
Active Ingredients from Euodia ruticarpa Steam Distilled Essential Oil Inhibit PC-3 Prostate Cancer Cell Growth via Direct Action and Indirect Immune Cells Conditioned Media In Vitro


Abstract
:1. Background
2. Material and Methods
2.1. Preparation and Purification of Euodia ruticarpa (ER) Steam Distilled Essential Oil (SDEO)
2.2. Characterization of Active Ingredients in ER SDEO and Its Isolated Fractions
2.3. Isolation of Mouse Primary Immune Cells
2.4. Effects of Treatments with ER SDEO and Its Isolated Fractions ERF1–F6 on Th1/Th2 Cytokines Secreted by Mouse Primary Splenocytes
2.5. Effects of Treatments with ER SDEO and Its Isolated Fractions ERF1–F3 on Pro-/Anti-Inflammatory Cytokines Secreted by Mouse Peritoneal Macrophages in the Absence or Presence of LPS
2.6. Th1/Th2 and Pro-/Anti-Inflammatory Cytokine Assays with an Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Preparation of Splenocyte-Conditioned Media (SCM) and Macrophage-Conditioned Media (MCM) with ER SDEO and Its Isolated Fractions ERF1–3
2.8. Culture of Human Prostate Cancer PC-3 Cells
2.9. Direct Treatment Effects of ER SDEO and Its Isolated Fractions ERF1–3 on the Growth of Human Prostate Cancer PC-3 Cells
2.10. Effects of Treatments with SCM or MCM Cultured with ER SDEO and Its Isolated Fractions ERF1–F3 on Human Prostate Cancer PC-3 Cell Growth
2.11. Statistical Analysis
3. Results
3.1. Characterization of Active Ingredients in ER SDEO and Its Isolated Fractions
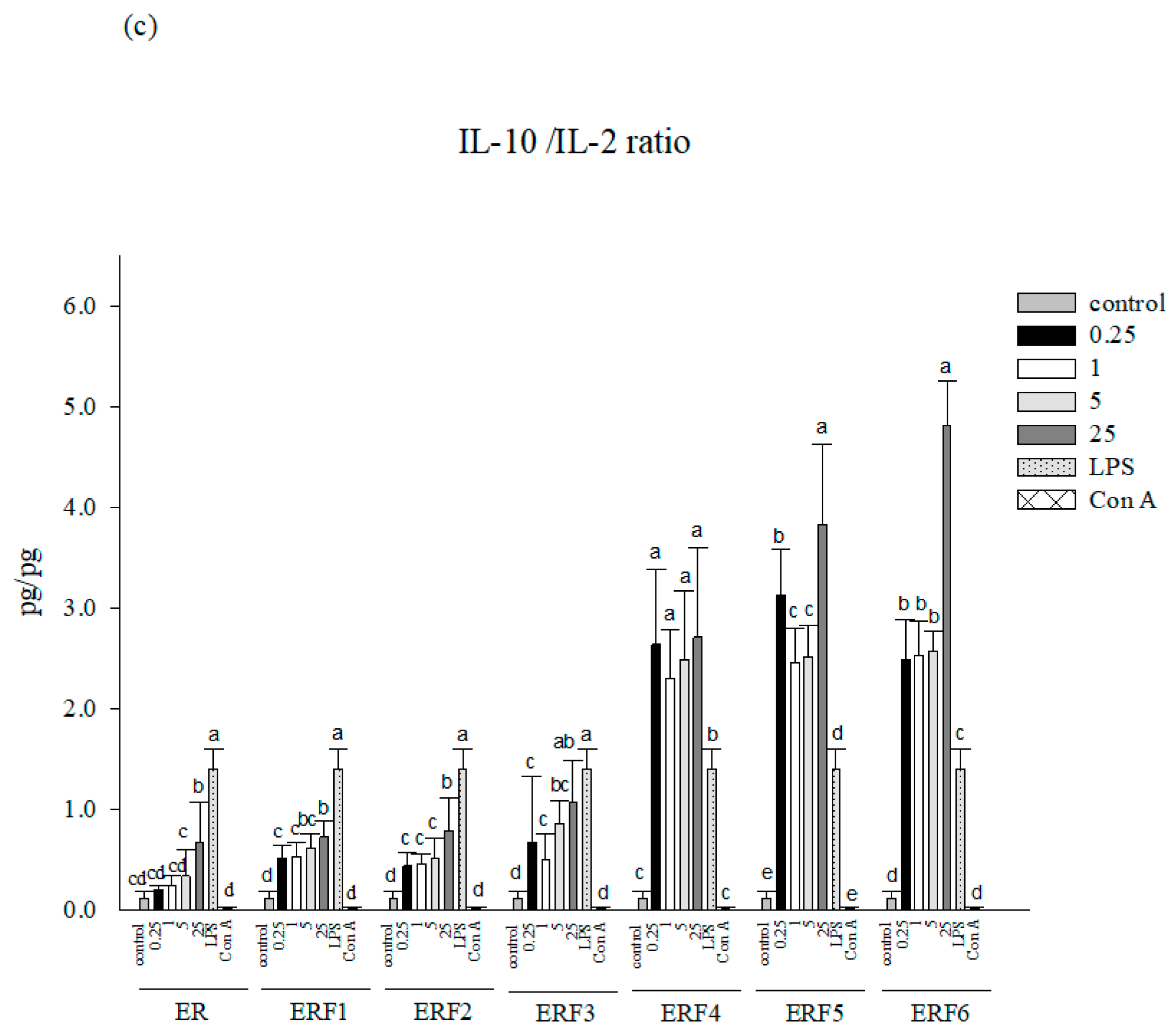
3.2. ER SDEO and ERF1–F6 Effects on Th1/Th2 Cytokine Secretions Using Mouse Primary Splenocytes
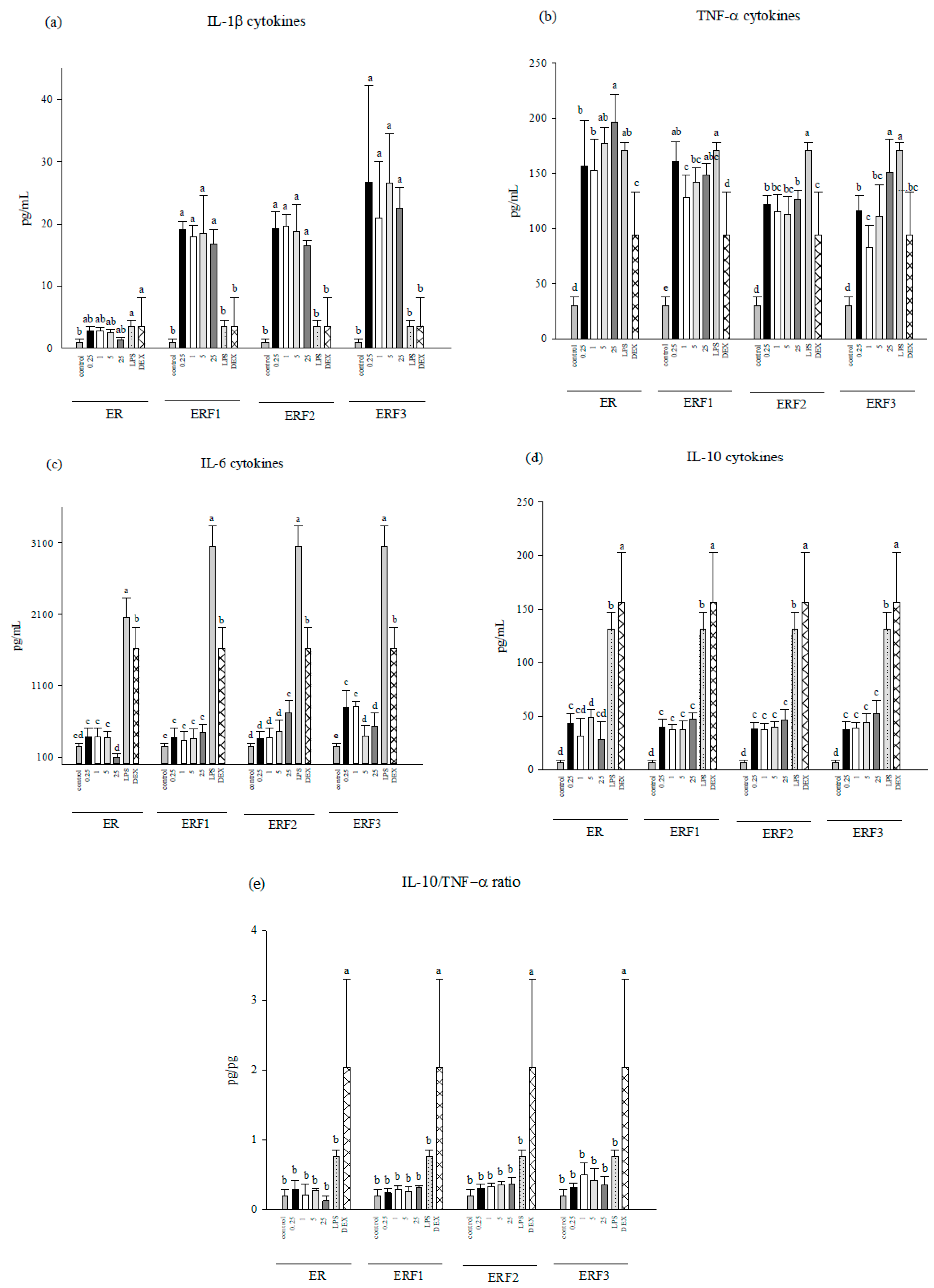
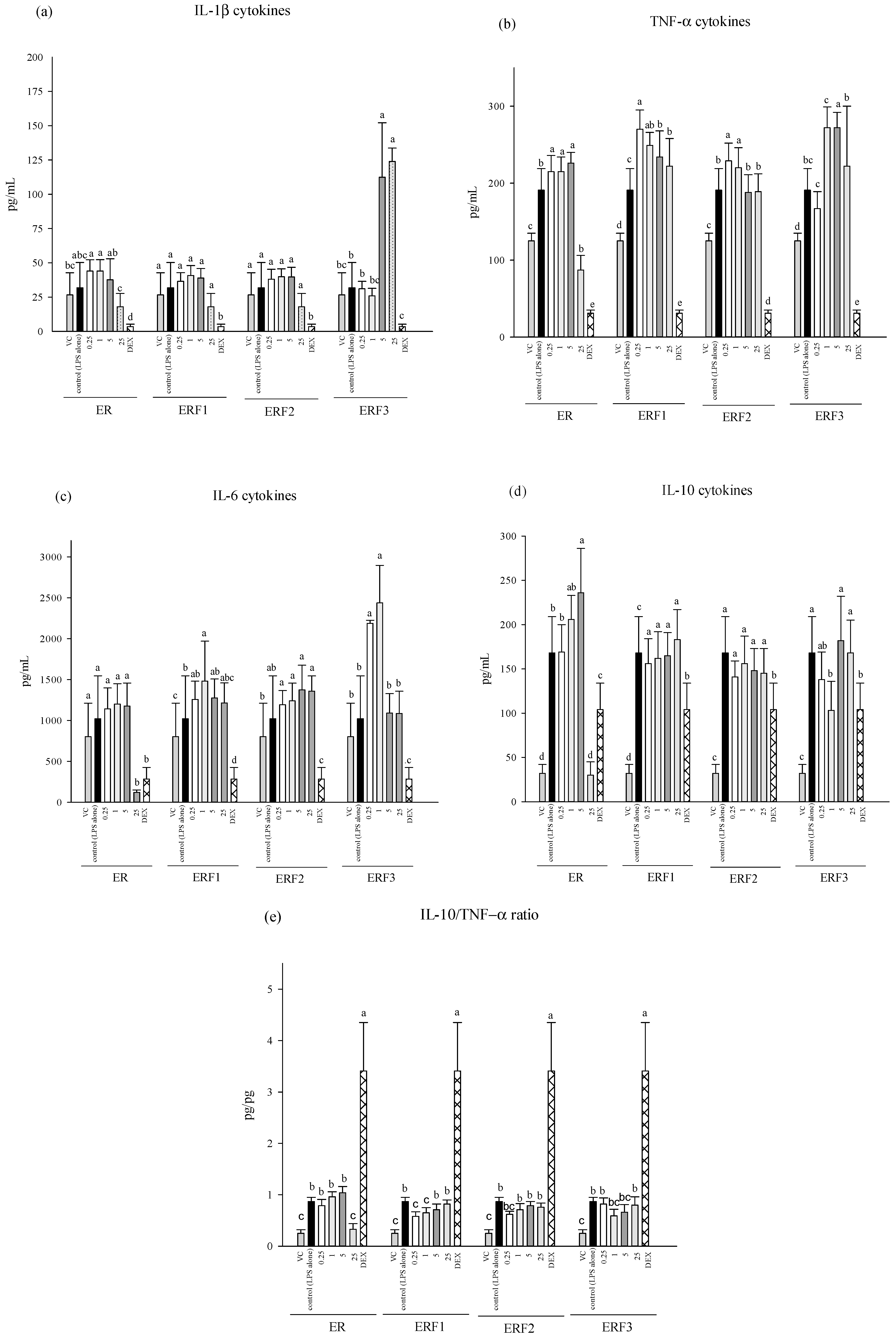
3.3. ER SDEO and ERF1–F3 Effects on Pro-/Anti-Inflammatory Cytokine Secretions Using Mouse Peritoneal Macrophages in the Absence or Presence of LPS
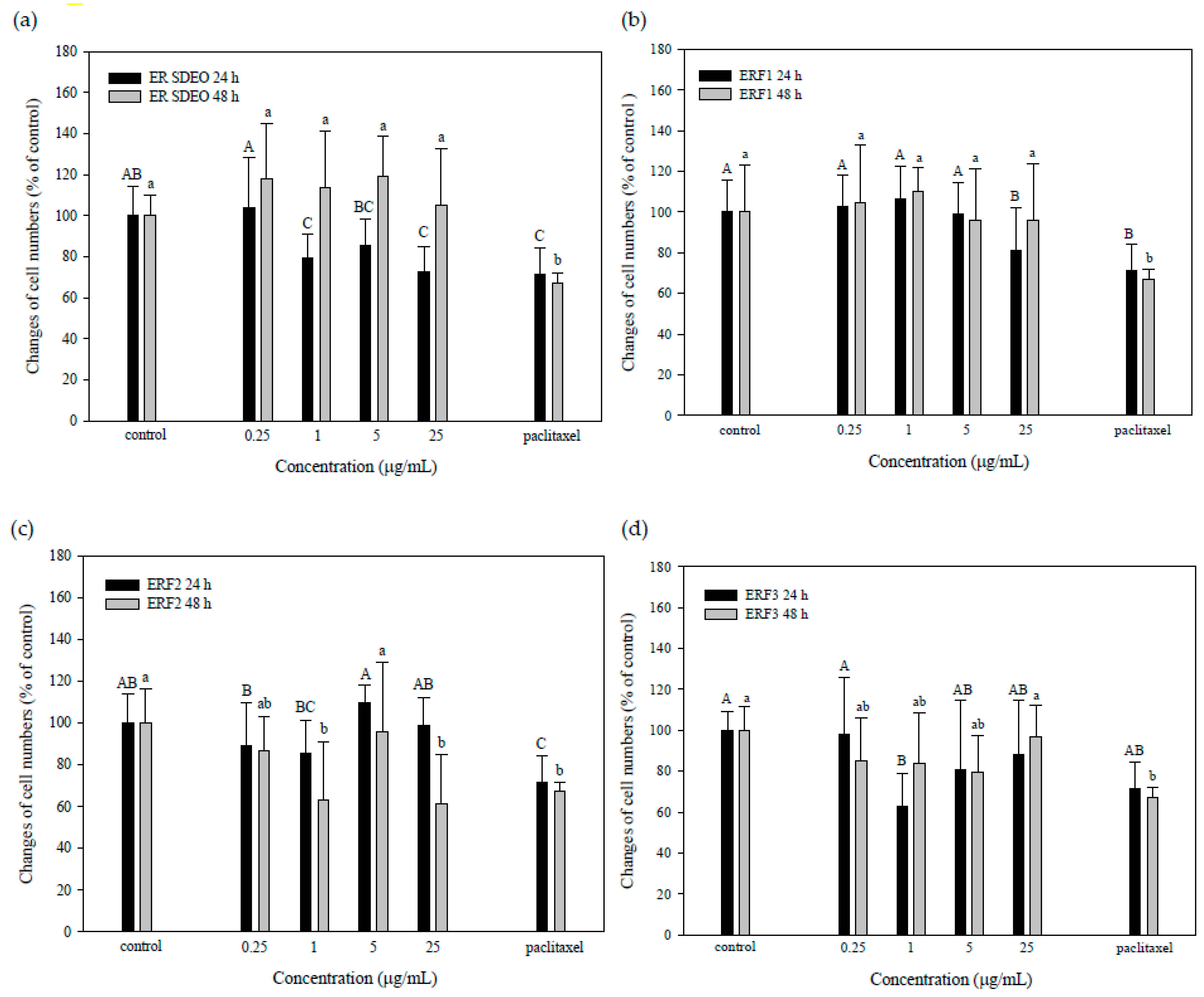
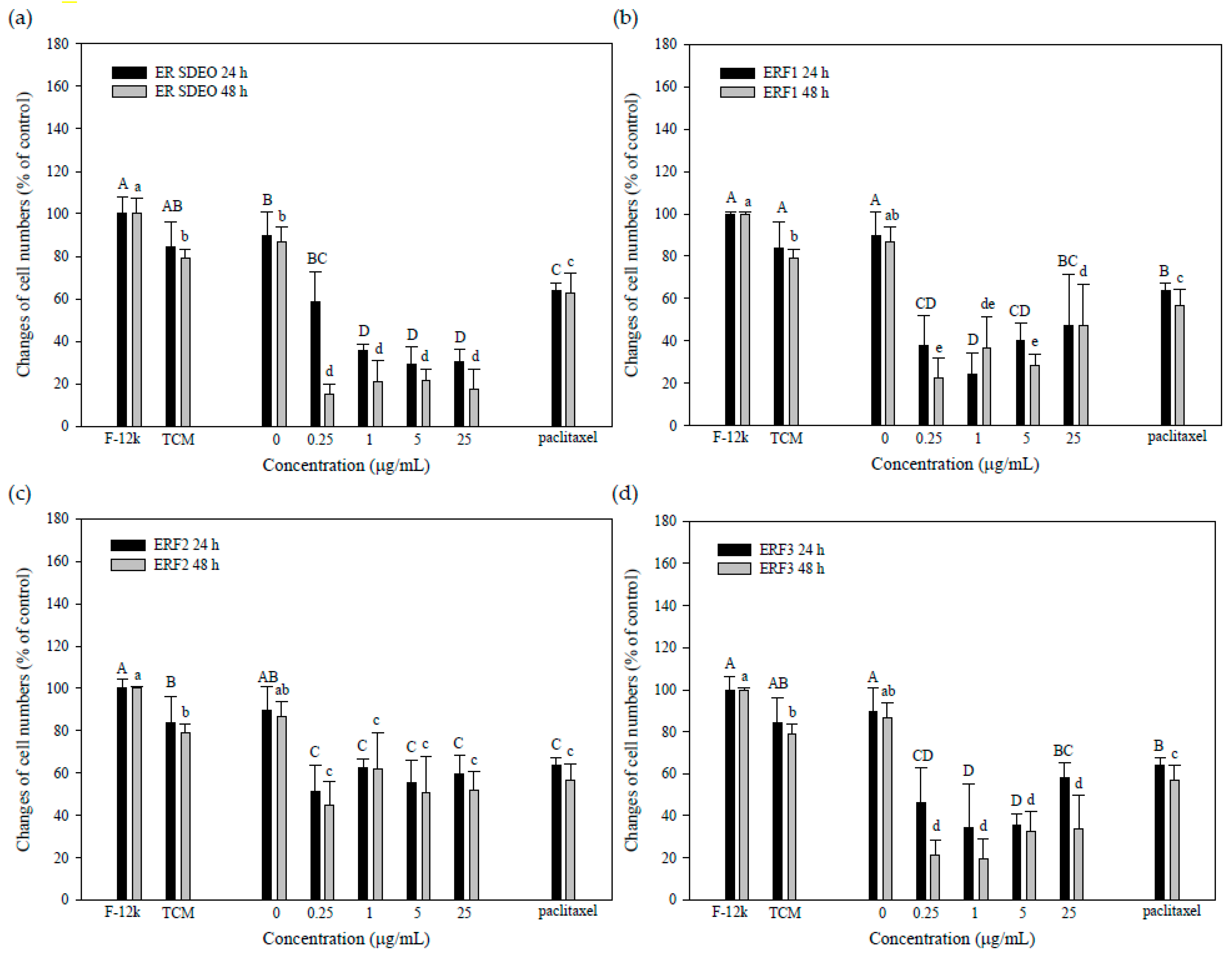
3.4. ER SDEO and ERF1–F3 Direct Action against PC-3 Cell Growth
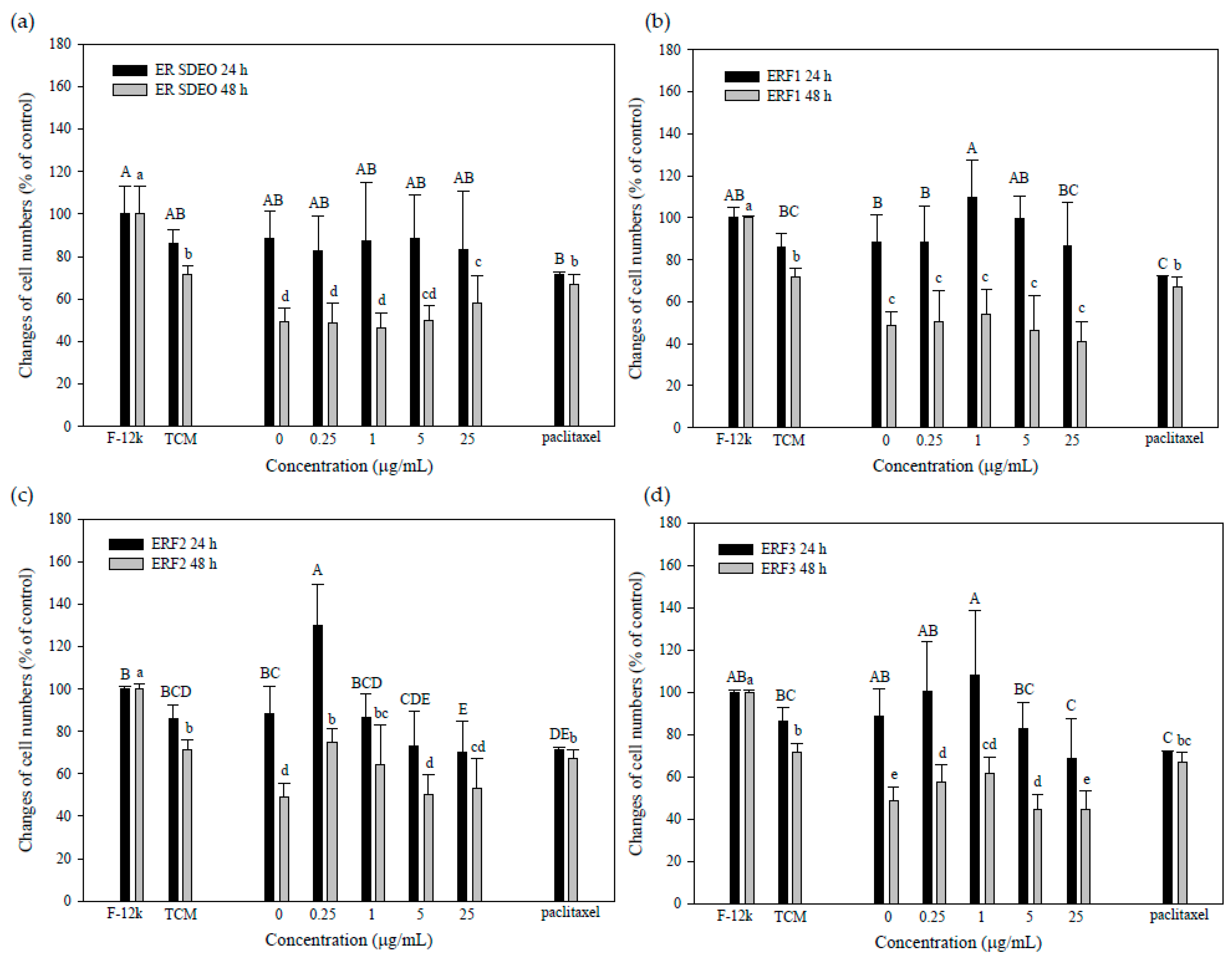
3.5. Indirect Treatment against PC-3 Cell Growth by SCM or MCM in the Absence or Presence of ER SDEO and Its Isolated Fractions ERF1–F3
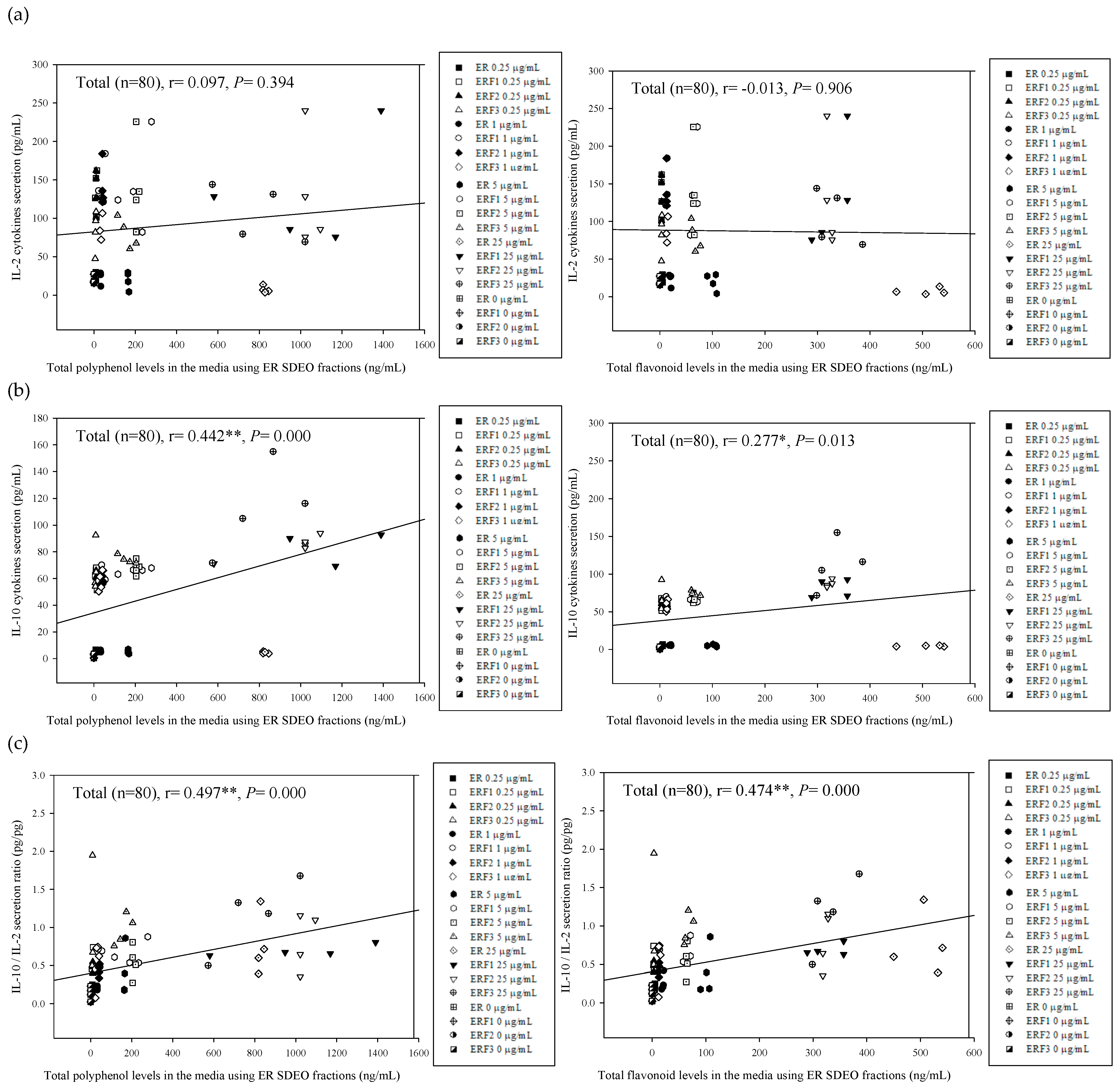
3.6. Associations between Th2 (IL-10)/Th1 (IL-2) Cytokine Secretion Levels in Primary Splenocyte Cultures Treated with ER SDEO as Well as Its Isolated Fractions ERF1–F3 and Total Polyphenol and Flavonoid Contents in Their Corresponding SCM Media
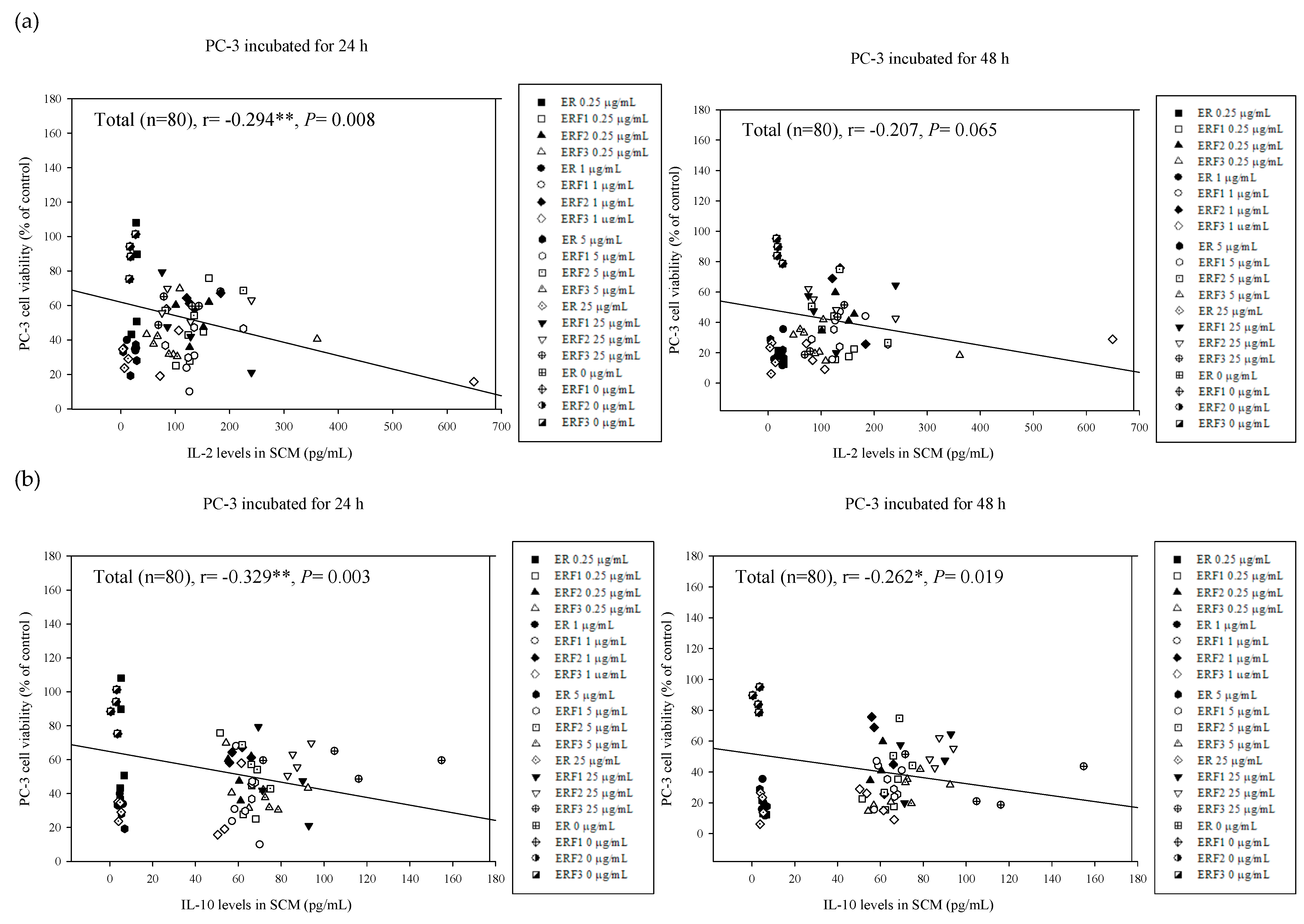
3.7. Associations between PC-3 Cell Viabilities Treated with SCM and Cytokine Secretion Levels in Their Corresponding SCM
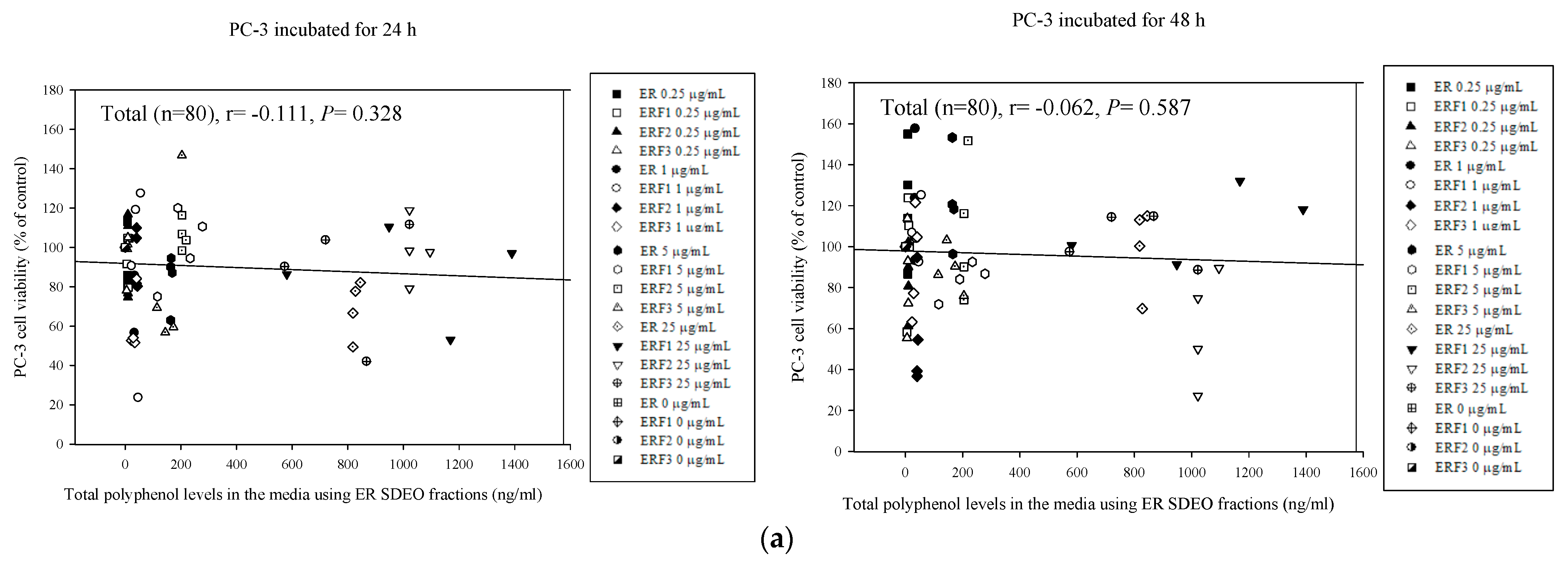
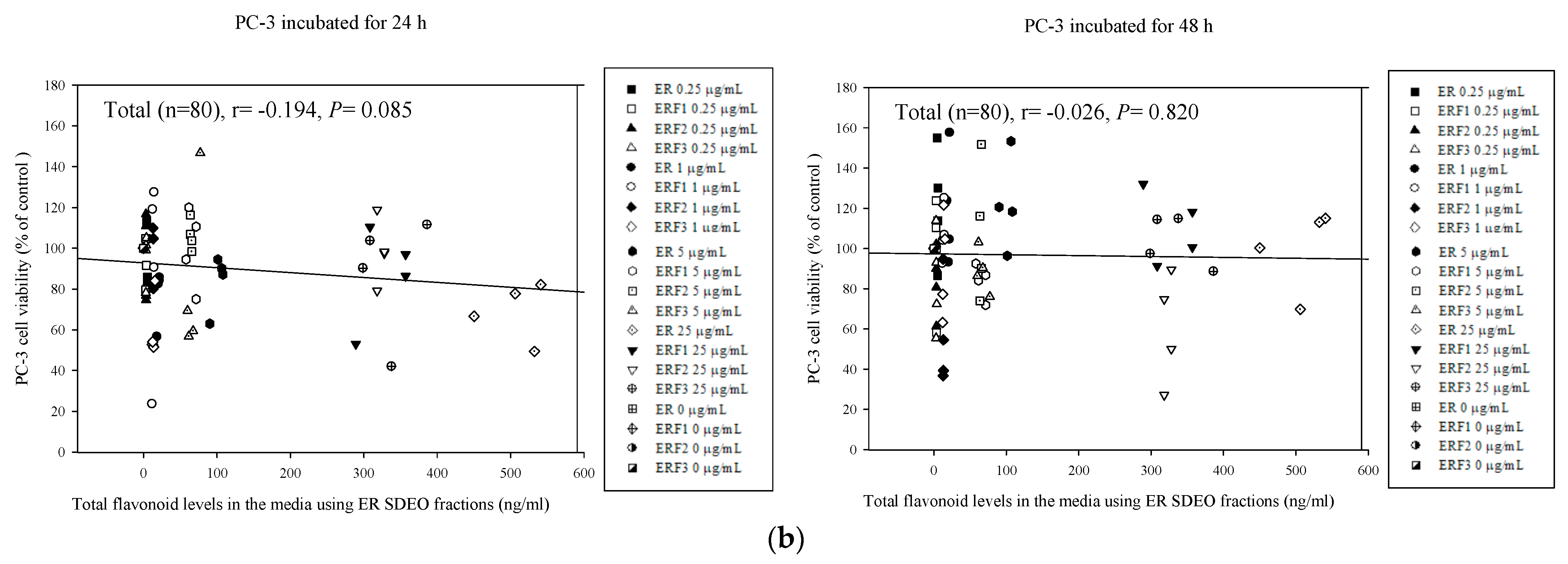
