3. Results
The baseline characteristics include age, gender, weight, height, BMI, adipose tissue mass, skeletal muscles mass, and visceral fat levels. Body weight and BMI values did not differ significantly before and after the therapy (
Table 3).
According to the World Health Organization interpretation of the BMI values, all subjects were obese to varying degrees. At the same time, this observation was also true for the interpretation of the BMI values after the therapy. After the end of the 3 months of vitamin D supplementation, the value of this index did not change significantly. Among the individuals, both before and after vitamin D supplementation, the majority were patients with first-degree obesity (before: 39.39%; after: 36.36%) and second-degree obesity (before: 36.36%; after: 39.39%). After the therapy, the percentage of people with first-degree obesity (39.39% vs. 36.36%, before vs. after) and third-degree (21.21% vs. 18.18%, before vs. after) slightly decreased. With second-degree obesity (36.36% vs. 39.39%, before vs. after) and overweight (3.03% vs. 6.06%, before vs. after) it slightly increased. The anthropometric features assessed before and after vitamin D supplementation also included skeletal muscle mass, adipose tissue mass, % of adipose tissue, and the level of visceral fat (
Table 4).
Results for any of the four characteristics listed (skeletal muscle mass, adipose tissue mass, % of adipose tissue, and visceral fat levels) did not show significant differences between the time points 0 and 1. The concentration of vitamin D 25-(OH) in all subjects before the therapy equaled, accordingly, an average level of 18.22 ± 1.103 ng/mL, and after the therapy it increased to the level of 29.89 ± 1.160 ng/mL. The Wilcoxon pair test was statistically significant (p < 0.001).
In
Table 5 and
Table 6, divided results for women and men are presented. In
Table 7 we have shown Vitamin D concentration before and after the 3-month supplementation.
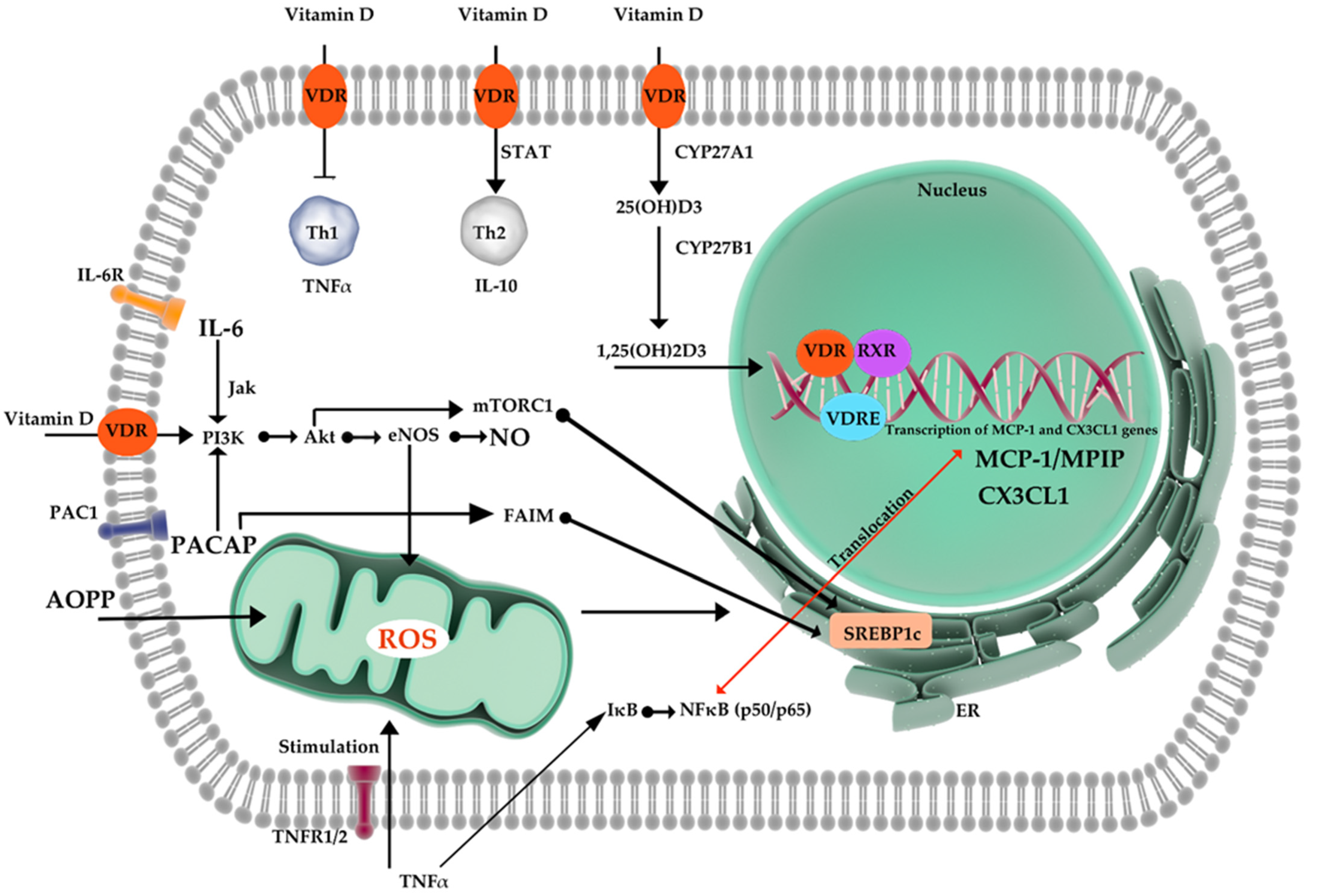
In the next step of our experiment, molecular parameters involved in the inflammatory process were determined. Our measurements included interleukin-6 (IL-6), Pituitary adenylate cyclase-activating peptide (PACAP), Advanced oxidation protein products (AOPP), C-X3-C Motif Chemokine Ligand 1 (CX3CL1), Monocyte Chemoattractant Protein-1 (MCP-1), and nitric oxide (NO). All parameters were compared between two time points (Time point 0 vs. time point 1). MCP-1, IL-6, CX3CL1, and PACAP did not show significant differences before and after vitamin D therapy. Results have been presented in
