, in the lateral wall, modiolar region, OHC, inner hair cells (IHC), and pillar cells in the guinea pig cochlea [22,43,47,48]. Studies on the relationship between autophagy and oxidative stress in NIHL indicated that 3-NT was elevated in OHC [23]. The accumulation of ROS/RNS after noise exposure activated the intrinsic Caspase-mediated apoptotic pathway via JNK and p38 MAPK signaling, resulting in cochlear cell death [43].
Nitrative stress also plays an important role in drug-induced hearing loss, especially after treatment with anti-cancer drugs such as cisplatin [47,48,49] or aminoglycosides such as gentamicin [51]. Anticancer drug cisplatin damaged the inner ear through generation of ROS and RNS and via the formation of DNA adducts [47]. The cisplatin-induced increase in the levels of 3-NT and peroxynitrite generation in mice cochlea accompanied the onset of cochlear apoptosis and hearing loss [47,48]. In addition, the levels of iNOS activity and Nf-κB were found to be significantly elevated. A strong correlation was observed between cisplatin-induced hearing loss and nitrative modifications of cochlear proteins in a dose-dependent manner. The most abundantly nitrated protein in the cochlea after cisplatin treatment was LMO4, a transcription factor regulator [47]. Nitration of Tyr-65 and Tyr-77 was detected in LMO4 by mass spectrometry. LMO4 levels were also decreased with cisplatin treatment, suggesting that this could be due to the degradation of nitrated LMO4, one of the consequences of protein nitration. In vitro studies with UB/OC-1 cell lines (derived from embryonic mouse inner ear) also verified the role of nitrative stress in cisplatin-induced ototoxicity [48,52]. These studies indicated that a marked increase in cell death, as well as Caspase-3 expression, a marker of apoptosis, after cisplatin treatment correlated with the increase in 3-NT levels [47]. This association was also detected in nonmammalian models such as zebrafish, where cisplatin-induced loss of hair cells in the neuromasts was accompanied by elevated expressions of 3-NT, decreased LMO4, and increased apoptosis [53]. Similarly, treatment of mouse organotypic cultures with gentamicin increased the 3-NT levels in the OHCs, indicating the induction of nitrative stress in aminoglycoside-mediated ototoxicity [51].
Nitrative stress was also reported in other otopathological conditions. Immunoreactivity to 3-NT was elevated in the Deiters’ cells, pillar cells, spiral ganglion cells, stria vascularis, and spiral ligament of aged CBA/J mice [54]. Immunostaining of 3-NT was found in vascular endothelial cells of the blood–labrynthine barrier in Meniere’s disease patients [55]. Together, these studies suggested that nitrative stress is a key factor in many otopathological conditions.
2.3. Assessment of Nitrative Stress
The effectiveness of interventional strategies that attenuate stress-induced damage to cells often depends on the early detection of oxidative/nitrative stress. Several molecules have been identified as reliable biomarkers of nitrative stress. Detection of these biomarkers is not only valuable for diagnostic purposes but also useful for assessing prognosis and determining the therapeutic value of interventions. A number of techniques are employed to detect different markers of nitrative stress in in vivo models. However, it is challenging to employ many of them for assessing the damage in the auditory organ because of the difficulty in accessing the inner ear and the short half-life of RNS such as ONOO−. For example, electrochemical sensors are used in other models to detect ONOO− released from living cells in trace amounts but are yet to be employed in the auditory system. Nevertheless, selective NO electrodes and electrometers have been used to assess NO concentration in the perilymph [44]. Changes in NO levels in auditory dysfunction can also be assessed using fluorescent dyes such as 4,5-diaminofluorescein diacetate (DAF) and 4,5-diaminofluorescein diacetate-2 (DAF-2), which probe for NO production in cells [44]. In addition, a chemiluminescence assay using nitrate reductase can be used to detect NO in the inner ear. The NOS family of enzymes that enable the production of NO are assessed as an indirect measure of nitrative stress. The expression of iNOS in the auditory organ has been reported in many studies and is measured by employing immunohistochemistry and/or immunoblotting methods [50]. However, the detection of 3-NT in cochlea appears to be the most commonly employed approach for measuring nitrative stress in the auditory system. Techniques such as immunoblotting, immunocytochemistry, and mass spectrometry are employed to detect 3-NT in the inner ear and to identify nitrated cochlear proteins [47,48,49]. Together, these biochemical techniques have enabled us to gain important insights into the role of nitrative stress in auditory dysfunction.