3.8. Associations between PC-3 Cell Viability Treated with Direct Action and Total Polyphenol and Flavonoid Contents in ER SDEO and Its Isolated Fractions ERF1–F3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADT | androgen deprivation therapy |
| Con A | concanavalin A |
| CRPC | castration-resistant prostate cancer |
| DEX | dexamethasone |
| DMSO | dimethyl sulfoxide |
| ELISA | enzyme-linked immuno-sorbent assay |
| ER | Euodia ruticarpa |
| ERF1-F6 | ER SDEO fraction 1–6 |
| GC-MS | gas chromatography–mass spectrometry |
| IL | interleukin |
| LPS | lipopolysaccharide |
| MCM | macrophage-conditioned medium |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| SCM | splenocyte-conditioned medium |
| SDEO | steam distilled essential oil |
| Th1 | T helper type 1 lymphocyte |
| Th2 | T helper type 2 lymphocyte |
| TNF | tumor necrosis factor |
References
- Kimura, T.; Egawa, S. Epidemiology of prostate cancer in Asian countries. Int. J. Urol. 2018, 25, 524–531. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chen, R.; Wang, Y.; Qing, C.; Wang, W.; Yang, Y. In vitro and in vivo efficacy studies of Lavender angustifolia essential oil and its active constituents on the proliferation of human prostate cancer. Integr. Cancer Ther. 2017, 16, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.M.; Abate-Shen, C. Molecular genetics of prostate cancer: New prospects for old challenges. Genes Dev. 2010, 24, 1967–2000. [Google Scholar] [CrossRef] [Green Version]
- Gabriele, F.; Martinelli, C.; Comincini, S. Prostate cancer cells at a therapeutic gunpoint of the autophagy process. J. Cancer Metastasis Treat. 2018, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.C.; Lin, J.Y. Immune cell–conditioned media suppress prostate cancer PC-3 cell growth correlating with decreased proinflammatory/anti-inflammatory cytokine ratios in the media using 5 selected crude polysaccharides. Integr. Cancer Ther. 2016, 15, NP13–NP25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, T.H.; Lin, J.Y. Acorus gramineusand and Euodia ruticarpa steam distilled essential oils exert anti-inflammatory effects through decreasing Th1/Th2 and pro-/anti-inflammatory cytokine secretion ratios in vitro. Biomolecules 2020, 10, 338. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.C.; Lin, J.Y. Pharmacological effects of guava (Psidium guajava L.) seed polysaccharides: GSF3 inhibits PC-3 prostate cancer cell growth through immunotherapy in vitro. Int. J. Mol. Sci. 2021, 22, 3631. [Google Scholar] [CrossRef] [PubMed]
- Yeh, T.H.; Lin, J.Y. Rutaecarpine administration inhibits cancer cell growth in allogenic TRAMP-C1 prostate cancer mice correlating with immune balance in vivo. Biomed. Pharmacother. 2021, 139, 111648. [Google Scholar]