Table 8.
The AOPP concentration in the subjects before the therapy reached the average level of 55.07 ± 12.21 nmoL/mL, and after the therapy it increased to the level of 58.94 ± 12.10 nmoL/mL. The difference between the mean concentrations of AOPP before and after treatment (3.87), as shown by the Wilcoxon pair test, was statistically significant (p = 0.047). The concentration of NO in all subjects before the therapy reached the average level of 39.19 ± 10.96 µmoL/L, and after the therapy it increased to the level of 70.02 ± 13.80 µmoL/L. The difference between the mean concentrations of NO before and after treatment (30.83) µmoL/L), as shown by the Wilcoxon pair test, was statistically significant (p = 0.021).
The Spearman r correlation analysis between vitamin D concentration before its supplementation and the concentration of molecular parameters (MCP1, IL-6, NO, CX3CL1, AOPP, and PACAP) before Vitamin D therapy did not show any significant relations between the studied values (
Table 9).
The Spearman r correlation analysis between vitamin D concentration after its supplementation and the concentration of molecular parameters (MCP1, IL-6, NO, CX3CL1, AOPP, and PACAP) after the 3-month Vitamin D therapy did not show any significant relations between the studied values (
Table 10).
4. Discussion
Nitric oxide’s modulatory influence on vascular and immune function is beyond discussion. Vast experimental evidence presents NO as a potentially protective agent as well as an unbeneficial one, related to oxidative injuries [
52,
53]. The nature of vitamin D’s impact on NO signaling remains controversial. Calcitriol is known to inhibit LPS-induced immune activation in human endothelial cells [
54]. Activation of 1α-hydroxylase in macrophages elevates the level of calcitriol, which inhibits the iNOS expression and reduces NO production within LPS-stimulated macrophages [
55]. In that case, calcitriol production by macrophages may constitute a protective mechanism against the oxidative injuries that are caused by the so-called NO burst. The same effect on iNOS has been observed in rat CNS during an experimental model of allergic encephalomyelitis [
56]. Conversely, according to Andrukhova et. al., vitamin D has a potential to improve endothelial function and health by an increase in signaling for the transcription of endothelial nitric oxide synthase (eNOS) [
50]. Studies in mice have proven that subjects deprived of the eNOS and/or nNOS gene exhibit metabolic syndrome, leading to possible vascular consequences [
57,
58].
