Vitamin D Deficiency Reduces Vascular Reactivity of Coronary Arterioles in Male Rats

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Treatment and Experimental Protocol
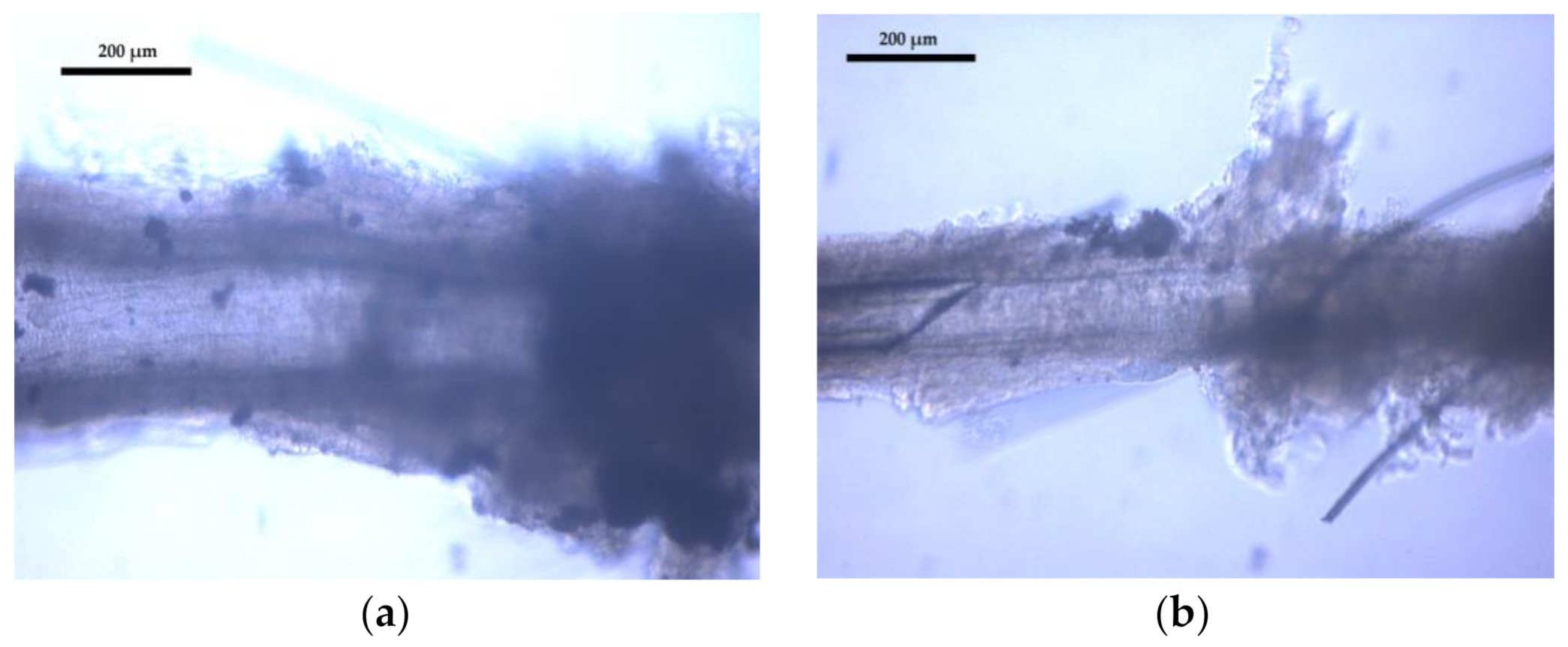
2.3. Pressure Microarteriography of Coronary Arterioles
2.4. Calculations
2.5. Immunohistochemistry of Coronary Arterioles
2.6. Statistical Analysis
3. Results
3.1. Heart Weight
3.2. Coronary Arteriole Morphology and Function
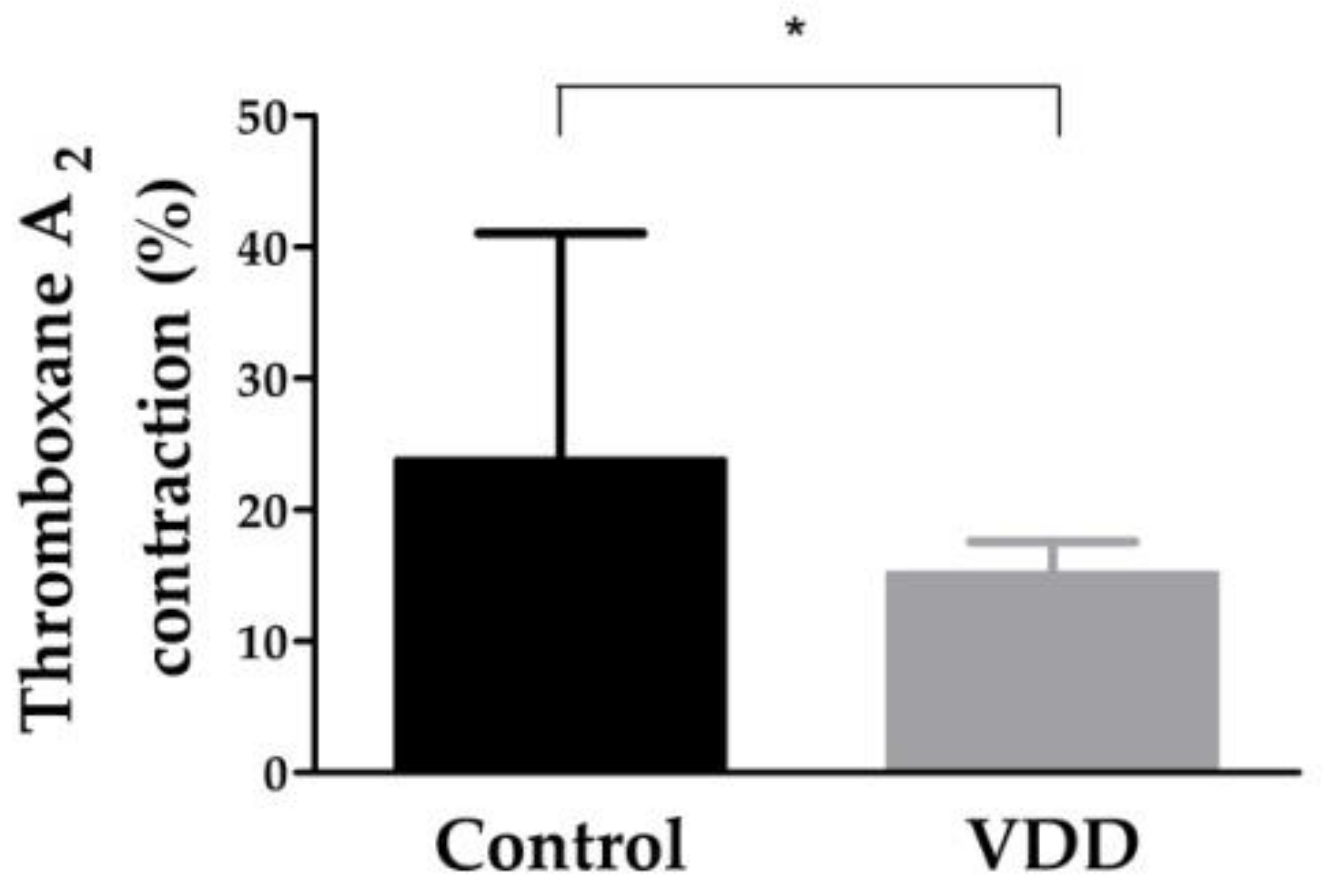
3.3. Constriction Capacity of Coronary Arterioles: Thromboxane-A2-Induced Contraction
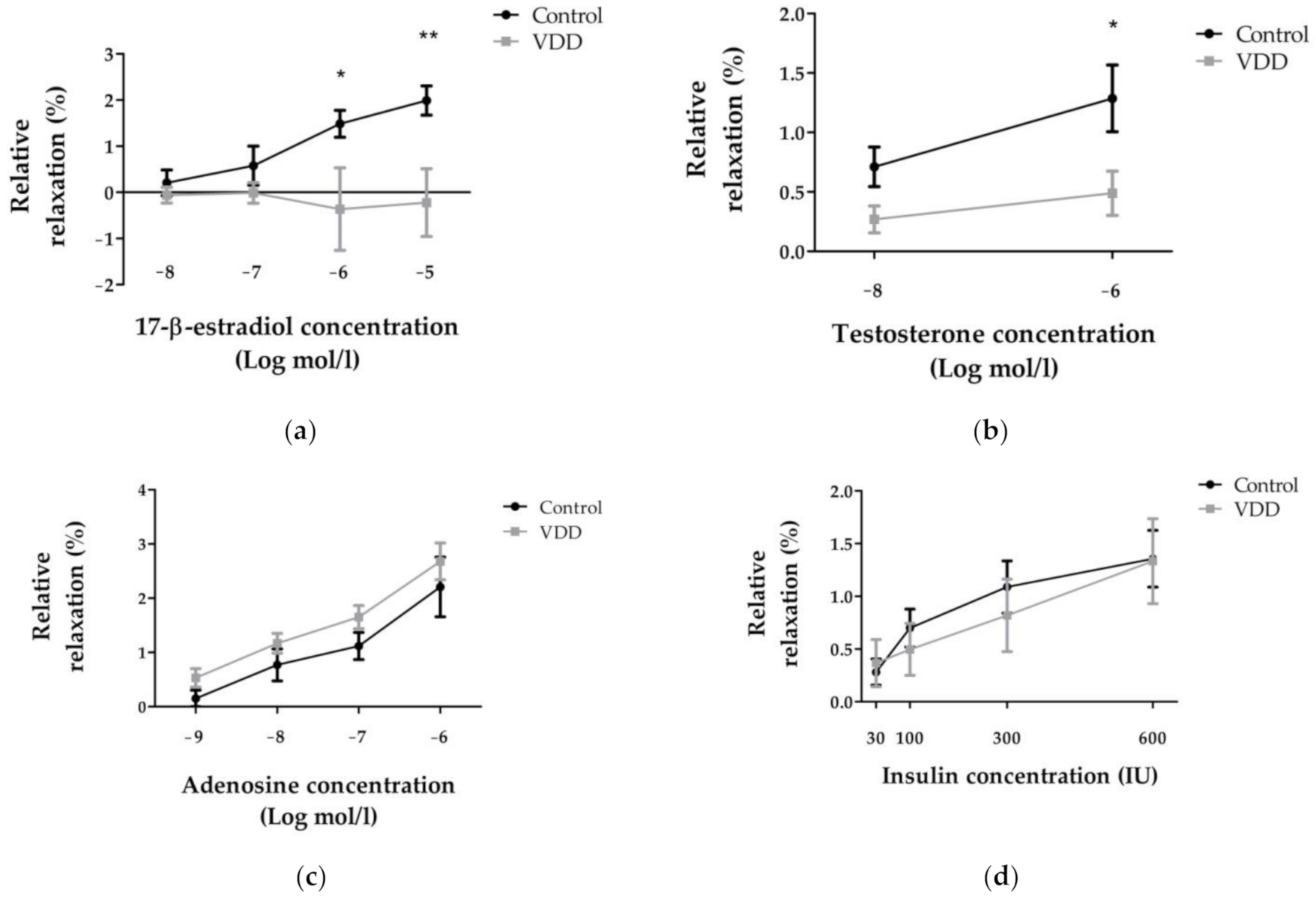
3.4. Relaxation Ability of Coronary Arterioles: 17-β-Estradiol-, Testosterone-, Adenosine-, and Insulin-Induced Relaxation
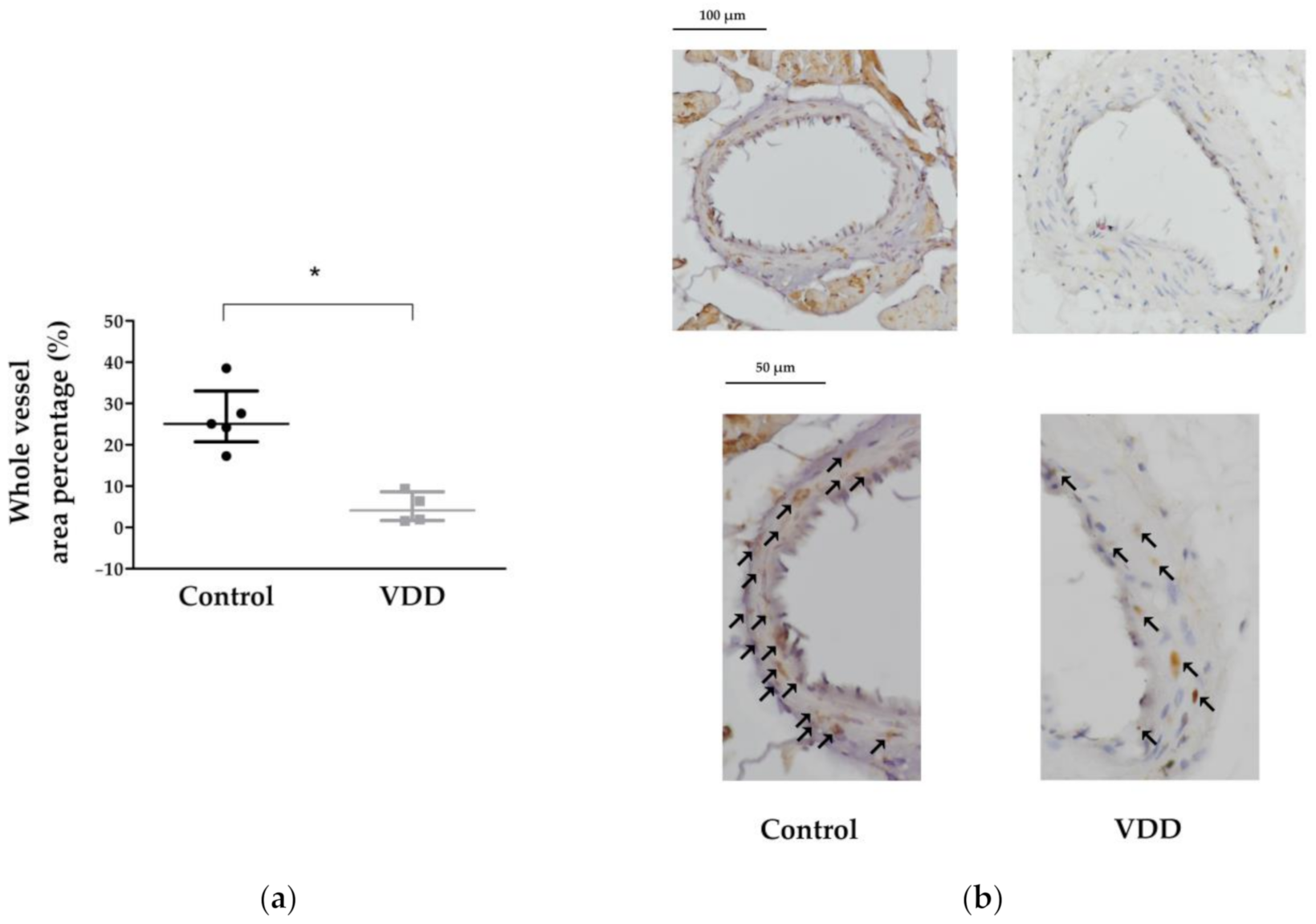
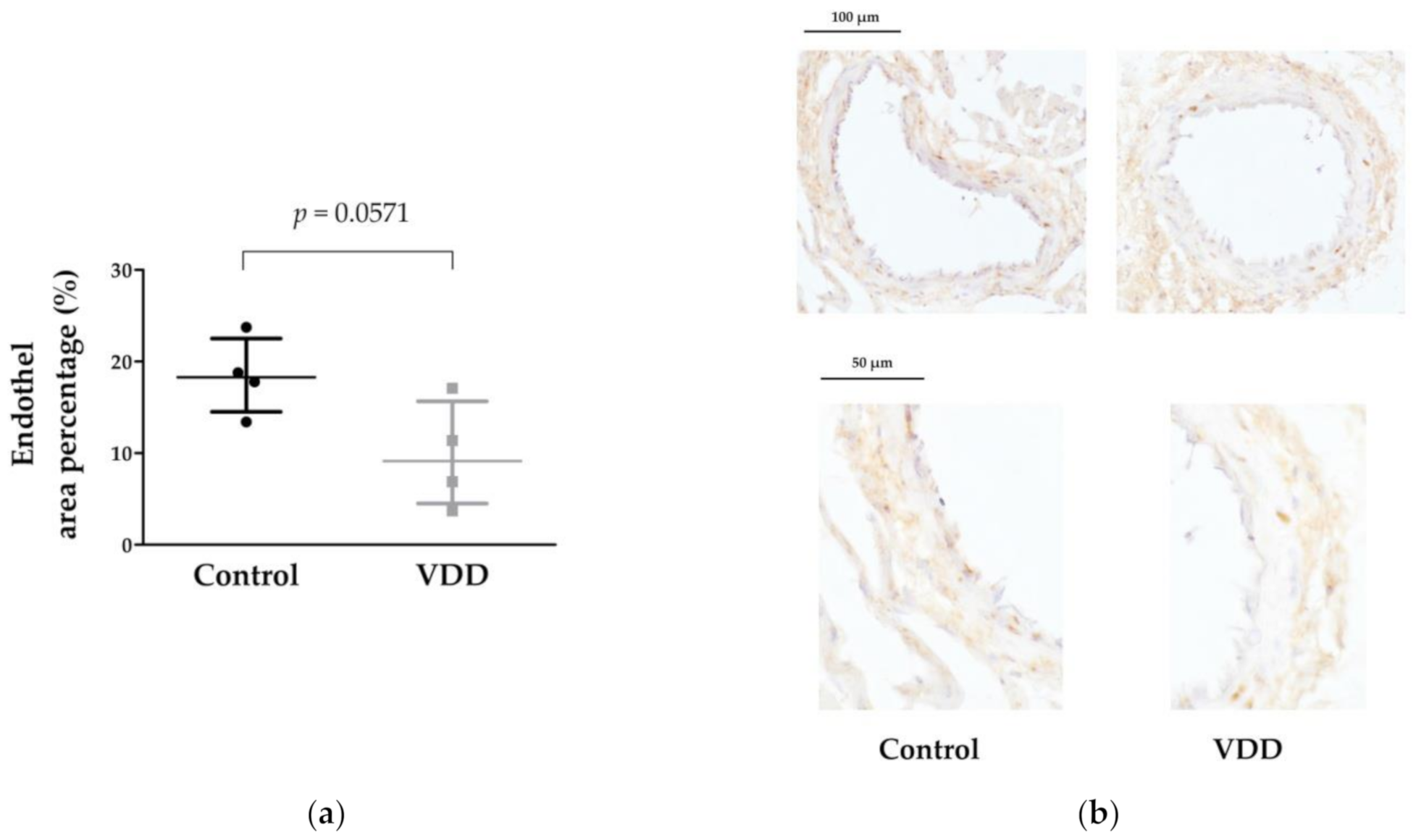
3.5. Immunohistochemical Stainings: TP, ERα, AR, eNOS, and VDR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bouillon, R. Vitamin D and cardiovascular disorders. Osteoporos. Int. 2019, 30, 2167–2181. [Google Scholar] [CrossRef] [PubMed]
- Kienreich, K.; Tomaschitz, A.; Verheyen, N.; Pieber, T.; Gaksch, M.; Grubler, M.R.; Pilz, S. Vitamin D and cardiovascular disease. Nutrients 2013, 5, 3005–3021. [Google Scholar] [CrossRef] [PubMed]
- Gholami, F.; Moradi, G.; Zareei, B.; Rasouli, M.A.; Nikkhoo, B.; Roshani, D.; Ghaderi, E. The association between circulating 25-hydroxyvitamin D and cardiovascular diseases: A meta-analysis of prospective cohort studies. BMC Cardiovasc. Disord. 2019, 19, 248. [Google Scholar] [CrossRef] [PubMed]
- Pludowski, P.; Karczmarewicz, E.; Bayer, M.; Carter, G.; Chlebna-Sokol, D.; Czech-Kowalska, J.; Debski, R.; Decsi, T.; Dobrzanska, A.; Franek, E.; et al. Practical guidelines for the supplementation of vitamin D and the treatment of deficits in Central Europe–recommended vitamin D intakes in the general population and groups at risk of vitamin D deficiency. Endokrynol. Pol. 2013, 64, 319–327. [Google Scholar] [CrossRef]
