
Toll-Like Receptor and Cytokine Responses to Infection with Endogenous and Exogenous Koala Retrovirus, and Vaccination as a Control Strategy


Abstract
:1. Introduction
2. Health Effects of KoRV on Koalas
3. Innate Immune Response to KoRV Infection in Koalas
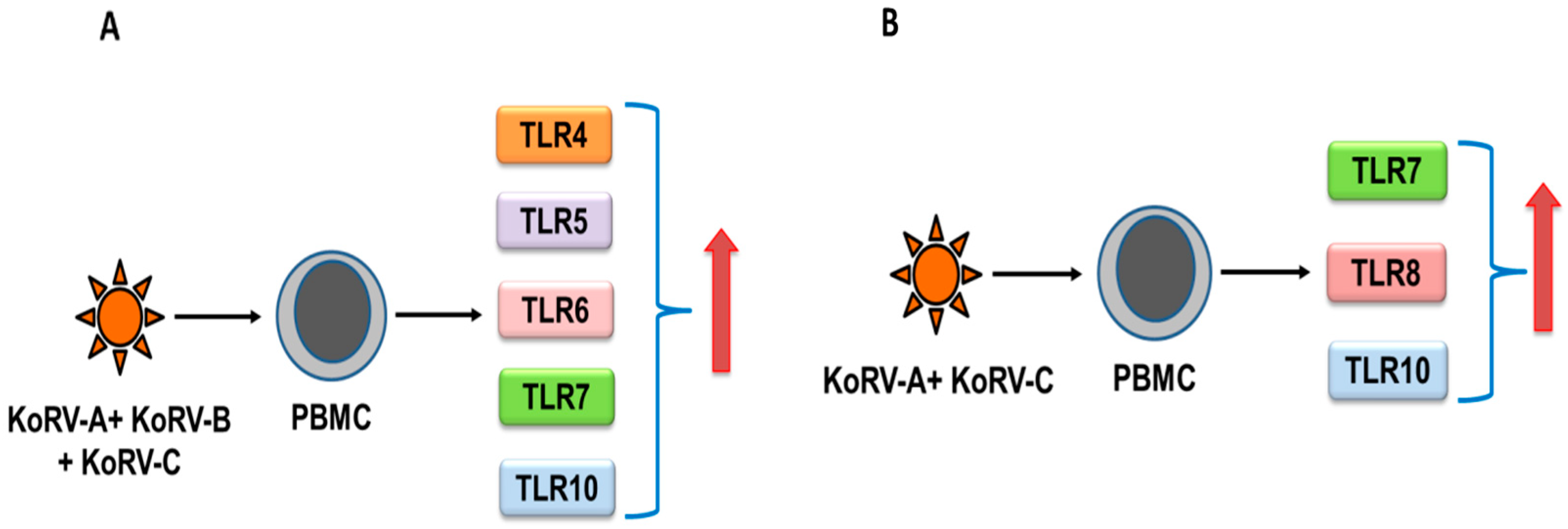
3.1. Toll-Like Receptors (TLRs)
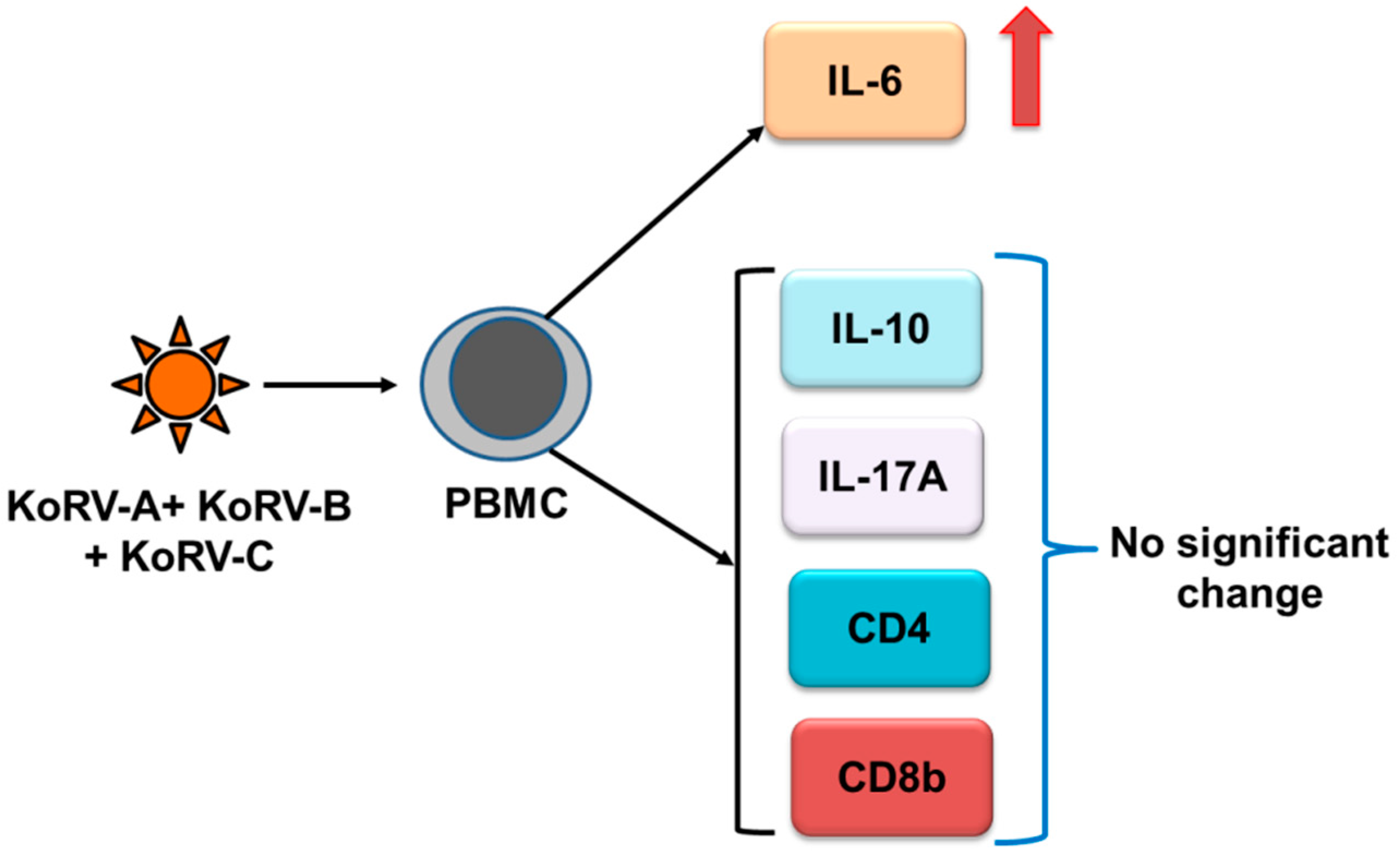
3.2. Cytokines
4. Maintenance of Koala Health and Conservation by Vaccine Development and the Investigation of Alternative Strategies
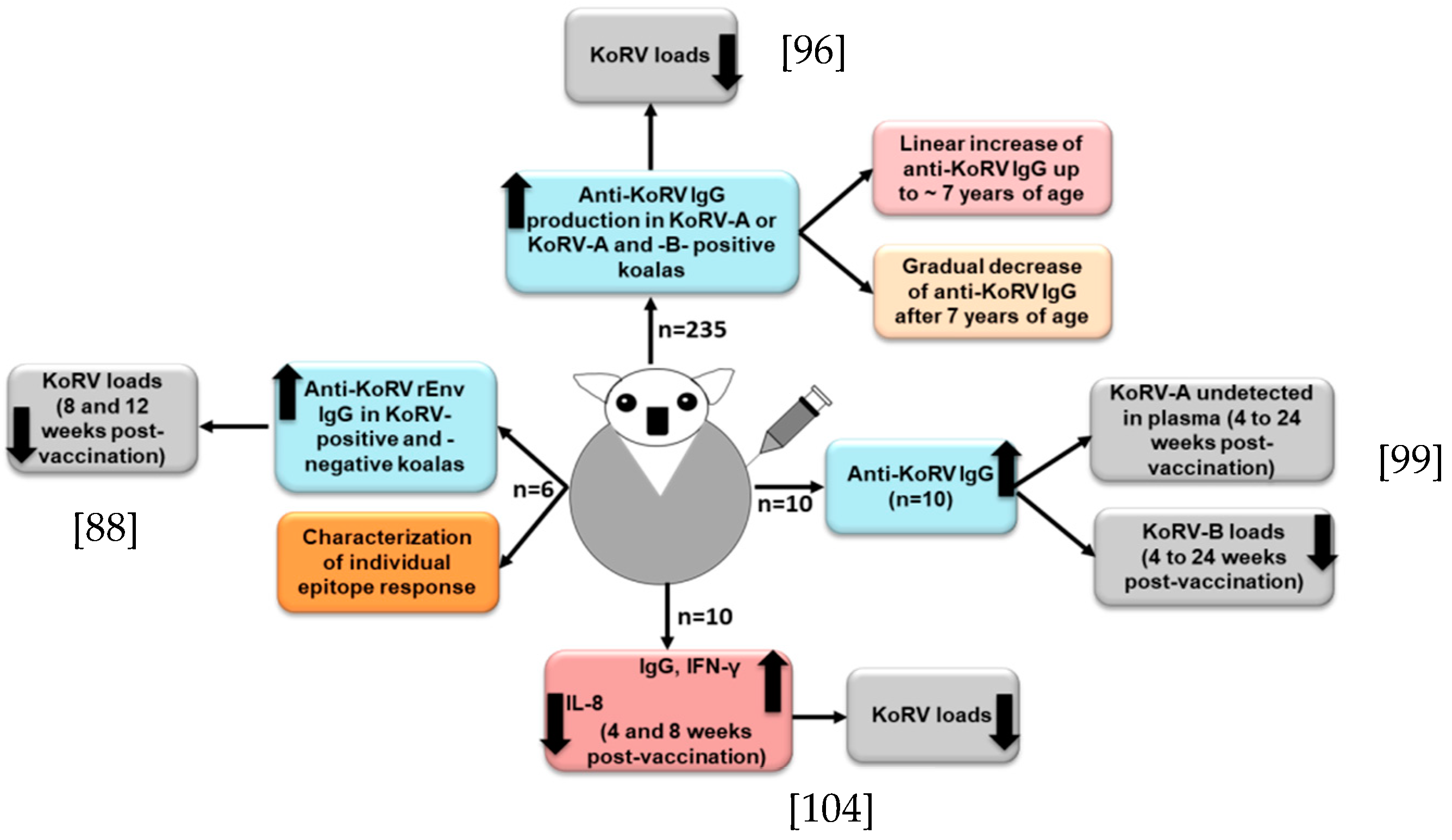
4.1. Vaccine Response in Koalas against KoRV
4.2. Alternative Strategies to Control KoRV Infection
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Melzer, A.; Carrick, F.; Menkhorst, P.; Lunney, D.; John, B.S. Overview, critical assessment, and conservation implications of koala distribution and abundance. Conserv. Biol. 2000, 14, 619–628. [Google Scholar] [CrossRef]
- Quigley, B.L.; Timms, P. Helping koalas battle disease–Recent advances in Chlamydia and koala retrovirus (KoRV) disease understanding and treatment in koalas. FEMS Microbiol. Rev. 2020, 44, 583–605. [Google Scholar] [CrossRef] [PubMed]
- Connolly, J.H.; Canfield, P.J.; Hemsley, S.; Spencer, A.J. Lymphoid neoplasia in the koala. Aust. Vet. J. 1998, 76, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Tarlinton, R.; Meers, J.; Hanger, J.; Young, P. Real-time reverse transcriptase PCR for the endogenous koala retrovirus reveals an association between plasma viral load and neoplastic disease in koalas. J. Gen. Virol. 2005, 86, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Stadler, C.K.; Gorman, K.; Jensen, N.; Kim, D.; Zheng, H.Q.; Tang, S.; Switzer, W.M.; Pye, G.W.; Eiden, M.V. An exogenous retrovirus isolated from koalas with malignant neoplasias in a US zoo. Proc. Natl. Acad. Sci. USA 2013, 110, 11547–11552. [Google Scholar] [CrossRef] [Green Version]
- Waugh, C.A.; Hanger, J.; Loader, J.; King, A.; Hobbs, M.; Johnson, R.; Timms, P. Infection with koala retrovirus subgroup B (KoRV-B), but not KoRV-A, is associated with chlamydial disease in free-ranging koalas (Phascolarctos cinereus). Sci. Rep. 2017, 7, 134. [Google Scholar] [CrossRef] [Green Version]
- Fabijan, J.; Woolford, L.; Lathe, S.; Simmons, G.; Hemmatzadeh, F.; Trott, D.J.; Speight, K.N. Lymphoma, koala retrovirus infection and reproductive chlamydiosis in a koala (Phascolarctos cinereus). J. Comp. Pathol. 2017, 157, 188–192. [Google Scholar] [CrossRef]
- Quigley, B.L.; Phillips, S.; Olagoke, O.; Robbins, A.; Hanger, J.; Timms, P. Changes in endogenous and exogenous koala retrovirus subtype expression over time reflect koala health outcomes. J. Virol. 2019, 93, e00849-19. [Google Scholar] [CrossRef] [Green Version]
- Hashem, M.A.; Kayesh, M.E.H.; Yamato, O.; Maetani, F.; Eiei, T.; Mochizuki, K.; Sakurai, H.; Ito, A.; Kannno, H.; Kohara, K.T.; et al. Coinfection with koala retrovirus subtypes A and B and its impact on captive koalas in Japanese zoos. Arch. Virol. 2019, 164, 2735–2745. [Google Scholar] [CrossRef]
