
Antidiabetic Effect of Taxifolin in Cultured L6 Myotubes and Type 2 Diabetic Model KK-Ay/Ta Mice with Hyperglycemia and Hyperuricemia


Abstract
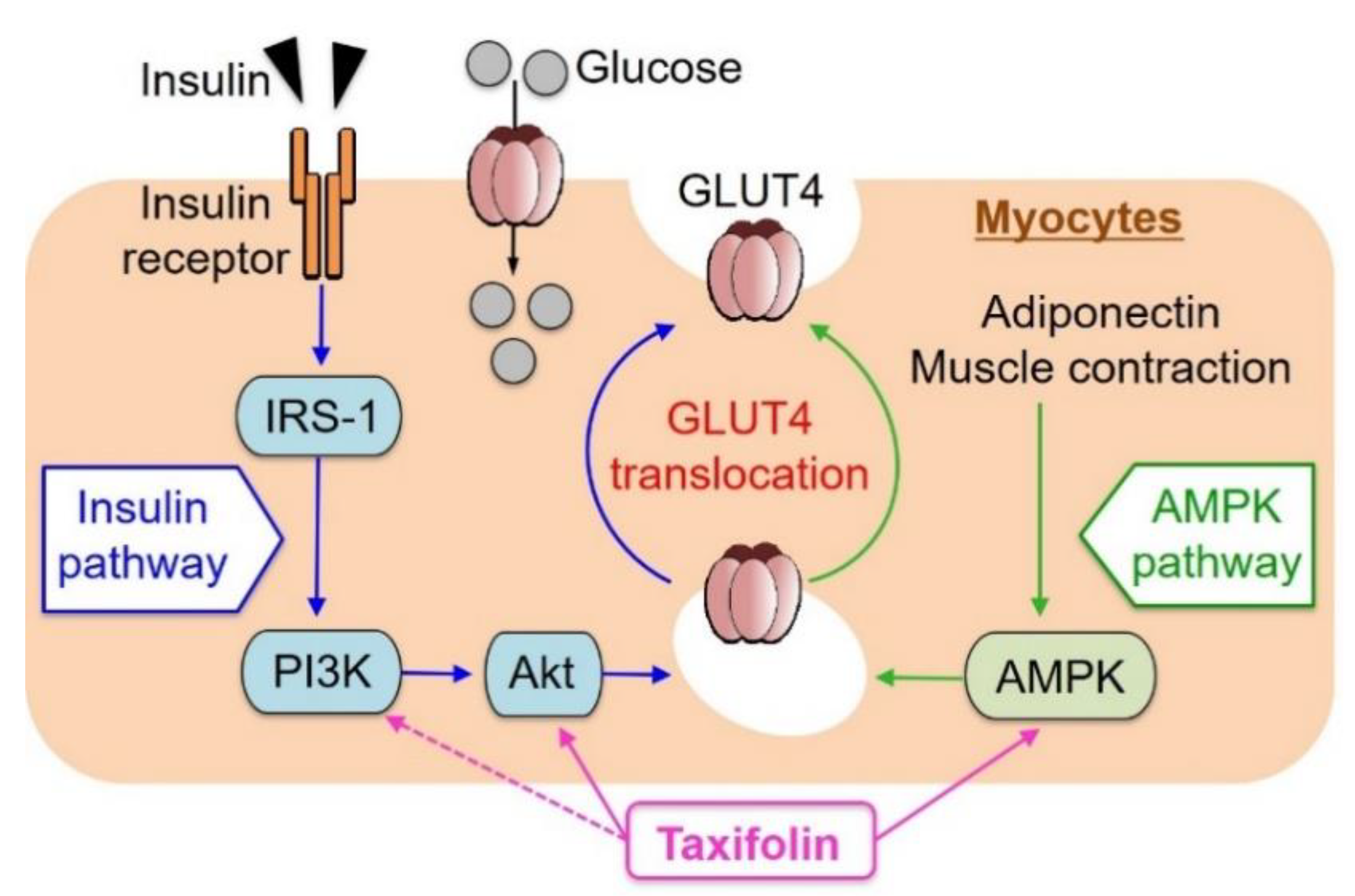
:1. Introduction
2. Materials and Methods
2.1. Evaluation of Glucose Uptake in L6 Myotubes
2.2. Protein Extraction from L6 Myotubes
2.3. Protein Extraction of Plasma Membrane from L6 Myotubes
2.4. Western Blotting
2.5. Animal Experiments