- Giustina, A.; Bouillon, R.; Binkley, N.; Sempos, C.; Adler, R.A.; Bollerslev, J.; Dawson-Hughes, B.; Ebeling, P.R.; Feldman, D.; Heijboer, A.; et al. Controversies in Vitamin D: A Statement From the Third International Conference. JBMR Plus 2020, 4, e10417. [Google Scholar] [CrossRef]
- Hii, C.S.; Ferrante, A. The Non-Genomic Actions of Vitamin D. Nutrients 2016, 8, 135. [Google Scholar] [CrossRef] [Green Version]
- Pike, J.W.; Christakos, S. Biology and Mechanisms of Action of the Vitamin D Hormone. Endocrinol. Metab. Clin. N. Am. 2017, 46, 815–843. [Google Scholar] [CrossRef]
- Mozos, I.; Marginean, O. Links between Vitamin D Deficiency and Cardiovascular Diseases. Biomed. Res. Int. 2015, 2015, 109275. [Google Scholar] [CrossRef]
- Dawber, T.R.; Kannel, W.B.; Revotskie, N.; Stokes, J., 3rd; Kagan, A.; Gordon, T. Some factors associated with the development of coronary heart disease: Six years’ follow-up experience in the Framingham study. Am. J. Public Health Nations Health 1959, 49, 1349–1356. [Google Scholar] [CrossRef]
- Dawber, T.R.; Moore, F.E.; Mann, G.V. Coronary heart disease in the Framingham study. Am. J. Public Health Nations Health 1957, 47, 4–24. [Google Scholar] [CrossRef]
- Kannel, W.B.; Dawber, T.R.; Kagan, A.; Revotskie, N.; Stokes, J., 3rd. Factors of risk in the development of coronary heart disease—Six year follow-up experience. The Framingham Study. Ann. Intern. Med. 1961, 55, 33–50. [Google Scholar] [CrossRef]
- Wong, N.D.; Levy, D. Legacy of the framingham heart study: Rationale, design, initial findings, and implications. Glob. Heart 2013, 8, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Madan, N.; Gajo, E.; Sanghani, R.M.; Volgman, A.S. Sex-Based Considerations in the Evaluation of Chest Pain and Management of Obstructive Coronary Artery Disease. Curr. Atheroscler. Rep. 2020, 22, 39. [Google Scholar] [CrossRef]
- Kristensen, S.D.; Husted, S.E.; Nielsen, H.K.; Ravn, H.B.; Vissinger, H. Interaction between thrombocytes and blood vessel wall—Significance for acute ischemic coronary syndromes. Ugeskr. Laeger 1995, 157, 2295–2298. [Google Scholar]