2.4. Targeting Nitrative Stress for Otoprotection
Numerous studies have indicated that nitration of proteins can be reversed, and targeting nitrative stress is a promising interventional strategy for mitigating oxidative damage to the cochlea [48,52,56,57,58]. Protein nitration and its signaling have been targeted to prevent apoptosis in organs such as the eye, pancreas, and kidney [59,60]. Unlike ROS, which has a role in normal cell signaling, a beneficial role of ONOO−, which causes cochlear damage, has not yet been reported in the auditory system. Although nitration of proteins occurs in tissues under physiological conditions [61], the use of peroxynitrite decomposition catalysts (PNDCs) does not interfere with physiological functions and, to date, no evidence exists that selective targeting of peroxynitrite has any side-effects. Treatment of mice with PNDCs such as SRI110, a metalloporphyrin, attenuated the cisplatin-induced increase in the levels of 3-NT and Caspase-3, while auditory brainstem responses (ABR) of 6 week old CBA/J mice indicated that SRI110 cotreatment mitigated cisplatin-induced hearing loss [48]. Further investigation of the role of PNDCs in mitigating cisplatin-induced ototoxicity suggested that SRI110 cotreatment inhibited cisplatin-induced protein nitration, prevented cisplatin-induced inactivation of Stat3 protein, a critical protein required for activating the Jak1/Stat3 pathway, and enhanced antiapoptotic signaling in UBOC1 cell lines [57]. Compounds such as ebselen, which is both a glutathione peroxidase mimic and an excellent scavenger of peroxynitrite radicals, and antioxidants, such as Trolox, were used to prevent the nitration of tyrosine in cochlear proteins and protect the cochlea from cisplatin-induced damage and hearing loss [47,62,63,64]. Although a direct link between inhibition of nitrative stress with ebselen and attenuation of NIHL is yet to be reported, phase 2 clinical trials in healthy adults aged 18–31 years indicated that 400 mg of ebselen treatment twice daily prevented temporary and permanent noise-induced hearing loss [64]. Treatment of mice with antagonists of nitric oxide synthase such as L-NAME attenuated the noise-induced increase in the level of 3-NT [43,58]. Additionally, compounds such as sodium butyrate, adenosine amine congener (ADAC), polyphenols, Astragaloside IV, L-cysteine, and N-acetyl cysteine (NAC) were reported to prevent cochlear nitrative stress and hearing loss [56,65,66,67,68,69]. Recent findings from our laboratory showed that selective scavenging of ONOO− using pure MnTBAP prevents the cisplatin-induced decrease in the amplitude and increase in the latency of ABR wave I in CBA/J mice, which suggests that targeting nitrative stress also prevents synaptic dysfunction and hidden hearing loss (unpublished data). Thus, compounds that target nitrative stress are increasingly used to confer significant otoprotection after noise- or drug-induced cochlear damage and hearing loss. More importantly, unlike antioxidants that were used to target oxidative stress for mitigating ototoxicity induced by the anticancer drug cisplatin, selective inhibition of nitrative stress by PNDCs does not interfere with the anticancer activity of a drug. Therefore, targeting nitrative stress meets the critical criteria for an effective therapeutic (i.e., otoprotective, safe, and well-tolerated, without compromising the activity of the primary drug). A list of studies that employed different pharmacological compounds to inhibit nitrative stress in the inner ear is provided in Table 2.
3. Conclusions
Overall, a plethora of studies have indicated that nitrative stress is an important contributor to auditory dysfunction after exposure to noise or other ototoxic agents. Although the number of studies on the role of S-nitrosylation in auditory dysfunction is limited, recent technological advancements have enabled the detection of multiple biomarkers of nitrative stress in the auditory organ, identification of nitrated cochlear proteins, and delineation of associated signaling mechanisms. Compelling evidence from recent studies indicated the otoprotective efficacy of anti-nitrative compounds. However, this promising target is yet to be fully exploited for treating sensory neural hearing loss. Moreover, the potential role of nitrative stress in sensorineural hearing loss induced by viruses and autoimmune disorders is yet to be fully understood [70,71]. Continued progress in understanding the role of protein nitration and nitrosylation mechanisms in different otopathological conditions would enable the design of effective interventional strategies and development of a potential class of novel drugs for acquired hearing loss.
Author Contributions
Conceptualization, S.J. and M.S.; writing—original draft preparation, M.S.; writing—review and editing, S.J. All authors read and agreed to the published version of the manuscript.