- Rekoske, B.T.; McNeel, D.G. Immunotherapy for prostate cancer: False promises or true hope? Cancer 2016, 122, 3598–3607. [Google Scholar] [CrossRef] [Green Version]
- Conniot, J.; Silva, J.M.; Femandes, J.G.; Silva, L.C.; Gaspar, R.; Brocchini, S.; Florindo, H.F.; Barata, T.S. Cancer immunotherapy: Nanodelivery approaches for immune cell targeting and tracking. Front. Chem. 2014, 2, 105. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, C. Traditional uses, phytochemistry, pharmacology, pharmacokinetics and toxicology of the fruit of Tetradium ruticarpum: A review. J. Ethnopharmacol. 2020, 263, 113231. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Park, C.; Park, S.H.; Hong, S.H.; Kim, G.Y.; Hong, S.H.; Choi, Y.H. Induction of apoptosis by ethanol extract of Evodia rutaecarpa in HeLa human cervical cancer cells via activation of AMP-activated protein kinase. Biosci. Trends 2016, 10, 467–476. [Google Scholar]
- Liao, J.F.; Chiou, W.F.; Shen, Y.C.; Wang, G.J.; Chen, C.F. Anti-inflammatory and anti-infectious effects of Evodia rutaecarpa (Wuzhuyu) and its major bioactive components. Chin. Med. 2011, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Shih, Y.S.; Tsai, C.H.; Li, T.C.; Yu, C.J.; Chou, J.W.; Feng, C.L.; Wang, K.T.; Lai, H.C.; Hsieh, C.L. Effect of wu chu yu tang on gastroesophagel reflux disease: Randomized, double-blind, placebo-controlled trial. Phytomedicine 2019, 56, 118–125. [Google Scholar] [CrossRef]
- Sarto, M.P.M.; da Silva, H.F.L.; de Souza Fernandes, N.; de Abreu, A.P.; Junior, G.Z.; de Ornelas Toledo, M.J. Essential oils from Syzygium aromaticum and Zingiber officinale, administered alone or in combination with benznidazole, reduce the parasite load in mice orally inoculated with Trypanosoma cruzi II. BMC Complement. Med. Ther. 2021, 21, 77. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Grothaus, P.G.; Newman, D. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [PubMed]
- Edris, A.E. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents: A review. Phytother. Res. 2007, 21, 308–323. [Google Scholar] [CrossRef]
- Raut, J.S.; Karuppayil, S.M. A status review on the medicinal properties of essential oils. Ind. Crop. Prod. 2014, 62, 250–264. [Google Scholar]
- Lorenzo-Leal, A.C.; Palou, E.; López-Malo, A.; Bach, H. Antimicrobial, cytotoxic, and anti-inflammatory activities of Pimenta dioica and Rosmarinus officinalis essential oils. Biomed Res. Int. 2019, 2019, 1639726. [Google Scholar] [CrossRef] [Green Version]