- Ellinsworth, D.C.; Shukla, N.; Fleming, I.; Jeremy, J.Y. Interactions between thromboxane A(2), thromboxane/prostaglandin (TP) receptors, and endothelium-derived hyperpolarization. Cardiovasc. Res. 2014, 102, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Braune, S.; Kupper, J.H.; Jung, F. Effect of Prostanoids on Human Platelet Function: An Overview. Int. J. Mol. Sci. 2020, 21, 9020. [Google Scholar] [CrossRef]
- Aryan, L.; Younessi, D.; Zargari, M.; Banerjee, S.; Agopian, J.; Rahman, S.; Borna, R.; Ruffenach, G.; Umar, S.; Eghbali, M. The Role of Estrogen Receptors in Cardiovascular Disease. Int. J. Mol. Sci. 2020, 21, 4314. [Google Scholar] [CrossRef]
- Lorigo, M.; Mariana, M.; Oliveira, N.; Lemos, M.C.; Cairrao, E. Vascular Pathways of Testosterone: Clinical Implications. J. Cardiovasc. Transl. Res. 2020, 13, 55–72. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, N.; Silveyra, P. Estrogen receptor signaling mechanisms. Adv. Protein Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar] [CrossRef]
- Guivarc’h, E.; Buscato, M.; Guihot, A.L.; Favre, J.; Vessieres, E.; Grimaud, L.; Wakim, J.; Melhem, N.J.; Zahreddine, R.; Adlanmerini, M.; et al. Predominant Role of Nuclear Versus Membrane Estrogen Receptor alpha in Arterial Protection: Implications for Estrogen Receptor alpha Modulation in Cardiovascular Prevention/Safety. J. Am. Heart Assoc. 2018, 7, e008950. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Herald, A.K.; Alves-Lopes, R.; Montezano, A.C.; Ahmed, S.F.; Touyz, R.M. Genomic and non-genomic effects of androgens in the cardiovascular system: Clinical implications. Clin. Sci. 2017, 131, 1405–1418. [Google Scholar] [CrossRef] [Green Version]
- Salzano, A.; Demelo-Rodriguez, P.; Marra, A.M.; Proietti, M. A Focused Review of Gender Differences in Antithrombotic Therapy. Curr. Med. Chem. 2017, 24, 2576–2588. [Google Scholar] [CrossRef]