Funding
This research was supported by a CURES P30 center grant (P30 ES020957) from the NIH/NIEHS and a K01 grant (K01 ES028750) to S.J. from NIH/NIEHS.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Shahani, N.; Sawa, A. Protein S-nitrosylation: Role for nitric oxide signaling in neuronal death. Biochim. Biophys. Acta 2012, 1820, 736–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.T.; Pae, H.O.; Choi, B.M.; Billiar, T.R.; Kim, Y.M. Nitric oxide as a bioregulator of apoptosis. Biochem. Biophys. Res. Commun. 2001, 282, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.M.; Pae, H.O.; Jang, S.I.; Kim, Y.M.; Chung, H.T. Nitric oxide as a pro-apoptotic as well as anti-apoptotic modulator. J. Biochem. Mol. Biol. 2002, 35, 116–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.M.; Kim, P.K.; Kwon, Y.G.; Bai, S.K.; Nam, W.D.; Kim, Y.M. Regulation of apoptosis by nitrosative stress. J. Biochem. Mol. Biol. 2002, 35, 127–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razavi, H.M.; Hamilton, J.A.; Feng, Q. Modulation of apoptosis by nitric oxide: Implications in myocardial ischemia and heart failure. Pharmacol. Ther. 2005, 106, 147–162. [Google Scholar] [CrossRef]
- Martinez, M.C.; Andriantsitohaina, R. Reactive nitrogen species: Molecular mechanisms and potential significance in health and disease. Antioxid. Redox Signal. 2009, 11, 669–702. [Google Scholar] [CrossRef] [PubMed]
- Quijano, C.; Hernandez-Saavedra, D.; Castro, L.; McCord, J.M.; Freeman, B.A.; Radi, R. Reaction of peroxynitrite with Mn-superoxide dismutase. Role of the metal center in decomposition kinetics and nitration. J. Biol. Chem. 2001, 276, 11631–11638. [Google Scholar] [CrossRef] [Green Version]
- Radi, R. Nitric oxide, oxidants, and protein tyrosine nitration. Proc. Natl. Acad. Sci. USA 2004, 101, 4003–4008. [Google Scholar] [CrossRef] [Green Version]
- Donnini, S.; Monti, M.; Roncone, R.; Morbidelli, L.; Rocchigiani, M.; Oliviero, S.; Casella, L.; Giachetti, A.; Schulz, R.; Ziche, M. Peroxynitrite inactivates human-tissue inhibitor of metalloproteinase-4. FEBS Lett. 2008, 582, 1135–1140. [Google Scholar] [CrossRef] [Green Version]
- Yakovlev, V.A.; Bayden, A.S.; Graves, P.R.; Kellogg, G.E.; Mikkelsen, R.B. Nitration of the tumor suppressor protein p53 at tyrosine 327 promotes p53 oligomerization and activation. Biochemistry 2010, 49, 5331–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubbo, H.; Radi, R. Protein and lipid nitration: Role in redox signaling and injury. Biochim. Biophys. Acta 2008, 1780, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Lanone, S.; Manivet, P.; Callebert, J.; Launay, J.M.; Payen, D.; Aubier, M.; Boczkowski, J.; Mebazaa, A. Inducible nitric oxide synthase (NOS2) expressed in septic patients is nitrated on selected tyrosine residues: Implications for enzymic activity. Biochem. J. 2002, 366, 399–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohinata, Y.; Miller, J.M.; Schacht, J. Protection from noise-induced lipid peroxidation and hair cell loss in the cochlea. Brain Res. 2003, 966, 265–273. [Google Scholar] [CrossRef]
- Foster, M.W. S-nitrosylation in health and disease. Trends Mol. Med. 2003, 9, 160–168. [Google Scholar] [CrossRef]
- Kovacs, L.; Lindermayr, T. Nitric oxide-based protein modification: Formation and site-specificity of protein S-nitrosylation. Front. Plant Sci. 2013, 4, 137. [Google Scholar] [CrossRef] [Green Version]
- Hess, D.T.; Matsumoto, A.; Kim, S.; Marshall, H.E.; Stamler, J.S. Protein S-nitrosylation: Purview and parameters. Nat. Rev. Mol. Cell Biol. 2005, 6, 150–166. [Google Scholar] [CrossRef]
- Nakamura, T.; Lipton, S.A. Emerging roles of S-nitrosylation in protein misfolding and neurodegenerative diseases. Antioxid. Redox Signal. 2008, 10, 87–101. [Google Scholar] [CrossRef]