- Zheng, H.; Pan, Y.; Tang, S.; Pye, G.W.; Stadler, C.K.; Vogelnest, L.; Herrin, K.V.; Rideout, B.A.; Switzer, W.M. Koala retrovirus diversity, transmissibility, and disease associations. Retrovirology 2020, 17, 34. [Google Scholar] [CrossRef]
- Hashem, M.A.; Kayesh, M.E.H.; Maetani, F.; Eiei, T.; Mochizuki, K.; Ochiai, S.; Ito, A.; Ito, N.; Sakurai, H.; Asai, T.; et al. Koala retrovirus (KoRV) subtypes and their impact on captive koala (Phascolarctos cinereus) health. Arch. Virol. 2021. (Accepted). [Google Scholar] [CrossRef] [PubMed]
- Hanger, J.J.; Bromham, L.D.; McKee, J.J.; O’Brien, T.M.; Robinson, W.F. The nucleotide sequence of Koala (Phascolarctos cinereus) retrovirus: A novel type C endogenous virus related to Gibbon ape leukemia virus. J. Virol. 2000, 74, 4264–4272. [Google Scholar] [CrossRef] [Green Version]
- Tarlinton, R.E.; Meers, J.; Young, P.R. Retroviral invasion of the koala genome. Nature 2006, 442, 79–81. [Google Scholar] [CrossRef]
- Denner, J.; Young, P.R. Koala retroviruses: Characterization and impact on the life of koalas. Retrovirology 2013, 10, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobbs, M.; King, A.; Salinas, R.; Chen, Z.; Tsangaras, K.; Greenwood, A.D.; Johnson, R.N.; Belov, K.; Wilkins, M.R.; Timms, P. Long-read genome sequence assembly provides insight into ongoing retroviral invasion of the koala germline. Sci. Rep. 2017, 7, 15838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, Y.; Zhao, K.; Greenwood, A.D.; Roca, A.L. Proliferation of endogenous retroviruses in the early stages of a host germ line invasion. Mol. Biol. Evol. 2015, 32, 109–120. [Google Scholar] [CrossRef]
- Yu, T.; Koppetsch, B.S.; Pagliarani, S.; Johnston, S.; Silverstein, N.J.; Luban, J.; Chappell, K.; Weng, Z.; Theurkauf, W.E. The piRNA Response to Retroviral Invasion of the Koala Genome. Cell 2019, 179, 632–643.e12. [Google Scholar] [CrossRef]
- Alfano, N.; Michaux, J.; Fabre, P.H.; Morand, S.; Alpin, K.; Tsangaras, K.; Löber, U.; Fitriana, Y.; Semiadi, G.; Ishida, Y.; et al. Endogenous gibbon ape leukemia virus identified in a rodent (Melomys burtoni subsp.) from Wallacea (Indonesia). J. Virol. 2016, 90, 8169–8180. [Google Scholar] [CrossRef] [Green Version]
- Simmons, G.; Clarke, D.; McKee, J.; Young, P.; Meers, J. Discovery of a novel retrovirus sequence in an Australian native rodent (Melomys burtoni): A putative link between gibbon ape leukemia virus and koala retrovirus. PLoS ONE 2014, 9, e106954. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, A.D.; Ishida, Y.; O’Brien, S.P.; Roca, A.L.; Eiden, M.V. Transmission, Evolution, and Endogenization: Lessons Learned from Recent Retroviral Invasions. Microbiol. Mol. Biol. Rev. 2017, 82, e00044-17. [Google Scholar] [CrossRef] [Green Version]
- Hayward, J.A.; Tachedjian, M.; Kohl, C.; Johnson, A.; Dearnley, M.; Jesaveluk, B.; Langer, C.; Solymosi, P.D.; Hille, G.; Nitsche, A.; et al. Infectious KoRV-related retroviruses circulating in Australian bats. Proc. Natl. Acad. Sci. USA 2020, 117, 9529–9536. [Google Scholar] [CrossRef] [Green Version]
- Kayesh, M.E.H.; Hashem, M.A.; Tsukiyama-Kohara, K. Koala retrovirus epidemiology, transmission mode, pathogenesis, and host immune response in koalas (Phascolarctos cinereus): A review. Arch. Virol. 2020, 165, 2409–2417. [Google Scholar] [CrossRef]