- Pal, E.; Hadjadj, L.; Fontanyi, Z.; Monori-Kiss, A.; Mezei, Z.; Lippai, N.; Magyar, A.; Heinzlmann, A.; Karvaly, G.; Monos, E.; et al. Vitamin D deficiency causes inward hypertrophic remodeling and alters vascular reactivity of rat cerebral arterioles. PLoS ONE 2018, 13, e0192480. [Google Scholar] [CrossRef]
- Nadasy, G.L.; Szekeres, M.; Dezsi, L.; Varbiro, S.; Szekacs, B.; Monos, E. Preparation of intramural small coronary artery and arteriole segments and resistance artery networks from the rat heart for microarteriography and for in situ perfusion video mapping. Microvasc. Res. 2001, 61, 282–286. [Google Scholar] [CrossRef]
- Sziva, R.E.; Fontanyi, Z.; Pal, E.; Hadjadj, L.; Monori-Kiss, A.; Horvath, E.M.; Benko, R.; Magyar, A.; Heinzlmann, A.; Benyo, Z.; et al. Vitamin D Deficiency Induces Elevated Oxidative and Biomechanical Damage in Coronary Arterioles in Male Rats. Antioxidants 2020, 9, 997. [Google Scholar] [CrossRef]
- Varbiro, S.; Matrai, M.; Szekeres, M.; Nadasy, G.L.; Szaky, E.; Mericli, M.; Banhidy, F.; Monos, E.; Szekacs, B. Intramural coronary artery constrictor reactivity to thromboxane is higher in male than in female rats. Gynecol. Endocrinol. 2006, 22, 44–47. [Google Scholar] [CrossRef]
- Moreno, J.; Krishnan, A.V.; Swami, S.; Nonn, L.; Peehl, D.M.; Feldman, D. Regulation of prostaglandin metabolism by calcitriol attenuates growth stimulation in prostate cancer cells. Cancer Res. 2005, 65, 7917–7925. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.; Krishnan, A.V.; Peehl, D.M.; Feldman, D. Mechanisms of vitamin D-mediated growth inhibition in prostate cancer cells: Inhibition of the prostaglandin pathway. Anticancer Res. 2006, 26, 2525–2530. [Google Scholar]
- Feldman, D.; Krishnan, A.; Moreno, J.; Swami, S.; Peehl, D.M.; Srinivas, S. Vitamin D inhibition of the prostaglandin pathway as therapy for prostate cancer. Nutr. Rev. 2007, 65, S113–S115. [Google Scholar] [CrossRef]
- Horiuchi, H.; Nagata, I.; Komoriya, K. Protective effect of vitamin D3 analogues on endotoxin shock in mice. Agents Actions 1991, 33, 343–348. [Google Scholar] [CrossRef]