In the current investigation, NO concentrations in the studied patients reached an average level of 39.19 ± 10.96 µmoL/L prior to vitamin D supplementation. Subsequent therapy increased its levels to 70.02 ± 13.80 µmoL/L in a statistically significant manner (
p = 0.021). Previous studies have shown that vitamin D may upregulate eNOS expression and increase NO bioavailability. Vitamin D administration in the study of Abeer M. Mahmoud et al. improved flow-induced and acetylcholine-induced dilation of arterioles isolated from adipose and subcutaneous tissues of bariatric patients. The effects have been connected with increased NO production in the resistance arterioles. Achieved improvement in vessels’ reactivity was diminished by eNOS inhibition via NO synthase inhibitor- N(ω)-nitro-L-arginine methyl ester (L-NAME). Interestingly, researchers noted that vitamins D actions were more significant before the bariatric surgery and consequent weight loss [
59]. The extent of the therapeutical response to the vitamin D treatment may be related to skin pigmentation. Due to UV ray-absorbing properties, melanin and its relatively higher concentrations can contribute to impaired vitamin D production in adults with darker pigmentation, placing such populations at risk for vitamin D deficiency [
60,
61,
62]. Reduced cutaneous microvascular vasodilation in response to local heating and reduced nitric oxide (NO) contribution to that response have been observed in college-aged African Americans (AA) compared to European American (EA) adults [
63,
64,
65]. It is possible that modulation of NO-mediated signaling can be utilized to limit the discrepancies. Wolf, S. T. et al. reported a mitigation of such differences after 4 weeks of 2000 IU/day oral vitamin D supplementation. They achieved significant improvement in serum 25(OH)D concentrations of AA (from 17.93 ± 5.24 to 26.07 ± 3.73 ng/mL,
p = 0.04; g = 1.66) but not of EA (
p = 0.16). Vitamin D supplementation increased the NO contribution to the local heating-induced vessel dilation in AA (from pre- to post-supplementation (29.83 ± 13.70 vs. 46.79 ± 21.93% max,
p = 0.01; g = 0.89)), abolishing the difference between groups (
p = 0.47). Its assessment was possible thanks to L-NAME administration and subsequent NO-dependent vasodilation (%NO) quantification [
66]. The report of Harris et al. seems to be consistent in terms of vascular effect. Sixteen-week-long supplementation of 60,000 IU monthly oral vitamin D3 (~2000 IU/day) proved to be effective at improving vascular endothelial function in AA adults [
67]. Although the observed increase in NO concentrations appears to be statistically significant in the present study, the Spearman r correlation analysis did not show any significant relations between the values of NO and vitamin D concentration after its supplementation. A similar neutral effect on nitric oxide has been observed in PCOS women. The condition makes the suffering prone to multiple metabolic disorders [
68,
69,
70]. A meta-analysis of Akbari et al. concluded that vitamin D supplementation resulted in a significant decrease in hs-CRP, but did not affect NO levels [
71].
The second parameter in which significant increase might be observe is AOPP. The difference between the mean concentrations of AOPP before and after treatment (3.87), as shown by the Wilcoxon pair test, was statistically relevant (
p = 0.047). Characterized for the first time by Witko-Sarsat et al. [
72], advanced oxidation protein products (AOPPs) are considered adequate markers of oxidative stress and related tissue injury. Increased AOPP levels have been reported in both obesity and diabetes [
73,
74]. Koçak et al. demonstrated that serum AOPP levels were significantly elevated in obese and diabetic women compared to healthy control. In all cases, AOPP levels have been positively correlated with blood glucose concentration and age [
75]. A simultaneous relative increase in IL-6 levels could be observed, which supports previous findings associating women’s obesity and IL-6 elevation [
76,
77]. In the study of Oliveira et al. assessing AOPP, they showed a relation between oxidative and nitrosative stresses and vitamin D deficiency in multiple sclerosis patients. Researchers observed decreased levels of AOOP in patients with 25(OH)D concentrations lower than 20 ng/mL than in those with ≥20 ng/mL (133.83 ± 58.95 vs. 164.99 ± 91.40,
p = 0.046). However, after further analysis, it remained statistically irrelevant [
78]. Gradinaru et al. found significantly negative associations between vitamin D status and the susceptibility of LDL to oxidation, the AOPPs, and certain cardiovascular risk biochemical markers like AGEs and nitric oxide metabolic pathway products (NOx) in the elderly, mostly obese patients with IFG and/or T2DM [
79]. After correlation analysis in our own study, there was no relation between vitamin D supplementation and post-therapy concentrations of AOPP.