- Fukuyama, N.; Takizawa, S.; Ishida, H.; Hoshiai, K.; Shinohara, Y.; Nakazawa, H. Peroxynitrite formation in focal cerebral ischemia-reperfusion in rats occurs predominantly in the peri-infarct region. J. Cereb. Blood Flow Metab. 1998, 18, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Ozdemir, Y.; Bolay, H.; Saribas, O.; Dalkara, T. Role of Endothelial Nitric Oxide Generation and Peroxynitrite Formation in Reperfusion Injury After Focal Cerebral Ischemia. Stroke 2000, 31, 1974–1980. [Google Scholar] [CrossRef]
- Park, J.S.; Jou, I.; Park, S.M. Attenuation of noise-induced hearing loss using methylene blue. Cell Death Dis. 2014, 5, e1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, W.; Shi, X.; Nuttall, A. Noise-induced nitrotyrosine increase and outer hair cell death in guinea pig cochlea. Chin. Med. J. 2013, 126, 2923–2927. [Google Scholar] [PubMed]
- Yuan, H.; Wang, X.; Hill, K.; Chen, J.; Lemasters, J.; Yang, S.; Sha, S. Autophagy Attenuates Noise-Induced Hearing Loss by Reducing Oxidative Stress. Antioxid. Redox Signal. 2015, 22, 1308–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, J.M.; Daikhin, E.; Yudkoff, M.; Raman, C.S.; Ischiropoulos, H. Factors determining the selectivity of protein tyrosine nitration. Arch. Biochem. Biophys. 1999, 371, 169–178. [Google Scholar] [CrossRef] [PubMed]
- MacMillan-Crow, L.A.; Crow, J.P.; Kerby, J.D.; Beckman, J.S.; Thompson, J.A. Nitration and inactivation of manganese superoxide dismutase in chronic rejection of human renal allografts. Proc. Natl. Acad. Sci. USA 1996, 93, 11853–11858. [Google Scholar] [CrossRef] [Green Version]
- Cassina, A.M.; Hodara, R.; Souza, J.M.; Thomson, L.; Castro, H.; Ischiropoulos, H.; Freeman, B.; Radi, R. Cytochrome C nitration by peroxynitrite. J. Biol. Chem. 2000, 275, 21409–21415. [Google Scholar] [CrossRef] [Green Version]
- Savvides, S.N.; Scheiwein, M.; Bohme, C.C.; Artee, G.E.; Karplus, A.; Becker, K.; Heiner, R.S. Crystal structure of the antioxidant enzyme glutathione reductase inactivated by peroxynitrite. J. Biol. Chem. 2002, 277, 2779–2784. [Google Scholar] [CrossRef] [Green Version]
- Zou, M.; Martin, C.; Ullrich, V. Tyrosine nitration as a mechanism of selective inactivation of prostacyclin synthase by peroxynitrite. Biol. Chem. 1997, 378, 707–713. [Google Scholar] [CrossRef]
- Blanchard-Fillion, B.; Servy, C.; Ducrocq, C. 1-Nitrosomelatonin is a spontaneous NO-releasing compound. Free Radic. Res. 2001, 35, 857–866. [Google Scholar] [CrossRef]
- Vadseth, C.; Souza, J.M.; Thomson, L.; Seagraves, A.; Nagaswami, C.; Scheiner, T.; Torbet, J.; Vilaire, G.; Bennett, J.S.; Murciano, J.; et al. Pro-thrombotic state induced by post-translational modification of fibrinogen by reactive nitrogen species. J. Biol. Chem. 2004, 279, 8820–8826. [Google Scholar] [CrossRef] [Green Version]
- Hodara, R.; Norris, E.H.; Giasson, B.I.; Mishizen-Eberz, A.J.; Lynch, D.R.; Lee, V.; Ischiropoulos, H. Functional consequences of alpha-synuclein tyrosine nitration: Diminished binding to lipid vesicles and increased fibril formation. J. Biol. Chem. 2004, 279, 47746–47753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gole, M.D.; Souza, J.M.; Choi, I.; Hertkorn, C.; Malcolm, S.; FoustIII, R.F.; Finkel, B.; Lanken, P.N.; Ischiropoulos, H. Plasma proteins modified by tyrosine nitration in acute respiratory distress syndrome. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 278, L961–L967. [Google Scholar] [CrossRef] [PubMed]
- Forrester, M.T.; Benhar, M.; Stamler, J.S. Nitrosative stress in the ER: A new role for S-nitrosylation in neurodegenerative diseases. ACS Chem. Biol. 2006, 1, 355–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benhar, M.; Stamler, J.S. A central role for S-nitrosylation in apoptosis. Nat. Cell Biol. 2005, 7, 645–646. [Google Scholar] [CrossRef]
- Ozawa, K.; Komatsubara, A.T.; Nishimura, Y.; Sawada, T.; Kawafune, H.; Tsumoto, H.; Tsuji, Y.; Zhao, J.; Kyotani, Y.; Tanaka, T.; et al. S-nitrosylation regulates mitochondrial quality control via activation of parkin. Sci. Rep. 2013, 3, 2202. [Google Scholar] [CrossRef] [Green Version]