- Wong, M.S.; Delansorne, R.; Man, R.Y.; Svenningsen, P.; Vanhoutte, P.M. Chronic treatment with vitamin D lowers arterial blood pressure and reduces endothelium-dependent contractions in the aorta of the spontaneously hypertensive rat. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1226–H1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, M.S.; Delansorne, R.; Man, R.Y.; Vanhoutte, P.M. Vitamin D derivatives acutely reduce endothelium-dependent contractions in the aorta of the spontaneously hypertensive rat. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H289–H296. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Liu, Y.; Chu, L.M.; Clements, R.T.; Khabbaz, K.R.; Robich, M.P.; Bianchi, C.; Sellke, F.W. Thromboxane-induced contractile response of human coronary arterioles is diminished after cardioplegic arrest. Ann. Thorac. Surg. 2011, 92, 829–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandi, A.A.; Wadhwani, N.S.; Joshi, S.R. Maternal vitamin D deficiency increases the thromboxane/prostacyclin ratio through alterations in the one-carbon cycle in Wistar rats. Biofactors 2019, 45, 548–555. [Google Scholar] [CrossRef]
- Nandi, A.; Wadhwani, N.; Joshi, S.R. Vitamin D deficiency influences fatty acid metabolism. Prostaglandins Leukot. Essent. Fatty Acids 2019, 140, 57–63. [Google Scholar] [CrossRef]
- Hadjadj, L.; Monori-Kiss, A.; Horvath, E.M.; Heinzlmann, A.; Magyar, A.; Sziva, R.E.; Miklos, Z.; Pal, E.; Gal, J.; Szabo, I.; et al. Geometric, elastic and contractile-relaxation changes in coronary arterioles induced by Vitamin D deficiency in normal and hyperandrogenic female rats. Microvasc. Res. 2019, 122, 78–84. [Google Scholar] [CrossRef]
- Pugh, P.J.; Jones, R.D.; Jones, T.H.; Channer, K.S. Intrinsic responses of rat coronary arteries in vitro: Influence of testosterone, calcium, and effective transmural pressure. Endocrine 2002, 19, 155–161. [Google Scholar] [CrossRef]
- Pal, E.; Hadjadj, L.; Fontanyi, Z.; Monori-Kiss, A.; Lippai, N.; Horvath, E.M.; Magyar, A.; Horvath, E.; Monos, E.; Nadasy, G.L.; et al. Gender, hyperandrogenism and vitamin D deficiency related functional and morphological alterations of rat cerebral arteries. PLoS ONE 2019, 14, e0216951. [Google Scholar] [CrossRef]