- Oliveira, N.M.; Farrell, K.B.; Eiden, M.V. In vitro characterization of a koala retrovirus. J. Virol. 2006, 80, 3104–3107. [Google Scholar] [CrossRef] [Green Version]
- Shojima, T.; Yoshikawa, R.; Hoshino, S.; Shimode, S.; Nakagawa, S.; Ohata, T.; Nakaoka, R.; Miyazawa, T. Identification of a novel subgroup of Koala retrovirus from Koalas in Japanese zoos. J. Virol. 2013, 87, 9943–9948. [Google Scholar] [CrossRef] [Green Version]
- Simmons, G.S.; Young, P.R.; Hanger, J.J.; Jones, K.; Clarke, D.; McKee, J.J.; Meers, J. Prevalence of koala retrovirus in geographically diverse populations in Australia. Aust. Vet. J. 2012, 90, 404–409. [Google Scholar] [CrossRef]
- Fabijan, J.; Miller, D.; Olagoke, O.; Woolford, L.; Boardman, W.; Timms, P.; Polkinghorne, A.; Simmons, G.; Hemmatzadeh, F.; Trott, D.J.; et al. Prevalence and clinical significance of koala retrovirus in two South Australian koala (Phascolarctos cinereus) populations. J. Med. Microbiol. 2019, 68, 1072–1080. [Google Scholar] [CrossRef]
- Sarker, N.; Fabijan, J.; Seddon, J.; Tarlinton, R.; Owen, H.; Simmons, G.; Thia, J.; Blanchard, A.M.; Speight, N.; Kaler, J.; et al. Genetic diversity of Koala retrovirus env gene subtypes: Insights into northern and southern koala populations. J. Gen. Virol. 2019, 100, 1328–1339. [Google Scholar] [CrossRef] [PubMed]
- Chiu, E.S.; VandeWoude, S. Endogenous Retroviruses Drive Resistance and Promotion of Exogenous Retroviral Homologs. Annu. Rev. Anim. Biosci. 2021, 9, 225–248. [Google Scholar] [CrossRef]
- Löber, U.; Hobbs, M.; Dayaram, A.; Tsangaras, K.; Jones, K.; Alquezar-Planas, D.E.; Ishida, Y.; Meers, J.; Mayer, J.; Quedenau, C.; et al. Degradation and remobilization of endogenous retroviruses by recombination during the earliest stages of a germ-line invasion. Proc. Natl. Acad. Sci. USA 2018, 115, 8609–8614. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.N.; O’Meally, D.; Chen, Z.; Etherington, G.J.; Ho, S.Y.W.; Nash, W.J.; Grueber, C.E.; Cheng, Y.; Whittington, C.M.; Dennison, S.; et al. Adaptation and conservation insights from the koala genome. Nat. Genet. 2018, 50, 1102–1111. [Google Scholar] [CrossRef]
- Stoye, J.P. Koala retrovirus: A genome invasion in real time. Genome Biol. 2006, 7, 241. [Google Scholar] [CrossRef] [Green Version]
- Denner, J. Transspecies transmissions of retroviruses: New cases. Virology 2007, 369, 229–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarlinton, R.; Meers, J.; Young, P. Biology and evolution of the endogenous koala retrovirus. Cell. Mol. Life Sci. 2008, 65, 3413–3421. [Google Scholar] [CrossRef]
- Xu, W.; Eiden, M.V. Koala Retroviruses: Evolution and Disease Dynamics. Annu. Rev. Virol. 2015, 2, 119–134. [Google Scholar] [CrossRef]
- Kinney, M.E.; Pye, G.W. KOALA RETROVIRUS: A REVIEW. J. Zoo Wildl. Med. 2016, 47, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Madden, D.; Whaite, A.; Jones, E.; Belov, K.; Timms, P.; Polkinghorne, A. Koala immunology and infectious diseases: How much can the koala bear? Dev. Comp. Immunol. 2018, 82, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.P.; Lau, Q.; Maher, I. Koala Immunology and the Koala Retrovirus (KoRV). In The Koala and Its Retroviruses: Implications for Sustainability and Survival; Pye, G.W., Johnson, R.N., Greenwood, A.D., Eds.; Australian Museum: Sydney, Australia, 2014; Volume 24, pp. 35–38. [Google Scholar]