- Singel, D.S.; Stamler, J.S. Chemical physiology of blood flow regulation by red blood cells: The role of nitric oxide and S-nitrosohemoglobin. Annu. Rev. Physiol. 2005, 67, 99–145. [Google Scholar] [CrossRef]
- Jamesdaniel, S.; Manohar, S.; Hinduja, S. Is S-nitrosylation of cochlear proteins a critical factor in cisplatin-induced ototoxicity? Antioxid. Redox Signal. 2012, 17, 929–933. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, D.; Jiang, H.; Schacht, J.; Miller, J.M. Delayed production of free radicals following noise exposure. Brain Res. 2004, 1019, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, D.; Jiang, H.Y.; LePrell, C.G.; Schacht, J.; Miller, J.M. Post-exposure treatment attenuates noise-induced hearing loss. Neuroscience 2005, 134, 633–642. [Google Scholar] [CrossRef]
- Xu, F.; Cheng, Y.; Yan, W. Up-regulation of autophagy and apoptosis of cochlear hair cells in mouse models for deafness. Arch. Med. Sci. 2018, 17, 535–541. [Google Scholar] [CrossRef]
- Yamasoba, T.; Someya, S.; Yamada, C.; Weindruch, R.; Prolla, T.A.; Tanokura, M. Role of mitochondrial dysfunction and mitochondrial DNA mutations in age-related hearing loss. Hear. Res. 2007, 226, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ruan, R.S. Possible roles of nitric oxide in the physiology and pathophysiology of the mammalian cochlea. Ann. N. Y. Acad. Sci. 2002, 962, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, R.; Yamaguchi, T.; Tanaka, H.; Ogita, K. Mechanism underlying the protective effect of tempol and Nω-nitro-L-arginine methyl ester on acoustic injury: Possible involvement of c-JunNterminal kinase pathway and connexin26 in the cochlear spiral ligament. J. Pharmacol. Sci. 2010, 114, 50–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Ren, T.; Nuttall, A.L. The electrochemical and fluorescence detection of nitric oxide in the cochlea and its increase following loud sound. Hear. Res. 2002, 164, 49–58. [Google Scholar] [CrossRef]
- Honkura, Y.; Matsuo, H.; Murakami, S.; Sakiyama, M.; Mizutari, K.; Shiotani, A.; Yamamoto, M.; Morita, I.; Shinomiya, N.; Kawase, T.; et al. NRF2 Is a Key Target for Prevention of Noise-Induced Hearing Loss by Reducing Oxidative Damage of Cochlea. Sci. Rep. 2016, 6, 19329. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, J.; Fuentes-Santamaría, V.; Gabaldón-Ull, M.; Jareño-Flores, T.; Miller, J.; Juiz, J.M. Noise-Induced “Toughening” Effect in Wistar Rats: Enhanced Auditory Brainstem Responses Are Related to Calretinin and Nitric Oxide Synthase Upregulation. Front. Neuroanat. 2015, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Jamesdaniel, S.; Coling, D.; Hinduja, S.; Ding, D.; Li, J.; Cassidy, L.; Seige, G.M.; Qu, J.; Salvi, R. Cisplatin-induced ototoxicity is mediated by nitroxidative modification of cochlear proteins characterized by nitration of Lmo4. J. Biol. Chem. 2012, 287, 18674–18686. [Google Scholar] [CrossRef] [Green Version]
- Jamesdaniel, S.; Rathinam, R.; Neumann, W. Targeting nitrative stress for attenuating cisplatin-induced downregulation of cochlear LIM domain only 4 and ototoxicity. Redox Biol. 2015, 10, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Jamesdaniel, S. Downstream Targets of Lmo4 Are Modulated by Cisplatin in the Inner Ear of Wistar Rats 2014. PLoS ONE 2014, 9, e115263. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, U.-R.; Selivanova, O.; Feltens, R.; Brieger, J.; Mann, W. Endothelial nitric oxide synthase up-regulation in the guinea pig organ of Corti after acute noise trauma. Brain Res. 2005, 1047, 85–96. [Google Scholar] [CrossRef]