- Santos, R.L.; Abreu, G.R.; Bissoli, N.S.; Moyses, M.R. Endothelial mediators of 17 beta-estradiol-induced coronary vasodilation in the isolated rat heart. Braz. J. Med. Biol. Res. 2004, 37, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.L.; Marin, E.B.; Goncalves, W.L.; Bissoli, N.S.; Abreu, G.R.; Moyses, M.R. Sex differences in the coronary vasodilation induced by 17 beta-oestradiol in the isolated perfused heart from spontaneously hypertensive rats. Acta Physiol. 2010, 200, 203–210. [Google Scholar] [CrossRef]
- Santos, R.L.; Lima, J.T.; Rouver, W.N.; Moyses, M.R. Deficiency of sex hormones does not affect 17-ss-estradiol-induced coronary vasodilation in the isolated rat heart. Braz. J. Med. Biol. Res. 2016, 49, e5058. [Google Scholar] [CrossRef] [Green Version]
- Levy, A.S.; Chung, J.C.; Kroetsch, J.T.; Rush, J.W. Nitric oxide and coronary vascular endothelium adaptations in hypertension. Vasc. Health Risk Manag. 2009, 5, 1075–1087. [Google Scholar] [CrossRef] [Green Version]
- Hadjadj, L.; Varbiro, S.; Horvath, E.M.; Monori-Kiss, A.; Pal, E.; Karvaly, G.B.; Heinzlmann, A.; Magyar, A.; Szabo, I.; Sziva, R.E.; et al. Insulin resistance in an animal model of polycystic ovary disease is aggravated by vitamin D deficiency: Vascular consequences. Diabetes Vasc. Dis. Res. 2018, 15, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Lajtai, K.; Tarszabó, R.; Bányai, B.; Péterffy, B.; Gerszi, D.; Ruisanchez, É.; Sziva, R.E.; Korsós-Novák, Á.; Benkő, R.; Hadjadj, L.; et al. Effect of Vitamin D Status on Vascular Function of the Aorta in a Rat Model of PCOS. Oxid. Med. Cell. Longev. 2021, 2021, 8865979. [Google Scholar] [CrossRef]