2.6. Time-Dependent Evaluation of Blood Glucose Levels
2.7. Dissection
2.8. Statistical Analysis
3. Results
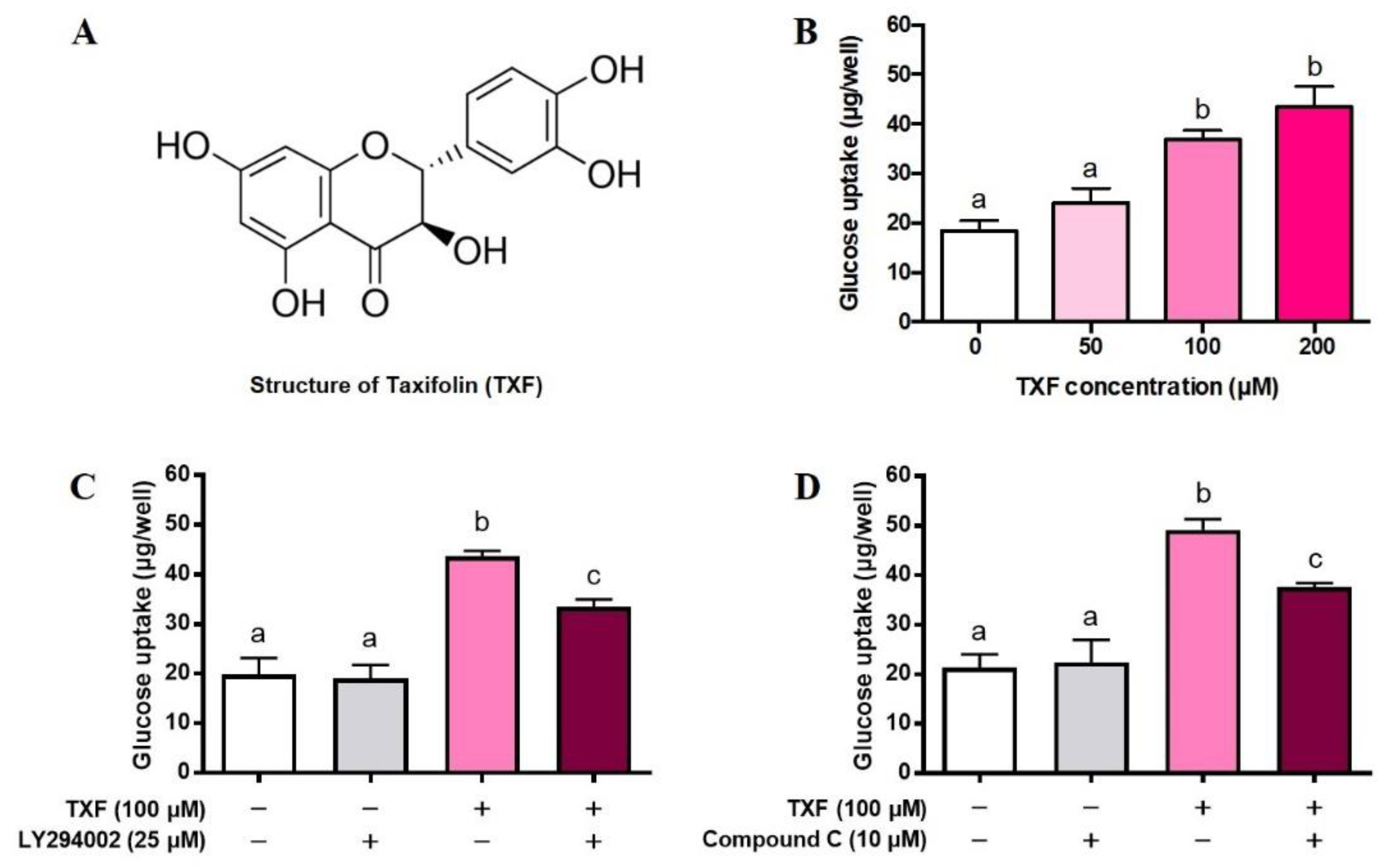
3.1. Effect of Taxifolin in Cultured Myocytes
3.1.1. Glucose Uptake in L6 Myotubes and Effect of Taxifolin
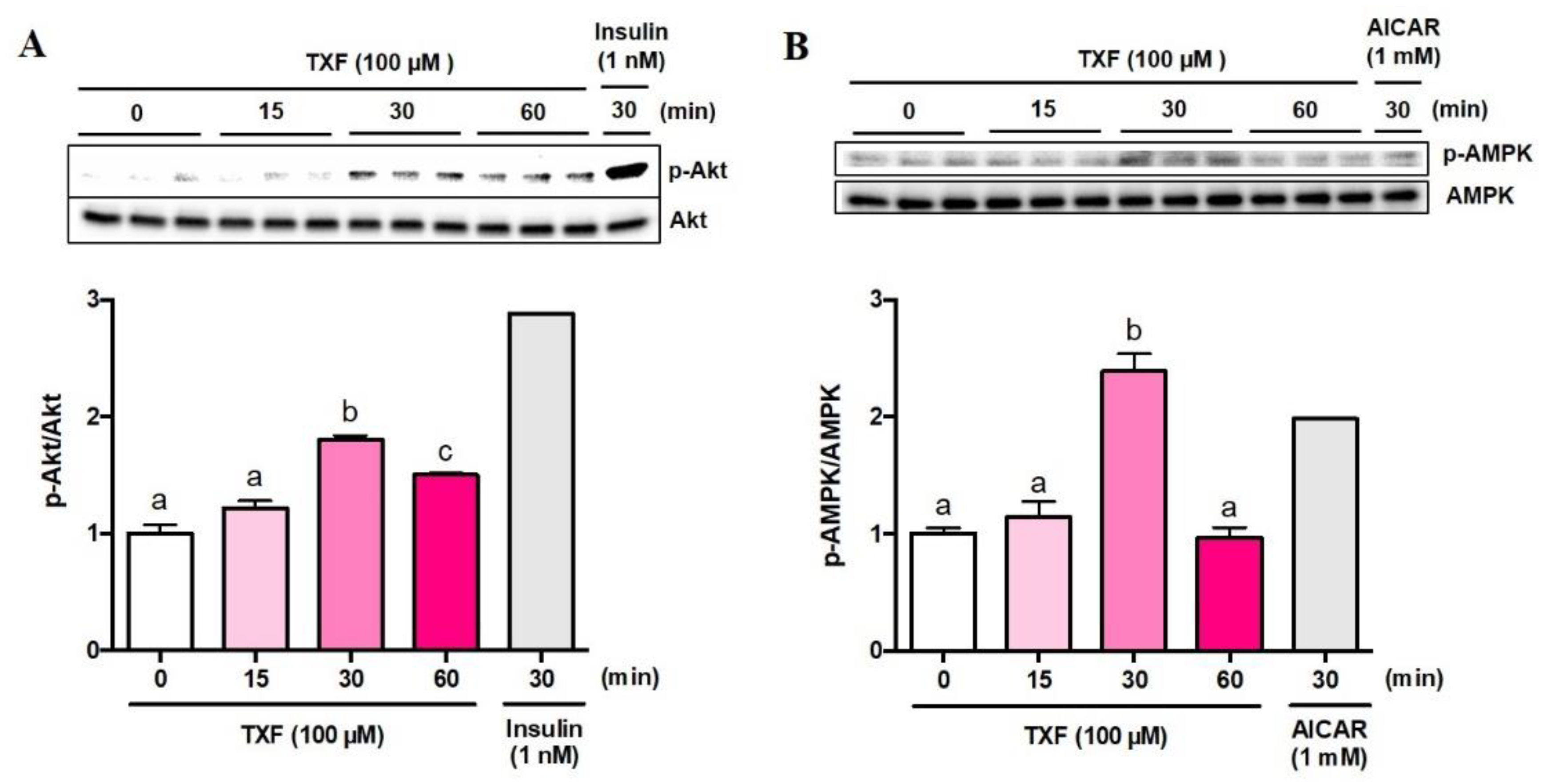
3.1.2. Phosphorylation and Activation of Akt and AMPK by Taxifolin in L6 Myotubes
3.1.3. Translocation of GLUT4 to Plasma Membrane by Taxifolin in L6 Myotubes