- Bendinelli, M.; Matteucci, D.; Friedman, H. Retrovirus-induced acquired immunodeficiencies. Adv. Cancer Res. 1985, 45, 125–181. [Google Scholar]
- Bishop, J.M. The molecular genetics of cancer. Science 1987, 235, 305–311. [Google Scholar] [CrossRef]
- Fiebig, U.; Hartmann, M.G.; Bannert, N.; Kurth, R.; Denner, J. Transspecies transmission of the endogenous koala retrovirus. J. Virol. 2006, 80, 5651–5654. [Google Scholar] [CrossRef] [Green Version]
- Maher, I.E.; Griffith, J.E.; Lau, Q.; Reeves, T.; Higgins, D.P. Expression profiles of the immune genes CD4, CD8β, IFNγ, IL-4, IL-6 and IL-10 in mitogen-stimulated koala lymphocytes (Phascolarctos cinereus) by qRT-PCR. Peer J. 2014, 2, e280. [Google Scholar] [CrossRef] [Green Version]
- Maher, I.E.; Higgins, D.P. Altered Immune Cytokine Expression Associated with KoRV B Infection and Season in Captive Koalas. PLoS ONE 2016, 11, e0163780. [Google Scholar] [CrossRef] [Green Version]
- Maher, I.E.; Patterson, J.; Curnick, M.; Devlin, J.; Higgins, D.P. Altered immune parameters associated with koala retrovirus (KoRV) and chlamydial infection in free ranging Victorian koalas (Phascolarctos cinereus). Sci. Rep. 2019, 9, 11170. [Google Scholar] [CrossRef] [Green Version]
- Fabijan, J.; Sarker, N.; Speight, N.; Owen, H.; Meers, J.; Simmons, G.; Seddon, J.; Emes, R.D.; Tarlinton, R.; Hemmatzadeh, F.; et al. Pathological findings in koala retrovirus-positive koalas (Phascolarctos cinereus) from Northern and Southern Australia. J. Comp. Pathol. 2020, 176, 50–66. [Google Scholar] [CrossRef]
- Sarker, N.; Fabijan, J.; Owen, H.; Seddon, J.; Simmons, G.; Speight, N.; Kaler, J.; Woolford, L.; Emes, R.D.; Hemmatzadeh, F.; et al. Koala retrovirus viral load and disease burden in distinct northern and southern koala populations. Sci. Rep. 2020, 10, 263. [Google Scholar] [CrossRef]
- Butcher, R.G.; Pettett, L.M.; Fabijan, J.; Ebrahimie, E.; Mohammadi-Dehcheshmeh, M.; Speight, K.N.; Boardman, W.; Bird, P.S.; Trott, D.J. Periodontal disease in free-ranging koalas (Phascolarctos cinereus) from the Mount Lofty Ranges, South Australia, and its association with koala retrovirus infection. Aust. Vet. J. 2020, 98, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Sarker, N.; Tarlinton, R.; Owen, H.; David Emes, R.; Seddon, J.; Simmons, G.; Meers, J. Novel insights into viral infection and oncogenesis from koala retrovirus (KoRV) infection of HEK293T cells. Gene 2020, 733, 144366. [Google Scholar] [CrossRef]