- Bouillon, R.; Marcocci, C.; Carmeliet, G.; Bikle, D.; White, J.H.; Dawson-Hughes, B.; Lips, P.; Munns, C.F.; Lazaretti-Castro, M.; Giustina, A.; et al. Skeletal and Extraskeletal Actions of Vitamin D: Current Evidence and Outstanding Questions. Endocr. Rev. 2019, 40, 1109–1151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; McCullough, P.A.; Tecson, K.M. Vitamin D deficiency in association with endothelial dysfunction: Implications for patients with COVID-19. Rev. Cardiovasc. Med. 2020, 21, 339–344. [Google Scholar] [CrossRef]
- de la Guia-Galipienso, F.; Martinez-Ferran, M.; Vallecillo, N.; Lavie, C.J.; Sanchis-Gomar, F.; Pareja-Galeano, H. Vitamin D and cardiovascular health. Clin. Nutr. 2020. [Google Scholar] [CrossRef]
- Maggio, M.; De Vita, F.; Lauretani, F.; Ceda, G.P.; Volpi, E.; Giallauria, F.; De Cicco, G.; Cattabiani, C.; Melhus, H.; Michaelsson, K.; et al. Vitamin D and endothelial vasodilation in older individuals: Data from the PIVUS study. J. Clin. Endocrinol. Metab. 2014, 99, 3382–3389. [Google Scholar] [CrossRef] [Green Version]
- Dusso, A.S.; Brown, A.J.; Slatopolsky, E. Vitamin D. Am. J. Physiol. Ren. Physiol. 2005, 289, F8–F28. [Google Scholar] [CrossRef]
- Bischoff-Ferrari, H.A.; Vellas, B.; Rizzoli, R.; Kressig, R.W.; da Silva, J.A.P.; Blauth, M.; Felson, D.T.; McCloskey, E.V.; Watzl, B.; Hofbauer, L.C.; et al. Effect of Vitamin D Supplementation, Omega-3 Fatty Acid Supplementation, or a Strength-Training Exercise Program on Clinical Outcomes in Older Adults: The DO-HEALTH Randomized Clinical Trial. JAMA 2020, 324, 1855–1868. [Google Scholar] [CrossRef]
- Tice, J.A.; Halalau, A.; Burke, H. Vitamin D Does Not Prevent Cancer or Cardiovascular Disease: The VITAL Trial: Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Gordon, D.; Copeland, T.; D’Agostino, D.; et al. Vitamin D Supplements and Prevention of Cancer and Cardiovascular Disease. N. Engl. J. Med. 2019, 380, 33–44. [Google Scholar] [CrossRef]
- Scragg, R. The Vitamin D Assessment (ViDA) study—Design and main findings. J. Steroid Biochem. Mol. Biol. 2020, 198, 105562. [Google Scholar] [CrossRef]
- Brondum-Jacobsen, P.; Benn, M.; Jensen, G.B.; Nordestgaard, B.G. 25-hydroxyvitamin d levels and risk of ischemic heart disease, myocardial infarction, and early death: Population-based study and meta-analyses of 18 and 17 studies. Arter. Thromb. Vasc. Biol. 2012, 32, 2794–2802. [Google Scholar] [CrossRef] [Green Version]