3.2. Effect of Taxifolin in T2D Model KK-Ay/Ta Mice
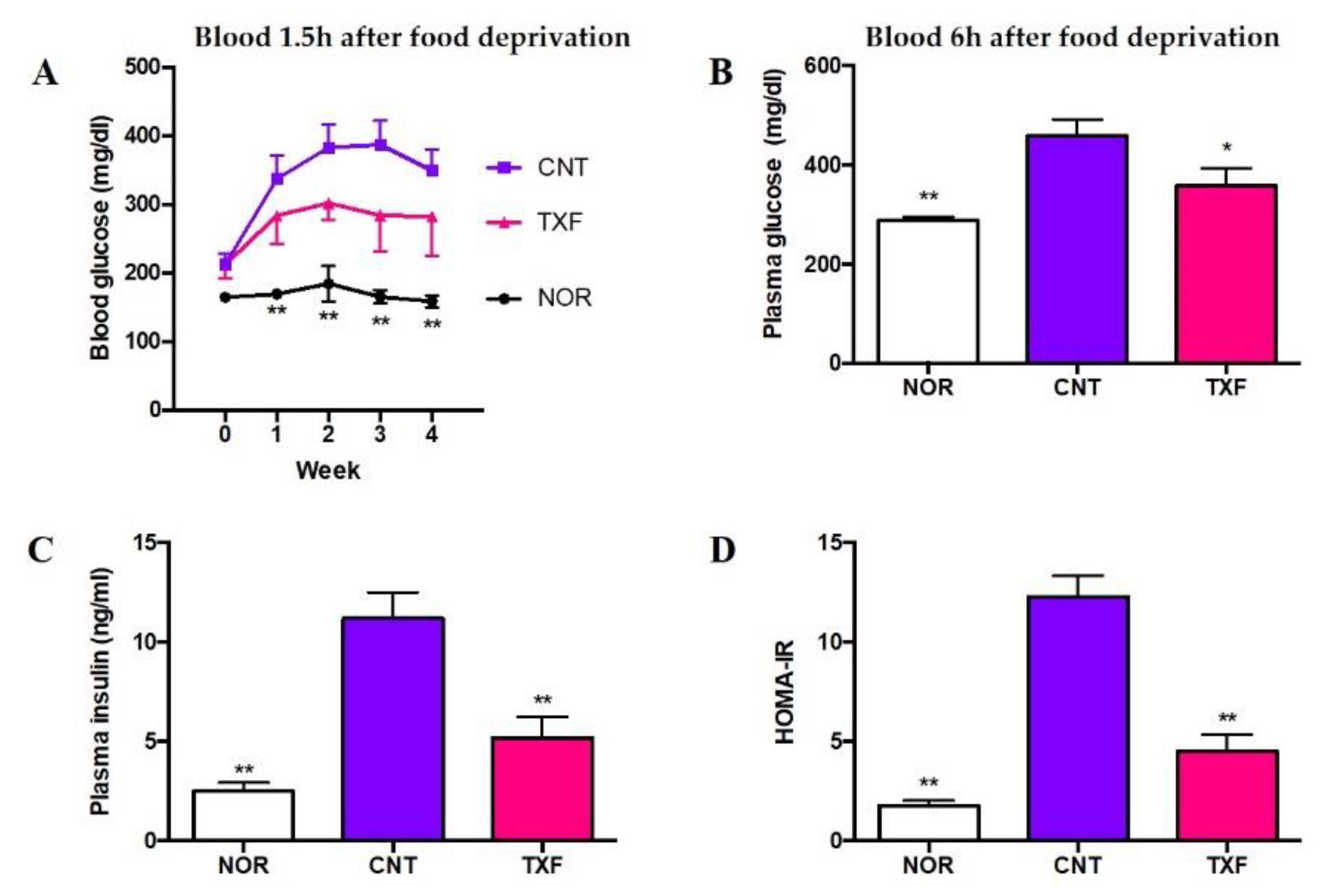
3.2.1. In Vivo Effect of Taxifolin on Food Intake and Body Weight Gain in T2D Model Mice
3.2.2. In Vivo Effect of Taxifolin on Plasma Glucose, Insulin, and HOMA-IR in T2D Model Mice
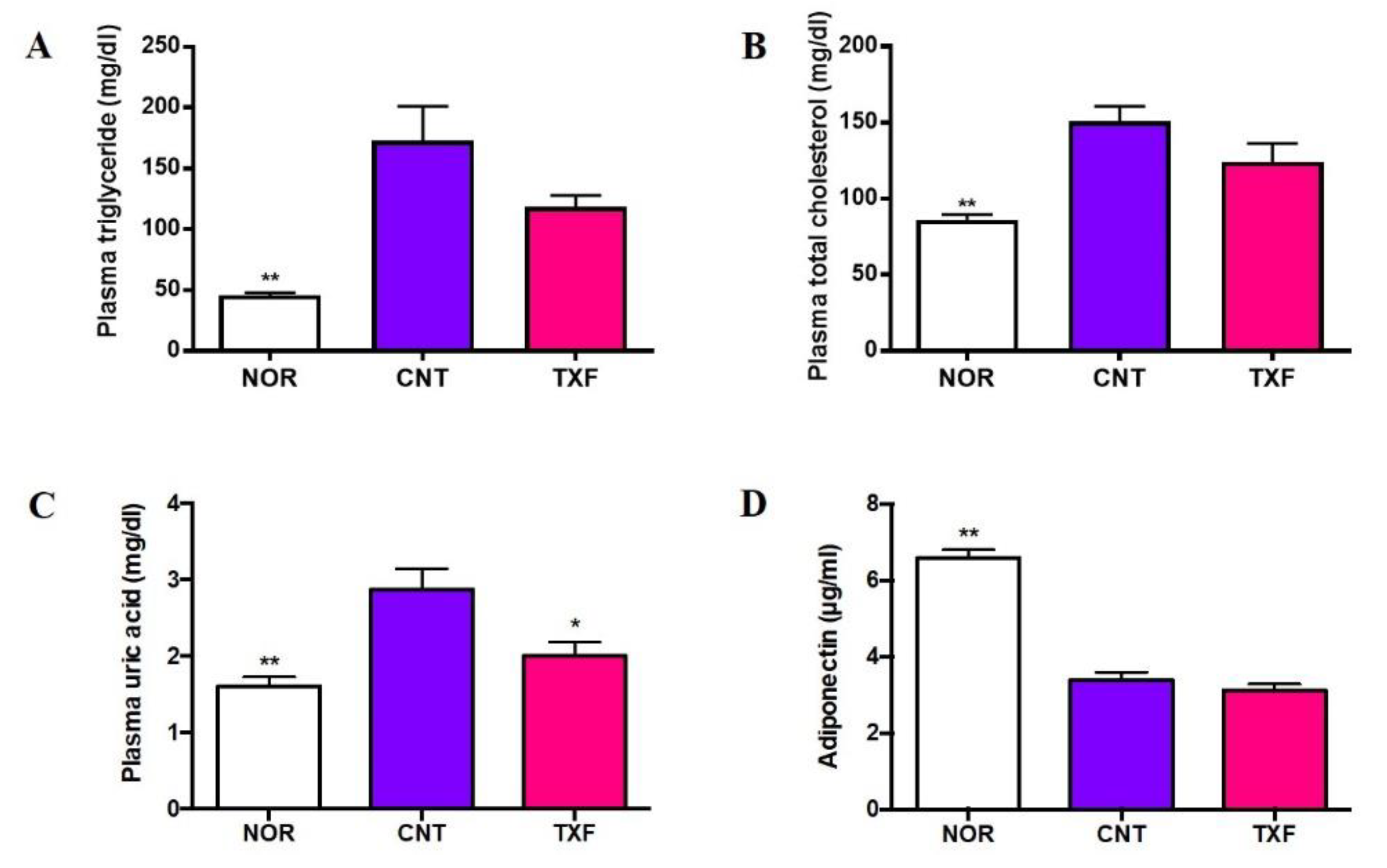
3.2.3. In Vivo Effect of Taxifolin on Plasma Lipids, Uric Acid, and Adiponectin in T2D Model Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AICAR | 5-aminoimidazole-4-carboxamide 1-β-D-ribofuranoside |
| Akt | protein kinase B |
| AMPK | 5’-adenosine monophosphate-activated protein kinase |
| BSA | bovine serum albumin |
| DTT | dithiothreitol |
| DMEM | Dulbecco’s modified Eagle medium |
| FBS | fetal bovine serum |
| GLUT4 | glucose transporter 4 |
| GU | glucose uptake |
| HOMA-IR | homeostasis model assessment of insulin resistance |
| KHH buffer | Krebs–Henseleit–HEPES buffer |
| PI3K | phosphatidylinositol 3-kinase |
| TBST | Tris buffered saline with Tween 20 |
| T2D | type 2 diabetes |
| TXF | taxifolin |
| UA | uric acid |
References
- International Diabetes Federation. IDF Diabetes Atlas, 9th ed.; International Diabetes Federation: Brussels, Belgium, 2019; Available online: https://www.diabetesatlas.org (accessed on 3 April 2021).