- McEwen, G.K.; Alquezar-Planas, D.E.; Dayaram, A.; Gillett, A.; Tarlinton, R.; Mongan, N.; Chappell, K.J.; Henning, J.; Tan, M.; Timms, P.; et al. Retroviral integrations contribute to elevated host cancer rates during germline invasion. Nat. Commun. 2021, 12, 1316. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, E.I.; Macal, M.; Lewis, G.M.; Harker, J.A. Innate and adaptive immune regulation during chronic viral infections. Annu. Rev. Virol. 2015, 2, 573–597. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, M.; Pavasovic, A.; King, A.G.; Prentis, P.J.; Eldridge, M.D.; Chen, Z.; Colgan, D.J.; Polkinghorne, A.; Wilkins, M.R.; Flanagan, C.; et al. A transcriptome resource for the koala (Phascolarctos cinereus): Insights into koala retrovirus transcription and sequence diversity. BMC Genomics 2014, 15, 786. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R.; Janeway, C., Jr. Innate immunity. N. Engl. J. Med. 2000, 343, 338–344. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol. 2001, 2, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like receptors and the control of immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Kaisho, T.; Akira, S. Toll-like receptor function and signaling. J. Allergy Clin. Immun. 2006, 117, 979–987. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Frankham, G.J.; Johnson, R.N.; Polkinghorne, A.; Timms, P.; O’Meally, D.; Cheng, Y.; Belov, K. SNP marker Discovery in koala TLR genes. PLoS ONE 2015, 10, e0121068. [Google Scholar]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Celhar, T.; Magalhaes, R.; Fairhurst, A.M. TLR7 and TLR9 in SLE: When sensing self goes wrong. Immunol. Res. 2012, 53, 58–77. [Google Scholar] [CrossRef]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [Green Version]
- McCartney, S.A.; Colonna, M. Viral sensors: Diversity in pathogen recognition. Immunol. Rev. 2009, 227, 87–94. [Google Scholar] [CrossRef]
- Campos, M.A.; Almeida, I.C.; Takeuchi, O.; Akira, S.; Valente, E.P.; Procópio, D.O.; Travassos, L.R.; Smith, J.A.; Golenbock, D.T.; Gazzinelli, R.T. Activation of Toll-like receptor-2 by glycosylphosphatidylinositol anchors from a protozoan parasite. J. Immunol. 2001, 167, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.K.; Mullen, G.E.; Leifer, C.A.; Mazzoni, A.; Davies, D.R.; Segal, D.M. Leucine-rich repeats and pathogen recognition in Toll-like receptors. Trends Immunol. 2003, 24, 528–533. [Google Scholar] [CrossRef]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.Y.; Van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in Tlr4 gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef] [Green Version]
- Oldenburg, M.; Krüger, A.; Ferstl, R.; Kaufmann, A.; Nees, G.; Sigmund, A.; Bathke, B.; Lauterbach, H.; Suter, M.; Dreher, S.; et al. TLR13 Recognizes Bacterial 23S rRNA Devoid of Erythromycin Resistance–Forming Modification. Science 2012, 337, 1111–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, A.V. The genomics and genetics of human infectious disease susceptibility. Annu. Rev. Genom. Hum. Genet. 2001, 2, 373–400. [Google Scholar] [CrossRef]
- Taylor, B.D.; Darville, T.; Ferrell, R.E.; Kammerer, C.M.; Ness, R.B.; Haggerty, C.L. Variants in toll-like receptor 1 and 4 genes are associated with Chlamydia trachomatis among women with pelvic inflammatory disease. J. Infect. Dis. 2012, 205, 603–609. [Google Scholar]
- Herniou, E.; Martin, J.; Miller, K.; Cook, J.; Wilkinson, M.; Tristem, M. Retroviral diversity and distribution in verte-brates. J. Virol. 1998, 72, 5955–5966. [Google Scholar] [CrossRef] [Green Version]