For the first time, a report on a potentially advantageous impact of a sufficient vitamin D level on inflammatory status in humans was presented by Jablonski et al. [
80]. They found that vascular endothelial cell expression of the p65 subunit of NFκB, being a major pro-inflammatory nuclear transcription factor, and IL-6, a pro-inflammatory cytokine and downstream target of NFκB, was higher in 25(OH)D deficient patients when compared to middle-aged/older adults who were sufficient in that manner. In the study of our own, measurements of MCP-1, IL-6, PACAP, and CX3CL1 concentrations did not show significant differences before and after vitamin D therapy. Ni et. al., in their research conducted on diabetic rats, demonstrated that vitamin D treatment decreases significantly hepatic expression of pro-inflammatory mediators such as NF-κB and MCP-1 [
49]. Wamberg et al., in a series of studies, reported that incubation with 1,25(OH)2D decreased the expression of MCP-1, IL-6, and IL-8 and reduced IL-8 protein secretion in human adipose tissue (AT) fragments [
81]. In the follow-up evaluation, supplementation with a daily dose of 7000 IU of vit. D for 26 weeks did not affect the expression of the inflammatory markers in AT or the concentration of circulating inflammatory markers [
82]. Similar discrepancies have been observed in numerous studies. On one hand, treatment for 3 years with 700 IU vitamin D plus 500 mg of calcium daily showed no effect of on circulating levels of IL-6 or CRP in healthy adults [
83]. Similarly, in the research of Jorde et al. in obese human patients, a 1-year-long treatment with a weekly dose of 40,000 IU of cholecalciferol had no effect on hsCRP, MCP-1, or several other markers of systemic inflammation such as IL-2, IL-4, IL-5, IL-10, IL-12IL-13, IL-17, intercellular adhesion molecule-1, and interferon-gamma [
84]. Furthermore, Calton et al. found a lack of benefit with vitamin D supplementation on inflammatory cytokines such as CRP and IL-6 [
85]. On the other hand, vitamin D supplementation with 3332 IU daily for 1 year during a weight loss trial of obese subjects enhanced the decrease in TNFα but not CRP or IL-6, compared to placebo [
86]. The strongest conclusions regarding the issue can be drawn on the basis of Jamka et al.’s metanalysis, including data from 13 randomized, controlled trials and 1955 overweight and obese subjects [
87]. The vitamin D supplementation did not have an influence on CRP (std. mean differences −0.11; 95% CI −0.27–0.04;
p = 0.15), TNF-α (std. mean differences −0.13; 95% CI −0.38–0.12;
p = 0.31), and IL-6 concentrations (std. mean differences 0.1; 95% CI −0.43–0.63;
p = 0.71). In the metanalysis, IL-6 concentrations were evaluated in eight studies. At baseline, in vitamin D groups, the average plasma concentrations of IL-6 varied between 1.00 and 8.90 pg/mL. There was no statistical significance of vitamin D supplementation on plasma concentrations of IL-6 (std. mean differences 0.1; 95% CI −0.43–0.63;
p = 0.71). The authors brought to attention that reported results might have been affected by differences in ethnicity, advanced age, and sex age [
87,
88].