- Mogosan, C.; Vostinaru, O.; Oprean, R.; Heghes, C.; Filip, L.; Balica, G.; Moldovan, R.I. A comparative analysis of the chemical composition, anti-inflammatory, and antinociceptive effects of the essential oils from three species of Mentha cultivated in Romania. Molecules 2017, 22, 263. [Google Scholar] [CrossRef]
- Saleh, A.M.; Al-Qudah, M.A.; Nasr, A.; Rizvi, S.A.; Borai, A.; Daghistani, M. Comprehensive analysis of the chemical composition and in vitro cytotoxic mechanisms of Pallines spinosa flower and leaf essential oils against breast cancer cells. Cell. Physiol. Biochem. 2017, 42, 2043–2065. [Google Scholar] [CrossRef]
- Sitarek, P.; Rijo, P.; Garcia, C.; Skała, E.; Kalemba, D.; Białas, A.J.; Szemraj, J.; Pytel, D.; Toma, M.; Wysokińska, H.; et al. Antibacterial, anti-inflammatory, antioxidant, and antiproliferative properties of essential oils from hairy and normal roots of Leonurus sibiricus L. and their chemical composition. Oxidative Med. Cell. Longev. 2017, 2017, 7384061. [Google Scholar] [CrossRef] [Green Version]
- Park, E.; Lee, M.Y.; Seo, C.S.; Jang, J.H.; Kim, Y.U.; Shin, H.K. Ethanol extract of Evodia rutaecarpa attenuates cell growth through Caspase-dependent apoptosis in benign prostatic Hyperplasia-1 cells. Nutrients 2018, 10, 523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, K.C.; Chiu, Y.J.; Tang, Y.J.; Lin, K.L.; Chiang, J.H.; Jiang, Y.L.; Jen, H.F.; Kuo, Y.H.; Agamaya, S.; Chung, J.G. Houttuynia cordata Thunb extract inhibits cell growth and induces apoptosis in human primary colorectal cancer cells. Anticancer Res. 2010, 30, 3549–3556. [Google Scholar] [PubMed]
- Ayeka, P.A.; Bian, Y.; Githaiga, P.M.; Zhao, Y. The immunomodulatory activities of licorice polysaccharides (Glycyrrhiza uralensis Fisch.) in CT 26 tumor-bearing mice. BMC Complem. Altern. Med. 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kou, W.Z.; Yang, J.; Yang, Q.H.; Wang, Y.; Wang, Z.F.; Xu, S.L.; Liu, J. Study on in-vivo anti-tumor activity of Verbena officinalis extract. Afr. J. Tradit. Complem. Altern. Med. 2013, 10, 512–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeson, P.D.; Springthorpe, B. The influence of drug-like concepts on decision-making in medicinal chemistry. Nat. Rev. Drug Discov. 2007, 6, 881–890. [Google Scholar] [CrossRef]
- Lin, J.Y.; Li, C.Y.; Hwang, I.F. Characterization of the pigment components in red cabbage (Brassica oleracea L. var.) juice and their anti-inflammatory effects on LPS-stimulated murine splenocytes. Food Chem. 2008, 109, 771–781. [Google Scholar] [CrossRef]
- Lin, J.Y.; Tang, C.Y. Strawberry, loquat, mulberry, and bitter melon juices exhibit prophylactic effects on LPS-induced inflammation using murine peritoneal macrophages. Food Chem. 2008, 107, 1587–1596. [Google Scholar] [CrossRef]