- Karohl, C.; Vaccarino, V.; Veledar, E.; Goldberg, J.; Tangpricha, V.; Bellasi, A.; Raggi, P. Vitamin D status and coronary flow reserve measured by positron emission tomography: A co-twin control study. J. Clin. Endocrinol. Metab. 2013, 98, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Lerchbaum, E.; Pilz, S.; Boehm, B.O.; Grammer, T.B.; Obermayer-Pietsch, B.; Marz, W. Combination of low free testosterone and low vitamin D predicts mortality in older men referred for coronary angiography. Clin. Endocrinol. 2012, 77, 475–483. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters/Groups | Control | VDD |
|---|---|---|
| Inner radii (μm) 1: | 89.06 ± 5.23 | 61.17 ± 5.81 ** |
| Outer radii (μm) 1: | 131.6 ± 8.49 | 108.3 ± 7.55 |
| Myogenic tone (%) 2: | 5.14 [1.30–18.41] | 2.44 [1.36–4.18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontányi, Z.; Sziva, R.E.; Pál, É.; Hadjadj, L.; Monori-Kiss, A.; Horváth, E.M.; Benkő, R.; Magyar, A.; Heinzlmann, A.; Benyó, Z.; et al. Vitamin D Deficiency Reduces Vascular Reactivity of Coronary Arterioles in Male Rats. Curr. Issues Mol. Biol. 2021, 43, 79-92. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010007
Fontányi Z, Sziva RE, Pál É, Hadjadj L, Monori-Kiss A, Horváth EM, Benkő R, Magyar A, Heinzlmann A, Benyó Z, et al. Vitamin D Deficiency Reduces Vascular Reactivity of Coronary Arterioles in Male Rats. Current Issues in Molecular Biology. 2021; 43(1):79-92. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010007
Chicago/Turabian StyleFontányi, Zoltán, Réka Eszter Sziva, Éva Pál, Leila Hadjadj, Anna Monori-Kiss, Eszter Mária Horváth, Rita Benkő, Attila Magyar, Andrea Heinzlmann, Zoltán Benyó, and et al. 2021. "Vitamin D Deficiency Reduces Vascular Reactivity of Coronary Arterioles in Male Rats" Current Issues in Molecular Biology 43, no. 1: 79-92. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010007