- Jia, Z.; He, Q.; Shun, C.; Li, F. Tauroursodeoxycholic acid attenuates gentamicin-induced cochlear hair cell death in vitro. Toxicol. Lett. 2018, 294, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, R.; Ghosh, S.; Neumann, W.L.; Jamesdaniel, S. Cisplatin-induced apoptosis in auditory, renal, and neuronal cells is associated with nitration and downregulation of LMO4. Cell Death Discov. 2015, 1, 15052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahab, M.; Rosati, R.; Meyers, D.; Sheilds, J.; Crofts, E.; Baker, T.R.; Jamesdaniel, S. Cisplatin-induced hair cell loss in zebrafish neuromasts is accompanied by protein nitration and Lmo4 degradation. Toxicol. Appl. Pharmacol. 2021, 410, 115342. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Talaska, A.E.; Schacht, J.; Sha, S.H. Oxidative imbalance in the aging inner ear. Neurobiol. Aging 2007, 28, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiyama, G.; Wester, J.; Lopez, I.A.; Beltran-Parrazal, L.; Ishiyama, A. Oxidative Stress in the Blood Labyrinthine Barrier in the Macula Utricle of Meniere’s Disease Patients. Front. Physiol. 2018, 9, 1068. [Google Scholar] [CrossRef] [PubMed]
- Vlajkovic, S.M.; Lee, K.; Wong, A.H.; Guo, C.; Gupta, R.; Housley, G.; Thorne, P. Adenosine aminecongener mitigates noise-induced cochlear injury. Purinergic Signal. 2010, 6, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Rosati, R.; Shahab, M.; Neumann, W.; Jamesdaniel, S. Inhibition of protein nitration prevents cisplatin-induced inactivation of STAT3 and promotes anti-apoptotic signaling in organ of Corti cells. Exp. Cell Res. 2019, 381, 105–111. [Google Scholar] [CrossRef]
- Diao, M.; Gao, W.; Sun, J. Nitric oxide synthase inhibitor reduces noise-induced cochlear damage in guinea pigs. Acta Otolaryngol. 2007, 127, 1162–1167. [Google Scholar] [CrossRef]
- Wu, G.; Lee, T.D.; Moore, R.E.; Rao, N.A. Photoreceptor mitochondrial tyrosine nitration in experimental uveitis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2271–2281. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.H.; Li, Y.N.; Qi, J.S.; Jia, X.X. Peroxynitrite-induced protein nitration is responsible for renal mitochondrial damage in diabetic rat. J. Endocrinol. Investig. 2010, 33, 140–146. [Google Scholar] [CrossRef]
- Greenacre, S.A.; Ischiropoulos, H. Tyrosine nitration: Localization, quantification, consequences for protein function and signal transduction. Free Radic. Res. 2001, 34, 541–581. [Google Scholar] [CrossRef] [PubMed]
- Pourbakht, A.; Yamasoba, T. Ebselen attenuates cochlear damage caused by acoustic trauma. Hear. Res. 2005, 181, 100–108. [Google Scholar] [CrossRef]
- Lynch, E.; Gua, R.; Piercea, C.; Kil, J. Combined oral delivery of ebselen and allopurinol reduces multiple cisplatin toxicities in rat breast and ovarian cancer models while enhancing anti-tumor activity. Anticancer Drugs 2005, 16, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Kil, J.; Lobarinas, E.; Spankovich, C.; Griffiths, S.; Antonelli, P.; Lynch, E.; Prell, C. Safety and efficacy of ebselen for the prevention of noise-induced hearing loss: A randomized, double-blind, placebo-controlled, phase 2 trial. Lancet 2017, 390, 969–979. [Google Scholar] [CrossRef]
- Wu, S.B.; Wu, Y.T.; Wu, T.P.; Wei, Y.H. Role of AMPK-mediated adaptive responses in human cells with mitochondrial dysfunction to oxidative stress. Biochem. Biophys. Acta 2020, 1840, 1331–1344. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, C.; Martin-Sanz, E.; Cuadrado, E.; Granizo, J.J.; Sanz-Fernandez, R. Protective effect of polyphenols on presbycusis via oxidative/nitrosative stress suppression in rats. Exp. Gerontol. 2016, 83, 31–36. [Google Scholar] [CrossRef]
- Xiong, M.; Lai, H.; He, Q.; Wang, J. Astragaloside IV attenuates impulse noise-induced trauma in guinea pig. Acta Otolaryngol. 2011, 131, 809–816. [Google Scholar] [CrossRef]
- Yang, D.H.; Xie, J.; Liu, K.; Peng, Z.; Guo, J.Y.; Yu, S.K.; Wang, G.P.; Gong, S.S. The histone deacetylase inhibitor sodium butyrate protects against noise-induced hearing loss in Guinea pigs. Neurosci. Lett. 2017, 660, 140–146. [Google Scholar] [CrossRef]
- Lu, S.; Fan, Z.; Xu, W.; Han, Y.; Zhang, G.; Liu, W.; Bai, X.; Wang, X.; Xin, H.; Li, J.; et al. L-cysteine attenuates peroxynitrite-elicited cytotoxicity to spiral ganglion neurons: Possible relation to hearing loss. Neurol. Res. 2011, 33, 935–941. [Google Scholar] [CrossRef]