- Lin, H.C.; Lin, J.Y. Characterization of guava (Psidium guajava Linn) seed polysaccharides with an immunomodulatory activity. Int. J. Biol. Macromol. 2020, 154, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Lin, J.Y. Five bitter compounds display different anti-inflammatory effects through modulating cytokine secretion using mouse primary splenocytes in vitro. J. Agric. Food Chem. 2011, 59, 184–192. [Google Scholar] [CrossRef]
- Lin, H.C.; Lin, J.Y. M1 polarization but anti-LPS-induced inflammation and anti-MCF-7 breast cancer cell growth effects of five selected polysaccharides. Evid-Based Complement. Altern. Med. 2020, 2020, 9450246. [Google Scholar] [CrossRef]
- Lin, H.C.; Lin, J.Y. GSF3, a polysaccharide from guava (Psidium guajava L.) seeds, inhibits MCF-7 breast cancer cell growth via increasing Bax/Bcl-2 ratio or Fas mRNA expression levels. Int. J. Biol. Macromol. 2020, 161, 1261–1271. [Google Scholar] [CrossRef]
- Ku, C.M.; Lin, J.Y. Anti-inflammatory effects of 27 selected terpenoid compounds tested through modulating Th1/Th2 cytokine secretion profiles using murine primary splenocytes. Food Chem. 2013, 141, 1104–1113. [Google Scholar] [CrossRef]
- Yang, C.; Chen, H.; Chen, H.; Zhong, B.; Luo, X.; Chun, J. Antioxidant and anticancer activities of essential oil from Gannan navel orange peel. Molecules 2017, 22, 1391. [Google Scholar] [CrossRef]
- Chang, W.L.; Cheng, F.C.; Wang, S.P.; Chou, S.T.; Shih, Y. Cinnamomum cassia essential oil and its major constituent cinnamaldehyde induced cell cycle arrest and apoptosis in human oral squamous cell carcinoma HSC-3 cells. Environ. Toxicol. 2017, 32, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Mekhail, T.M.; Markman, M. Paclitaxel in cancer therapy. Expert Opin. Pharmacother. 2002, 3, 755–766. [Google Scholar] [PubMed]
- Taherian, M.; Norenberg, M.D.; Panickar, K.S.; Shamaladevi, N.; Ahmad, A.; Rahman, P.; Jayakumar, A.R. Additive effect of resveratrol on astrocyte swelling post-exposure to ammonia, ischemia and trauma in vitro. Neurochem. Res. 2020, 45, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Dietary polyphenols: Good, bad, or indifferent for your health? Cardiovasc. Res. 2007, 73, 341–347. [Google Scholar] [CrossRef]
- Mennen, L.I.; Walker, R.; Bennetau-Pelissero, C.; Scalbert, A. Risks and safety of polyphenol consumption. Am. J. Clin. Nutr. 2005, 81 (Suppl. 1), 326S–329S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ofosu, F.K.; Daliri, E.B.M.; Elahi, F.; Chelliah, R.; Lee, B.H.; Oh, D.H. New insights on the use of polyphenols as natural preservatives and their emerging safety concerns. Front. Sustain. Food Syst. 2020, 4, 223. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Nugraha, A.S.; Triatmoko, B.; Wangchuk, P.; Keller, P.A. Vascular epiphytic medicinal plants as sources of therapeutic agents: Their ethnopharmacological uses, chemical composition, and biological activities. Biomolecules 2020, 10, 181. [Google Scholar] [CrossRef] [Green Version]