- Browne, E.P. The Role of Toll-like Receptors in Retroviral Infection. Microorganisms 2020, 8, 1787. [Google Scholar] [CrossRef]
- Salaun, B.; Romero, P.; Lebecque, S. Toll-like receptors’ two-edged sword: When immunity meets apoptosis. Eur. J. Immunol. 2007, 37, 3311–3318. [Google Scholar] [CrossRef]
- Huang, B.; Zhao, J.; Unkeless, J.C.; Feng, Z.H.; Xiong, H. TLR signaling by tumor and immune cells: A double-edged sword. Oncogene 2008, 27, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Basith, S.; Manavalan, B.; Yoo, T.H.; Kim, S.G.; Choi, S. Roles of toll-like receptors in Cancer: A double-edged sword for defense and offense. Arch. Pharmacal Res. 2012, 35, 1297–1316. [Google Scholar] [CrossRef]
- Kayesh, M.E.H.; Hashem, M.A.; Tsukiyama-Kohara, K. Toll-like receptor expression profiles in koala (Phascolarctos cinereus) peripheral blood mononuclear cells infected with multiple KoRV subtypes. Animals 2021, 11, 983. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Bryant, C.E.; Doyle, S.L. Therapeutic Targeting of Toll-Like Receptors for Infectious and Inflammatory Diseases and Cancer. Pharmacol. Rev. 2009, 61, 177–197. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, J.J.; Gadina, M.; Siegel, R.M. Cytokines and cytokine receptors. In Clinical Immunology, 5th ed.; Rich, R.R., Fleisher, T.A., Shearer, W.T., Schroeder, H.W., Frew, A.J., Weyand, C.M., Eds.; Content Repository Only: London, UK, 2019; pp. 127–155.e1. [Google Scholar]
- Mogensen, T.H.; Paludan, S.R. Molecular pathways in virus-induced cytokine production. Microbiol. Mol. Biol. Rev. 2001, 65, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, O.; Akira, S. MDA5/RIG-I and virus recognition. Curr. Opin. Immunol. 2008, 20, 17–22. [Google Scholar] [CrossRef]
- Carey, A.J.; Timms, P.; Rawlinson, G.; Brumm, J.; Nilsson, K.; Harris, J.M.; Beagley, K.W. A multi-subunit chlamydial vaccine induces antibody and cell-mediated immunity in immunized koalas (Phascolarctos cinereus): Comparison of three different adjuvants. Am. J. Reprod. Immunol. 2010, 63, 161–172. [Google Scholar] [CrossRef]
- Kollipara, A.; George, C.; Hanger, J.; Loader, J.; Polkinghorne, A.; Beagley, K.; Timms, P. Vaccination of healthy and diseased koalas (Phascolarctos cinereus) with a Chlamydia pecorum multi-subunit vaccine: Evaluation of immunity and pathology. Vaccine 2012, 30, 1875–1885. [Google Scholar] [CrossRef] [Green Version]
- Kayesh, M.E.H.; Hashem, M.A.; Maetani, F.; Eiei, T.; Mochizuki, K.; Ochiai, S.; Ito, A.; Ito, N.; Sakurai, H.; Asai, T.; et al. CD4, CD8b, and Cytokines Expression Profiles in Peripheral Blood Mononuclear Cells Infected with Different Subtypes of KoRV from Koalas (Phascolarctos cinereus) in a Japanese Zoo. Viruses 2020, 12, 1415. [Google Scholar] [CrossRef]
- Woinarski, J.; Burbidge, A.A. Phascolarctos cinereus (amended version of 2016 assessment). The IUCN Red List of Threatened Species 2020: E.T16892A166496779. 2020. Available online: https://www.iucnredlist.org/species/16892/166496779 (accessed on 16 January 2021).