- Ciodaro, F.; Freni, F.; Alberti, G.; Forelli, M.; Gazia, F.; Bruno, R.; Sherdell, E.P.; Galletti, B.; Galletti, F. Application of Cervical Vestibular-Evoked Myogenic Potentials in Adults with Moderate to Profound Sensorineural Hearing Loss: A Preliminary Study. Int. Arch. Otorhinolaryngol. 2020, 24, e5–e10. [Google Scholar] [CrossRef] [Green Version]
- Toumpas, C.J.; Clark, J.; Harris, A.; Beswick, R.; Nourse, C.B. Congenital cytomegalovirus infection is a significant cause of moderate to profound sensorineural hearing loss in Queensland children. J. Paediatr. Child Health 2015, 51, 541–544. [Google Scholar] [CrossRef] [PubMed]
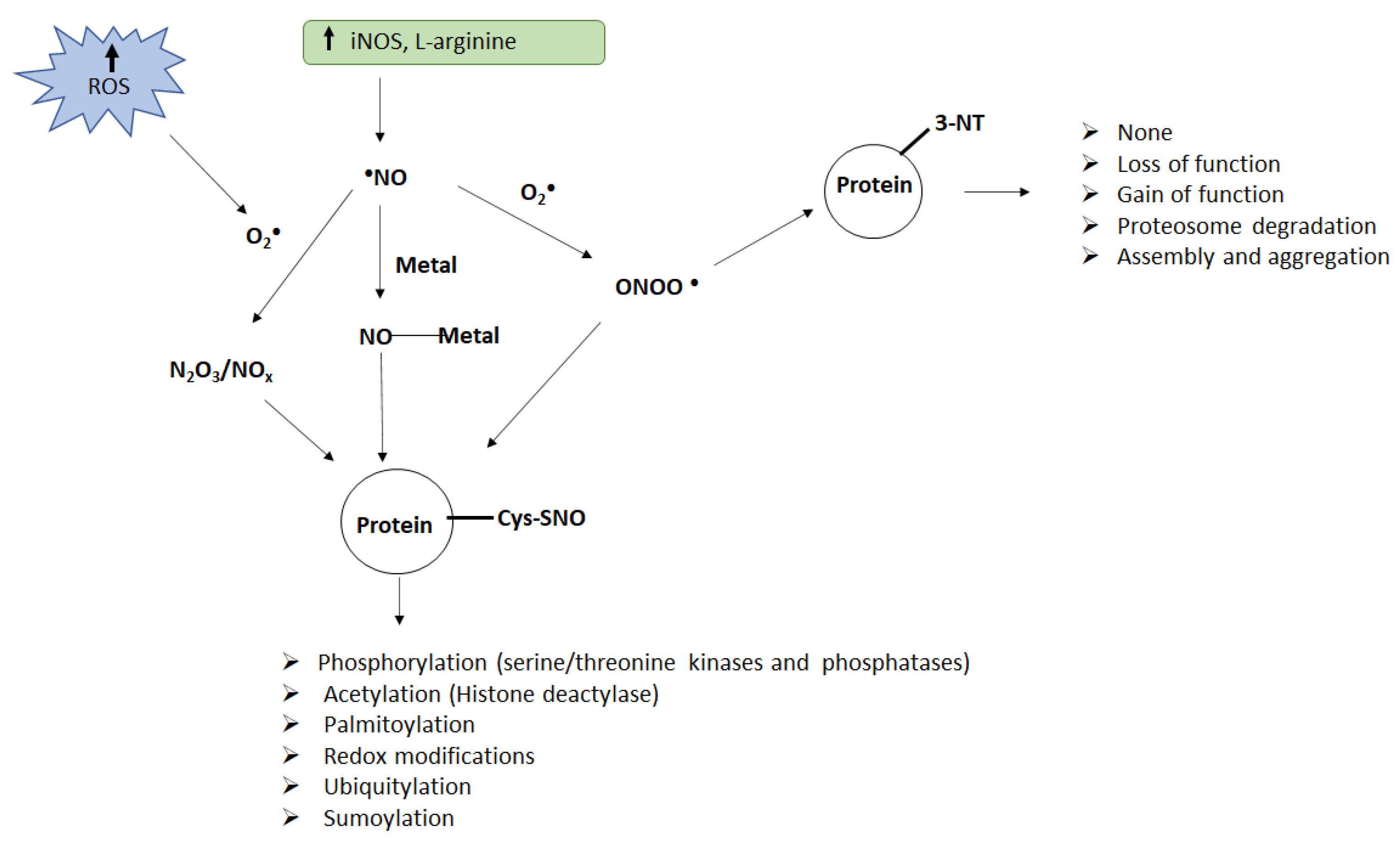
Figure 1.
Schematic of protein nitration and nitrosylation and their consequences. Enhanced ROS generation and accumulation of nitric oxide can lead to the formation of peroxynitrite (ONOO−), NO oxides (N2O3/NOx), or metal–NO complexes (M–NO), which act as nitrating agents in vivo. Peroxynitrite can induce nitration of tyrosine (Tyr) residues on the proteins while peroxynitrite, metal–NO complexes, or NO oxides can induce nitrosylation of cysteine (Cys) residues on the proteins. Both nitration and nitrosylation can alter protein function resulting in several adverse consequences.
Figure 1.
Schematic of protein nitration and nitrosylation and their consequences. Enhanced ROS generation and accumulation of nitric oxide can lead to the formation of peroxynitrite (ONOO−), NO oxides (N2O3/NOx), or metal–NO complexes (M–NO), which act as nitrating agents in vivo. Peroxynitrite can induce nitration of tyrosine (Tyr) residues on the proteins while peroxynitrite, metal–NO complexes, or NO oxides can induce nitrosylation of cysteine (Cys) residues on the proteins. Both nitration and nitrosylation can alter protein function resulting in several adverse consequences.


{kind=link}
Table 1.
Studies showing nitrative stress in auditory dysfunction.
| Author/Year | Model (Animal/Cell Culture) | Nitrative Stress Molecule | Ototraumatic Agent | Region Affected in Inner Ear |
|---|---|---|---|---|
| Alvarado et al., 2015 | Wistar rats | nNOS | Noise | Spiral ganglion, spiral ligament, and cochlear nerves |
| Han et al., 2013 | Guinea pigs | 3-NT | Noise | OHCs, IHCs, spiral ganglion, and pillar cells |
| Jamesdaniel et al., 2012 | UBOC1 cell lines | 3-NT | Cisplatin | UBOC1 cells |
| Jamesdaniel, 2014 | Wistar rats | 3-NT | Cisplatin | Spiral ganglion, stria vascularis, and organ of Corti |
| Jamesdaniel et al., 2016 | UBOC1 cell lines | 3-NT | Cisplatin | UBOC1 cells |
| Jiang et al., 2007 | CBA/J mice | 3-NT, OONO− | Aging | Deiters’ cells, pillar cells, stria vascularis, spiral ganglion |
| Lu et al., 2011 | Rats | 3-NT | OONO- | Spiral ganglion neurons |
| Lynch et al., 2005 | Female rats F-344 | 3-NT | Cisplatin | OHCs |
| Nagashima et al., 2010 | Std-ddY mice | 3-NT | Noise | Spiral ligament |
| Pourbakht et al., 2005 | Guinea pigs | 3-NT | Noise | OHCs |
| Rathinam et al., 2015 | UBOC1, HK-2, and SH-SY5Y cell lines | 3-NT | Cisplatin | UBOC1, HK-2, and SH-SY5Y cells |
| Rosati et al., 2019 | UBOC1 cell lines | 3-NT | Cisplatin | UBOC1 cells |
| Shahab et al., 2020 | Zebrafish | 3-NT | Cisplatin | Hair cells in neuromast |
| Shi et al., 2002 | Guinea pigs | 3-NT | Noise | OHCs |
| Vlajkovic et al., 2010 | Wistar rats | 3-NT | Noise | Organ of Corti, spiral ganglion neurons, spiral ligament, spiral limbus, inner sulcus cells, inner phalangeal cells, pillar cells, Deiters’ cells, Hensen’s cells |
| Wu et al., 2020 | CBA/J mice and HEI-OC1 cells | 3-NT | Noise | OHCs |
| Xiong et al., 2011 | Guinea pigs | 3-NT and iNOS synthase | Noise | Stria vascularis, spiral ligament, organ of Corti |
| Yamasoba et al., 2007 | Guinea pigs | NO | Noise | Organ of Corti, afferent dendrites beneath IHCs |
| Yamashita et al., 2004 | Guinea pigs | 3-NT | Noise | Spiral ganglion, organ of Corti, lateral wall |
| Yamashita et al., 2005 | Guinea pigs | 3-NT | Noise | OHCs |
| Yang et al., 2017 | Guinea pigs | 3-NT | Noise | OHCs |
| Yuan et al., 2015 | CBA/J mice | 3-NT | Noise | IHCs and OHCs |
Table 2.