- Maaliki, D.; Shaito, A.A.; Pintus, G.; El-Yazbi, A.; Eid, A.H. Flavonoids in hypertension: A brief review of the underlying mechanisms. Curr. Opin. Pharmacol. 2019, 45, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.R.; Appe, C.L. Polyphenols as dietary supplements: A double-edged sword. Nutr. Diet. Suppl. 2010, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lin, B.F.; Chiang, B.L.; Lin, J.Y. Amaranthus spinosus water extract directly stimulates proliferation of B lymphocyte in vitro. Int. Immunopharmacol. 2005, 5, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lu, P.; Li, B.; Zhang, W.; Yang, R.; Chu, Y.; Luo, K. Interleukin 2 and interleukin 10 function synergistically to promote CD8+ T cell cytotoxicity, which is suppressed by regulatory T cells in breast cancer. Int. J. Biochem. Cell Biol. 2017, 87, 1–7. [Google Scholar] [CrossRef]
- Santin, A.D.; Hermonat, P.L.; Ravaggi, A.; Bellone, S.; Pecorelli, S.; Roman, J.J.; Parham, G.P.; Cannon, M.J. Interleukin-10 increases Th1 cytokine production and cytotoxic potential in human papillomavirus-specific CD8+ cytotoxic T lymphocytes. J. Virol. 2000, 74, 4729–4737. [Google Scholar] [CrossRef] [Green Version]
- Trueblood, E.S.; Brown, W.C.; Palmer, G.H.; Davis, W.C.; Stone, D.M.; McElwain, T.F. B-lymphocyte proliferation during bovine leukemia virus-induced persistent lymphocytosis is enhanced by T-lymphocyte-derived interleukin-2. J. Virol. 1998, 72, 3169–3177. [Google Scholar] [CrossRef] [Green Version]
- Tai, S.; Sun, Y.; Squires, J.M.; Zhang, H.; Oh, W.E.; Liang, C.Z.; Huang, J. PC3 is a cell line characteristic of prostatic small cell carcinoma. Prostate 2011, 71, 1668–1679. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liang, Y.; He, C. Anticancer activities and mechanisms of heat-clearing and detoxicating traditional Chinese herbal medicine. Chin. Med. 2017, 12, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Vong, C.T.; Chen, H.; Gao, Y.; Lyu, P.; Qiu, L.; Zhao, M.; Liu, Q.; Cheng, Z.; Zou, J.; et al. Naturally occuring anti-cancer compounds: Shining from Chinese herbal medicine. Chin. Med. 2019, 14, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiina, Y.; Funabashi, N.; Lee, K.; Toyoda, T.; Sekine, T.; Honjo, S.; Hasegawa, R.; Kawata, T.; Wakatsuki, Y.; Hayashi, S. Relaxation effects of lavender aromatherapy improve coronary flow velocity reserve in healthy men evaluated by transthoracic doppler echocardiography. Int. J. Cardiol. Res. 2008, 129, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Tahvildari, M.; Dana, R. Low-dose IL-2 therapy in transplantation, autoimmunity, and inflammatory diseases. J. Immunol. 2019, 203, 2749–2755. [Google Scholar] [CrossRef]