Fractalkine is expressed in adipocytes and has been shown to promote monocyte adhesion in adipose tissue from obese patients [
89]. It takes part in the regulation of pancreatic islet β cell function and insulin secretion [
90]. There is vast evidence that fractalkine/CX3CL1 is actively involved in many states related to inflammation such as atherosclerosis, HIV infection, or cancer [
14]. Recent studies prove that serum fractalkine levels may constitute a potent diagnostic marker of childhood onset of SLE, independently of stage of lupus-related nephritis [
91]. In the studies of Shinzari et al., subgroups of obese patients presented impaired reactivity to nitric oxide-dependent vasodilator stimuli and enhanced ETA-dependent vasoconstrictor tone when increased circulating fractalkine levels were observed. The authors noted that vascular function impairment may be directly linked to increased plasma concentrations of atherogenic substances including fractalkine [
92,
93]. Thus, increased LDL-cholesterol or reduced HDL-cholesterol have been reported to elevate fractalkine’s levels. They proposed the presence of lipid abnormalities as a potential key factor in the underlying mechanism of its influence [
94,
95,
96]. In the study of Yegorov et al., 21 cytokines were measured in serum at baseline and after 6 months of vitamin D supplementation in deficient children. Vitamin D deficiency was linked to the reduction of such chemokines as MIP-1α (CCL3) and IL-8 (CXCL8). This relation was not observed with fractalkine. The median blood 25(OH)D concentration at baseline was 13.7 nmol/L (IQR = 10.0–21.7). Supplementation tripled blood 25(OH)D levels (
p < 0.001) [
97]. Another similar trial, conducted by Davaasambuu et al., also in children, showed no statistically significant elevation of various cytokines (IL-1b, 2, 4, 5, 6, 7, 8, 10, 12p70, 13, 17a, 21, 23, GM-CSF, IFN-γ, TNF-α, ITAC, Fractalkine, MIP1a, MIP1b, MIP3a) except the levels of granulocyte–macrophage colony-stimulating factor (GM-CSF) following vitamin D oral supplementation [
98].
Unfortunately, data available in literature regarding vitamin’s D influence on PACAP are limited. Nevertheless, PACAP relation with inflammatory processes and obesity-related metabolic changes has been previously described [
99,
100]. It is expressed both in the peripheral and central nervous systems. The cells of the mediobasal hypothalamus (MBH) contain PACAP in the ventromedial hypothalamic nucleus (VMN), which serves a key role in modulating sympathetic nervous system activation to regulate glucose metabolism, energy expenditure, thermogenesis, and satiety [
99]. The study of Green et al. showed that 14 days of treatment with the PACAP receptor antagonist had an unbeneficial effect on glucose tolerance and insulin sensitivity in obese diabetic ob/ob mice [
101]. Although details of PACAP’s influence remain unclear, it has been proven that PACAP mediates sympathetic effects of leptin in a tissue-specific manner. The data brought by Tanida et al. suggest that PACAP signaling is connected to leptin’s control of feeding patterns and lipocatabolic sympathetic outflow. What is interesting is that the renal sympathetic traffic appears to be spared and not affected [
102].
Despite many reports regarding the abovementioned markers and therapies, the primary outcomes remain the most important issues for clinical practitioners. There are many findings implying the efficacy of vitamin D supplementation in diseases and processes related to inflammation (reviewed in [
103]). Levels of vitamin D higher than 50 mmol/L were connected with reduced risk of prostate and colorectal cancer by 30–50% [
104,
105]. Martineau et al., in their metanalysis, presented evidence on a possible protective effect of vitamin D against acute renal injury [
106]. Some recent studies have presented data about differences in vitamin D dosages and their influence on the course of COVID-19 and the risk of developing severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) [
107,
108]. Vitamin D deficiency may be related to other inflammatory-based diseases such as allergies. Negative correlation between low concentrations of 25(OH)D and IgE in the serum of studied children have been observed [
109]. What is more, antenatal supplementation of vitamin D preparation has been shown to decrease a risk of food allergies in infants with GT/TT genotype [
110].
Our study has some potential limitations, within which our findings should be interpreted carefully. First, as in much quantitative research, was a relatively modest sample size and lack of probability sampling due to a strong regional focus of the studied population. Second, body mass composition was determined with the use of an impedance body composition analyzer rather than utilizing techniques based on magnetic resonance, computer tomography, or even near infrared spectroscopy. Although certain inclusion and exclusion criteria were applied, there were factors with the potential impact, such as medications other that hormonal treatment, that did not affect the qualification process. All the above remarks need to be taken into consideration in the follow-up studies in this field, which, hopefully, will be free from restrictions resulting from the pandemic period and related limitations in the collection of data.


 ,
, 

{kind=link}