- Hilleman, M.R. Vaccines in historic evolution and perspective: A narrative of vaccine discoveries. Vaccine 2000, 18, 1436–1447. [Google Scholar] [CrossRef]
- Graham, B.S. Advances in antiviral vaccine development. Immunol. Rev. 2013, 255, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Olagoke, O.; Miller, D.; Hemmatzadeh, F.; Stephenson, T.; Fabijan, J.; Hutt, P.; Finch, S.; Speight, N.; Timms, P. Induction of neutralizing antibody response against koala retrovirus (KoRV) and reduction in viral load in koalas following vaccination with recombinant KoRV envelope protein. NPJ Vaccines 2018, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Vaine, M.; Lu, S.; Wang, S. Progress on the induction of neutralizing antibodies against HIV type 1 (HIV-1). BioDrugs 2009, 23, 137–153. [Google Scholar] [CrossRef] [Green Version]
- Osterhaus, A.; Weijer, K.; UytdeHaag, F.; Knell, P.; Jarrett, O.; Akerblom, L.; Morein, B. Serological responses in cats vaccinated with FeLV ISCOM and an inactivated FeLV vaccine. Vaccine 1989, 7, 137–141. [Google Scholar] [CrossRef]
- Fiebig, U.; Keller, M.; Möller, A.; Timms, P.; Denner, J. Lack of antiviral antibody response in koalas infected with koala retroviruses (KoRV). Virus Res. 2015, 198, 30–34. [Google Scholar] [CrossRef]
- Fiebig, U.; Dieckhoff, B.; Wurzbacher, C.; Möller, A.; Kurth, R.; Denner, J. Induction of neutralizing antibodies specific for the envelope proteins of the koala retrovirus by immunization with recombinant proteins or with DNA. Virol. J. 2015, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Waugh, C.; Gillett, A.; Polkinghorne, A.; Timms, P. Serum antibody response to koala retrovirus antigens varies in free-ranging koalas (Phascolarctos cinereus) in Australia: Implications for vaccine design. J. Wildl. Dis. 2016, 52, 422–425. [Google Scholar] [CrossRef]
- Montero, M.; van Houten, N.E.; Wang, X.; Scott, J.K. The membrane-proximal external region of the human immunodeficiency virus type 1 envelope: Dominant site of antibody neutralization and target for vaccine design. Microbiol. Mol. Biol. Rev. 2008, 72, 54–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinos-Albert, L.M.; Clotet, B.; Blanco, J.; Carrillo, J. Immunologic Insights on the Membrane Proximal External Region: A Major Human Immunodeficiency Virus Type-1 Vaccine Target. Front. Immunol. 2017, 8, 1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olagoke, O.; Quigley, B.L.; Eiden, M.V.; Timms, P. Antibody response against koala retrovirus (KoRV) in koalas harboring KoRV-A in the presence or absence of KoRV-B. Sci. Rep. 2019, 9, 12416. [Google Scholar] [CrossRef] [Green Version]
- Gubbels Bupp, M.R.; Potluri, T.; Fink, A.L.; Klein, S.L. The Confluence of Sex Hormones and Aging on Immunity. Front. Immunol. 2018, 9, 1269. [Google Scholar] [CrossRef]
- Kayesh, M.E.H.; Yamato, O.; Rahman, M.M.; Hashem, M.A.; Maetani, F.; Eiei, T.; Mochizuki, K.; Sakurai, H.; Tsukiyama-Kohara, K. Molecular dynamics of koala retrovirus infection in captive koalas in Japan. Arch Virol. 2019, 164, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Olagoke, O.; Quigley, B.L.; Hemmatzadeh, F.; Tzipori, G.; Timms, P. Therapeutic vaccination of koalas harbouring endogenous koala retrovirus (KoRV) improves antibody responses and reduces circulating viral load. NPJ Vaccines 2020, 5, 60. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Regulation of adaptive immunity by the innate immune system. Science 2010, 327, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Young, G.R.; Eksmond, U.; Salcedo, R.; Alexopoulou, L.; Stoye, J.P.; Kassiotis, G. Resurrection of endogenous retroviruses in antibody-deficient mice. Nature 2012, 491, 774–778. [Google Scholar] [CrossRef]
- Yu, P.; Lübben, W.; Slomka, H.; Gebler, J.; Konert, M.; Cai, C.; Neubrandt, L.; Prazeres da Costa, O.; Paul, S.; Dehnert, S.; et al. Nucleic acid-sensing Toll-like receptors are essential for the control of endogenous retrovirus viremia and ERV-induced tumors. Immunity 2012, 37, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Pashine, A.; Valiante, N.M.; Ulmer, J.B. Targeting the innate immune response with improved vaccine adjuvants. Nat. Med. 2005, 11, S63–S68. [Google Scholar] [CrossRef]
- Olagoke, O.; Quigley, B.L.; Timms, P. Koalas vaccinated against Koala retrovirus respond by producing increased levels of interferon-gamma. Virol. J. 2020, 17, 168. [Google Scholar] [CrossRef] [PubMed]