- Kirkwood, J.M.; Butterfield, L.H.; Tarhini, A.A.; Zarour, H.; Kalinski, P.; Ferrone, S. Immunotherapy of cancer in 2012. CA Cancer J. Clin. 2012, 62, 309–335. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total Flavonoids (mg Quercetin Equivalent/g Sample) | Total Polyphenols (mg Gallic Acid Equivalent/g Sample) |
|---|---|---|
| ERF1 | 13.1 ± 1.4 a,b | 40.9 ± 13.8 a |
| ERF2 | 12.9 ± 0.2 a,b | 41.6 ± 1.5 a |
| ERF3 | 13.3 ± 1.6 a,b | 31.8 ± 17.7 a,b |
| ERF4 | 14.4 ± 0.7 a | 10.7 ± 1.5 b,c |
| ERF5 | 12.4 ± 0.8 b,c | 4.8 ± 1.4 c |
| ERF6 | 11.4 ± 0.5 c | 24.0 ± 10.6 b |
| Fractions | NO. | RT (min) | RI | Compounds | M.W. | Chemical Formula | CAS NO. |
|---|---|---|---|---|---|---|---|
| F1 | 1 | 33.510 | 1536.44 | 1-Dodecanethiol | 202.18 | C12H26S | 000112-55-0 |
| 2 | 47.825 | 2089.37 | 2-[3-Methoxyphenyl]-4H-1-benzopyran-4-one | 252.08 | C16H12O3 | 007622-32-4 | |
| 3 | 53.663 | 2364.47 | Cobalt(I), cyclopentadienyl-(η4-cis-5,6-diethylcyclohex-1,3-diene) | 260.10 | C15H21CO | 000000-00-0 | |
| 4 | 53.721 | 2367.58 | N,N′-Diphenyl-1,4-benzenediamine | 260.13 | C18H16N2 | 000074-31-7 | |
| 5 | 54.934 | 2427.84 | Dehydroabietic acid | 300.21 | C20H28O2 | 001740-19-8 | |
| 6 | 62.832 | 2779.08 | Erucylamide | 337.33 | C22H43NO | 000112-84-5 | |
| F2 | 1 | 47.823 | 2089.37 | 2-[3-Methoxyphenyl]-4H-1-benzopyran-4-one | 252.08 | C16H12O3 | 053906-83-5 |
| 2 | 53.658 | 2364.47 | Cobalt(I), cyclopentadienyl-(η4-cis-5,6-diethylcyclohex-1,3-diene) | 260.10 | C15H21Co | 000000-00-0 | |
| 3 | 53.716 | 2367.58 | N,N′-Diphenyl-1,4-benzenediamine | 260.13 | C18H16N2 | 000074-31-7 | |
| 4 | 62.781 | 2775.81 | 9-Octadecenamide | 281.27 | C18H35NO | 000301-02-0 | |
| F3 | 1 | 44.625 | 1952.63 | Palmitic acid | 256.24 | C16H32O2 | 000057-10-3 |
| 2 | 47.823 | 2089.37 | 2-[3-Methoxyphenyl]-4H-1-benzopyran-4-one | 252.08 | C16H12O3 | 053906-83-5 | |
| 3 | 48.701 | 2129.13 | Oleic acid | 282.26 | C18H34O2 | 000112-80-1 | |
| 4 | 49.833 | 2180.61 | 1,1-Diphenyl-3-methyl-1-silacyclopent-3-ene | 250.12 | C17H18Si | 051343-48-7 | |
| 5 | 53.371 | 2350.27 | Tetradecanoic acid | 256.24 | C16H32O2 | 000124-06-1 | |
| 6 | 53.659 | 2364.47 | Cobalt(I), cyclopentadienyl-(η4-4-cis-5,6-diethylcyclohex-1,3-diene) | 260.10 | C15H21CO | 000000-00-0 | |
| 7 | 53.718 | 2367.58 | N,N′-Diphenyl-1,4-benzenediamine | 260.13 | C18H16N2 | 000074-31-7 | |