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Saltiel, A.R.; Kahan, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.C.; Yoshizawa, F.; Anthony, T.G.; Vary, T.C.; Jefferson, L.S.; Kimball, S.R. Leucine stimulates translation initiation in skeletal muscle of postabsorptive rats via a rapamycin-sensitive pathway. J. Nutr. 2000, 130, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, D. Retention of differentiation potentialities during prolonged cultivation of myogenic cells. Proc. Natl. Acad. Sci. USA 1968, 61, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagasaki, K.; Morisaki, N.; Kitahara, Y.; Miura, A.; Funabiki, R. Involvement of protein kinase C activation in L-leucine-induced stimulation of protein synthesis in L6 myotubes. Cytotechnology 2003, 43, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Miura, Y.; Nakazawa, T.; Yagasaki, K. Possible involvement of calcium signaling pathways in L-leucine-stimulated protein synthesis in L6 myotubes. Biosci. Biotechnol. Biochem. 2006, 70, 1533–1536. [Google Scholar] [CrossRef] [Green Version]
- Nishitani, S.; Matsumura, T.; Fujitani, S.; Sonaka, I.; Miura, Y.; Yagasaki, K. Leucine promotes glucose uptake in skeletal muscles of rats. Biochem. Biophys. Res. Commun. 2002, 299, 693–696. [Google Scholar] [CrossRef]
- Miura, Y.; Shiomi, H.; Sakai, F.; Yagasaki, K. Assay systems for screening food components that have anti-proliferative and anti-invasive activity to rat ascites hepatoma cells: In vitro and ex vivo effects of green tea extract. Cytotechnology 1997, 23, 127–132. [Google Scholar] [CrossRef]
- Kawano, A.; Nakamura, H.; Hata, S.; Minakawa, M.; Miura, Y.; Yagasaki, K. Hypoglycemic effect of aspalathin, a rooibos tea component from Aspalathus linearis, in type 2 diabetic model db/db mice. Phytomedicine 2009, 16, 437–443. [Google Scholar] [CrossRef]
- Son, M.J.; Minakawa, M.; Miura, Y.; Yagasaki, K. Aspalathin improves hyperglycemia and glucose intolerance in obese diabetic ob/ob mice. Eur. J. Nutr. 2013, 52, 1607–1619. [Google Scholar] [CrossRef]
- Yagasaki, K. Anti-diabetic phytochemicals that promote GLUT4 translocation via AMPK signaling in muscle cells. Nutr. Aging 2014, 2, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Muller, C.J.F.; Malherbe, C.J.; Chellan, N.; Yagasaki, K.; Miura, Y.; Joubert, E. Potential of rooibos, its major C-glucosyl flavonoids, and Z-2-(β-D-glucopyranosyloxy)-3-phenylpropenoic acid in prevention of metabolic syndrome. Crit. Rev. Food Sci. Nutr. 2018, 58, 227–246. [Google Scholar] [CrossRef]
- Yagasaki, K. Chapter 26: Phytochemicals, Their Intestinal Metabolites, and Skeletal Muscle Function; In Nutrition and Skeletal Muscle, 1st ed.; Walrand, S., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 421–438. [Google Scholar] [CrossRef]
- Sunil, C.; Xu, B. An insight into the health-promoting effects of taxifolin (dihydroquercetin). Phytochemistry 2019, 166, 112066. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, X.; Yang, T.; Solvin, J.; Chen, P. Profiling polyphenols of two diploid strawberry (Fragaria vesca) inbred lines using UHPLC-HRMS(n.). Food Chem. 2014, 146, 289–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Saito, S.; Inoue, T.; Satoh-Asahara, N.; Ihara, M. Novel Therapeutic Potentials of Taxifolin for Amyloid-β-associated Neurodegenerative Diseases and Other Diseases: Recent Advances and Future Perspectives. Int. J. Mol. Sci. 2019, 20, 2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Hu, Y.; Huang, T.; Zhang, Y.; Li, Z.; Luo, C.; Luo, Y.; Yuan, H.; Hisatome, I.; Yamamoto, T.; et al. High uric acid directly inhibits insulin signalling and induces insulin resistance. Biochem. Biophys. Res. Commun. 2014, 447, 707–714. [Google Scholar] [CrossRef]