Studies showing inhibition of nitration and attenuated hearing loss and auditory dysfunction.
Table 2.
Studies showing inhibition of nitration and attenuated hearing loss and auditory dysfunction.
| Author/Year | Model (Animal/Cell Culture) | Inhibitors of Nitration | Biochemical Effect of Intervention | Outcome of Intervention |
|---|---|---|---|---|
| Diao et al., 2007 | Guinea pigs | NG-Nitro-L-arginine methyl ester (L-NAME) | Decreased NO production | L-NAME protected the cochlea after noise exposure |
| Jamesdaniel et al., 2012 | Wistar rats | Trolox | Decreased nitration of cochlear proteins | Attenuated cisplatin-induced OHC loss and hearing threshold shifts |
| Jamesdaniel, 2014 | Wistar rats | Trolox | Decreased nitration of cochlear proteins | Attenuated cisplatin-induced ototoxicity and nitration of cochlear proteins |
| Jamesdaniel et al., 2016 | UBOC1 cell lines | SRI110 | Decreased 3-NT | SRI110 inhibited cisplatin-induced cytotoxicity |
| Jia et al., 2018 | HEI-OC1 cells and explanted cochlear tissue | Tauroursodeoxycholic acid (TUDCA) | Decreased NO production | SRI110 inhibited cisplatin-induced cytotoxicity |
| Lu et al., 2011 | Rats | L-Cysteine | Decreased peroxynitrite | Decreased gentamicin-induced ototoxicity |
| Lyncha et al., 2005 | Fisher-344 rats | Ebselen | Decreased lipid peroxidation | Protected cochlea from cisplatin-induced OHC loss and hearing loss |
| Nagashima et al., 2010 | Std-ddY mice | Tempol and Nω-nitro-L-arginine methyl ester | Decreased 3-NT and 4-HNE | Protected noise exposed animals from hearing loss |
| Pourbakht et al., 2005 | Guinea pigs | Ebselen | Decreased peroxynitrite | Reduced the noise-induced permanent threshold shifts |
| Rosati et al., 2019 | UBOC1 cell lines | SRI110 | Decreased 3-NT | Prevented cisplatin-induced cytotoxicity |
| Sánchez-Rodríguez et al., 2016 | Sprague-Dawley Rats | Polyphenols | Decreased 3-NT and superoxide anions, increased glutathione peroxidase and SOD | Protected against age-related hearing loss |
| Wu et al., 2020 | CBA/J mice and HEI-OC1 cells | N-Acetyl cysteine (NAC) | Decreased 3-NT and superoxides | Prevented loss of OHCs induced by noise exposure |
| Vlajkovic et al., 2010 | Wistar rats | Adenosine amine congener (ADAC) | Decreased 3-NT | Attenuated hearing threshold shifts after noise exposure |
| Xiong et al., 2011 | Guinea pigs | Astragaloside IV | Inhibited iNOS | Protected the cochlea from noise-induced damage |
| Yamashita et al., 2005 | Guinea pigs | Salicylate and trolox | Decreased 3-NT and superoxides | Reduced ABR shifts post noise exposure |
| Yamasoba et al., 2005 | Guinea pigs | Ebselen | Decreased NO production | Reduced ABR shifts post noise exposure |
| Yang et al., 2017 | Guinea pigs | Sodium butyrate (SB) | Decreased 3-NT and HDAC1 | Attenuated noise-induced loss of OHC and hearing loss |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shahab, M.; Jamesdaniel, S. Nitrative Stress and Auditory Dysfunction. Pharmaceuticals 2022, 15, 649. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15060649
AMA Style
Shahab M, Jamesdaniel S. Nitrative Stress and Auditory Dysfunction. Pharmaceuticals. 2022; 15(6):649. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15060649
Chicago/Turabian StyleShahab, Monazza, and Samson Jamesdaniel. 2022. "Nitrative Stress and Auditory Dysfunction" Pharmaceuticals 15, no. 6: 649. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15060649
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.