| 8 | 55.087 | 2435.53 | Benzoic acid | 314.12 | C18H18O5 | 000120-55-8 | |
| 9 | 55.214 | 2441.89 | N-Propylbenzamide | 163.10 | C10H13NO | 010546-70-0 | |
| 10 | 55.701 | 2466.14 | Dipropylene glycol dibenzoate | 342.15 | C20H22O5 | 020109-39-1 | |
| 11 | 55.902 | 2476.09 | 2,2,4,5-Tetramethyl-6-(1-methyloctadecyl)-1,3-dioxane | 410.41 | C27H54O2 | 056324-82-4 | |
| 12 | 62.861 | 2779.08 | Erucylamide | 337.33 | C22H43NO | 000112-84-5 | |
| F4 | 1 | 44.607 | 1952.63 | Palmitic acid | 256.24 | C16H32O2 | 000057-10-3 |
| 2 | 55.893 | 2476.09 | 2,2,4,5-Tetramethyl-6-(1-methyloctadecyl)-1,3-dioxane | 410.41 | C27H54O2 | 056324-82-4 | |
| 3 | 57.294 | 2540.93 | 2-Propenoic acid, 2-ethylhexyl ester | 184.15 | C11H20O2 | 000103-11-7 | |
| 4 | 62.335 | 2775.81 | 9-Octadecenamide | 281.27 | C18H35NO | 000301-02-0 | |
| 5 | 62.865 | 2779.08 | Erucylamide | 337.33 | C22H43NO | 000112-84-5 | |
| F5 | 1 | 62.724 | 2775.81 | 9-Octadecenamide | 281.27 | C18H35NO | 000301-02-0 |
| F6 | 1 | 47.825 | 2089.08 | 2,4-Dibromo-phenol | 249.86 | C6H4OBr2 | 000615-58-7 |
| 2 | 49.475 | 2164.05 | 1,3-Bis(4-methylphenyl)-1,3-propanedione | 252.12 | C17H16O2 | 003594-36-3 | |
| 3 | 49.833 | 2180.61 | 1,1-Diphenyl-3-methyl-1-silacyclopent-3-ene | 250.12 | C17H18Si | 051343-48-7 | |
| 4 | 53.669 | 2364.44 | Titanium | 260.10 | C16H20Ti | 000000-00-0 | |
| 5 | 53.721 | 2367.58 | N,N′-Diphenyl-1,4-benzenediamine | 260.13 | C18H16N2 | 000074-31-7 | |
| 6 | 57.299 | 2540.50 | 4-(6-Methoxy-2-quinolyl)-benzonitrile | 260.10 | C17H12N2O | 000000-00-0 | |
| 7 | 62.865 | 2779.08 | Erucylamide | 337.33 | C22H43NO | 000112-84-5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, T.-H.; Lin, J.-Y. Active Ingredients from Euodia ruticarpa Steam Distilled Essential Oil Inhibit PC-3 Prostate Cancer Cell Growth via Direct Action and Indirect Immune Cells Conditioned Media In Vitro. Curr. Issues Mol. Biol. 2021, 43, 996-1018. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43020071
Yeh T-H, Lin J-Y. Active Ingredients from Euodia ruticarpa Steam Distilled Essential Oil Inhibit PC-3 Prostate Cancer Cell Growth via Direct Action and Indirect Immune Cells Conditioned Media In Vitro. Current Issues in Molecular Biology. 2021; 43(2):996-1018. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43020071
Chicago/Turabian StyleYeh, Tzu-He, and Jin-Yuarn Lin. 2021. "Active Ingredients from Euodia ruticarpa Steam Distilled Essential Oil Inhibit PC-3 Prostate Cancer Cell Growth via Direct Action and Indirect Immune Cells Conditioned Media In Vitro" Current Issues in Molecular Biology 43, no. 2: 996-1018. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43020071