- Colomer-Lluch, M.; Ruiz, A.; Moris, A.; Prado, J.G. Restriction factors: From intrinsic viral restriction to shaping cellular immunity against HIV-1. Front. Immunol. 2018, 9, 2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregorio, E.D.; Caproni, E.; Ulmer, J.B. Vaccine adjuvants: Mode of action. Front. Immunol. 2013, 4, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O’Carroll, D.; Zamore, P.D. PIWI-interacting RNAs: Small RNAs with big functions. Nat. Rev. Genet. 2019, 20, 89–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, Y.W.; Siomi, M.C.; Siomi, H. PIWI-Interacting RNA: Its Biogenesis and Functions. Annu. Rev. Biochem. 2015, 84, 405–433. [Google Scholar] [CrossRef] [PubMed]
- An, W.; Han, J.S.; Wheelan, S.J.; Davis, E.S.; Coombes, C.E.; Ye, P.; Triplett, C.; Boeke, J.D. Active retrotransposition by a synthetic L1 element in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 18662–18667. [Google Scholar] [CrossRef] [Green Version]
- Yeung, M.L.; Bennasser, Y.; Watashi, K.; Le, S.Y.; Houzet, L.; Jeang, K.T. Pyrosequencing of small non-coding RNAs in HIV-1 infected cells: Evidence for the processing of a viral-cellular double-stranded RNA hybrid. Nucleic Acids Res. 2009, 37, 6575–6586. [Google Scholar] [CrossRef]
- Yan, Z.; Hu, H.Y.; Jiang, X.; Maierhofer, V.; Neb, E.; He, L.; Hu, Y.; Hu, H.; Li, N.; Chen, W.; et al. Widespread expression of piRNA-like molecules in somatic tissues. Nucleic Acids Res. 2011, 39, 6596–6607. [Google Scholar] [CrossRef]
- Siomi, M.C.; Sato, K.; Pezic, D.; Aravin, A.A. PIWI-interacting small RNAs: The vanguard of genome defence. Nat. Rev. Mol. Cell Biol. 2011, 12, 246–258. [Google Scholar] [CrossRef]
- Ernst, C.; Odom, D.T.; Kutter, C. The emergence of piRNAs against transposon invasion to preserve mammalian genome integrity. Nat. Commun. 2017, 8, 1411. [Google Scholar] [CrossRef] [Green Version]
- Czech, B.; Hannon, G.J. One Loop to Rule Them All: The Ping-Pong Cycle and piRNA-Guided Silencing. Trends Biochem. Sci. 2016, 41, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Tarlinton, R.E.; Sarker, N.; Fabijan, J.; Dottorini, T.; Woolford, L.; Meers, J.; Simmons, G.; Owen, H.; Seddon, J.M.; Hemmatzedah, F.; et al. Differential and defective expression of Koala Retrovirus reveal complexity of host and virus evolution. bioRxiv 2017, 211466. [Google Scholar] [CrossRef] [Green Version]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., 3rd. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Güell, M.; Niu, D.; George, H.; Lesha, E.; Grishin, D.; Aach, J.; Shrock, E.; Xu, W.; Poci, J.; et al. Genome-wide inactivation of porcine endogenous retroviruses (PERVs). Science 2015, 350, 1101–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, D.; Wei, H.J.; Lin, L.; George, H.; Wang, T.; Lee, I.H.; Zhao, H.Y.; Wang, Y.; Kan, Y.; Shrock, E.; et al. Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9. Science 2017, 357, 1303–1307. [Google Scholar] [CrossRef] [Green Version]
- Yue, Y.; Xu, W.; Kan, Y.; Zhao, H.Y.; Zhou, Y.; Song, X.; Wu, J.; Xiong, J.; Goswami, D.; Yang, M.; et al. Extensive germline genome engineering in pigs. Nat. Biomed. Eng. 2021, 5, 134–143. [Google Scholar] [CrossRef]
- Tarlinton, R.E.; Fabijan, J.; Hemmatzadeh, F.; Meers, J.; Owen, H.; Sarker, N.; Seddon, J.M.; Simmons, G.; Speight, N.; Trott, D.J.; et al. Transcriptomic and genomic variants between koala populations reveals underlying genetic components to disorders in a bottlenecked population. Conserv. Genet. 2021. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kayesh, M.E.H.; Hashem, M.A.; Tsukiyama-Kohara, K. Toll-Like Receptor and Cytokine Responses to Infection with Endogenous and Exogenous Koala Retrovirus, and Vaccination as a Control Strategy. Curr. Issues Mol. Biol. 2021, 43, 52-64. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010005
Kayesh MEH, Hashem MA, Tsukiyama-Kohara K. Toll-Like Receptor and Cytokine Responses to Infection with Endogenous and Exogenous Koala Retrovirus, and Vaccination as a Control Strategy. Current Issues in Molecular Biology. 2021; 43(1):52-64. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010005
Chicago/Turabian StyleKayesh, Mohammad Enamul Hoque, Md Abul Hashem, and Kyoko Tsukiyama-Kohara. 2021. "Toll-Like Receptor and Cytokine Responses to Infection with Endogenous and Exogenous Koala Retrovirus, and Vaccination as a Control Strategy" Current Issues in Molecular Biology 43, no. 1: 52-64. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010005