- Adachi, S.I.; Yoshizawa, F.; Yagasaki, K. Hyperuricemia in type 2 diabetic model KK-Ay/Ta mice: A potent animal model with positive correlation between insulin resistance and plasma high uric acid levels. BMC Res. Notes 2017, 10, 577. [Google Scholar] [CrossRef] [Green Version]
- Adachi, S.I.; Nihei, K.I.; Ishihara, Y.; Yoshizawa, F.; Yagasaki, K. Anti-hyperuricemic effect of taxifolin in cultured hepatocytes and model mice. Cytotechnology 2017, 69, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Minakawa, M.; Kawano, A.; Miura, Y.; Yagasaki, K. Hypoglycemic effect of resveratrol in type 2 diabetic model db/db mice and its actions in cultured L6 myotubes and RIN-5F pancreatic β-cells. J. Clin. Biochem. Nutr. 2011, 48, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Ha, B.G.; Nagaoka, M.; Yonezawa, T.; Tanabe, R.; Woo, J.T.; Kato, H.; Chung, U.I.; Yagasaki, K. Regulatory mechanism for the stimulatory action of genistein on glucose uptake in vitro and in vivo. J. Nutr. Biochem. 2012, 23, 501–509. [Google Scholar] [CrossRef]
- Ha, B.G.; Yonezawa, T.; Son, M.J.; Woo, J.T.; Ohba, S.; Chung, U.I.; Yagasaki, K. Antidiabetic effect of nepodin, a component of Rumex roots, and its modes of action in vitro and in vivo. Biofactors 2014, 40, 436–447. [Google Scholar] [CrossRef]
- Nishiumi, S.; Ashida, H. Rapid preparation of a plasma membrane fraction from adipocytes and muscle cells: Application to detection of translocated glucose transporter 4 on the plasma membrane. Biosci. Biotechnol. Biochem. 2007, 71, 2343–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Cha, B.Y.; Saito, K.; Yamakawa, H.; Choi, S.S.; Yamaguchi, K.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.T. Nobiletin improves hyperglycemia and insulin resistance in obese diabetic ob/ob mice. Biochem. Pharmacol. 2010, 79, 1674–1683. [Google Scholar] [CrossRef] [PubMed]
- Cheong, S.H.; Furuhashi, K.; Ito, K.; Nagaoka, M.; Yonezawa, T.; Miura, Y.; Yagasaki, K. Antihyperglycemic effect of equol, a daidzein derivative, in cultured L6 myocytes and ob/ob mice. Mol. Nutr. Food Res. 2014, 58, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Furuhashi, K.; Son, M.J.; Toyozaki, M.; Yoshizawa, F.; Miura, Y.; Yagasaki, K. Antidiabetic effect of enterolactone in cultured muscle cells and in type 2 diabetic model db/db mice. Cytotechnology 2017, 69, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Minakawa, M.; Miura, Y.; Yagasaki, K. Piceatannol, a resveratrol derivative, promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and suppresses blood glucose levels in type 2 diabetic model db/db mice. Biochem. Biophys. Res. Commun. 2012, 422, 469–475. [Google Scholar] [CrossRef]
- Cheong, S.H.; Furuhashi, K.; Ito, K.; Nagaoka, M.; Yonezawa, T.; Miura, Y.; Yagasaki, K. Daidzein promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and improves glucose homeostasis in Type 2 diabetic model mice. J. Nutr. Biochem. 2014, 25, 136–143. [Google Scholar] [CrossRef]
- Rehman, K.; Chohan, T.A.; Waheed, I.; Gilani, Z.; Akash, M.S.H. Taxifolin prevents postprandial hyperglycemia by regulating the activity of α-amylase: Evidence from an in vivo and in silico studies. J. Cell Biochem. 2019, 120, 425–438. [Google Scholar] [CrossRef] [Green Version]
- Yoon, K.D.; Lee, J.Y.; Kim, T.Y.; Kang, H.; Ha, K.S.; Ham, T.H.; Ryu, S.N.; Kang, M.Y.; Kim, Y.H.; Kwon, Y.I. In Vitro and in Vivo Anti-Hyper glycemic Activities of Taxifolin and Its Derivatives Isolated from Pigmented Rice (Oryzae sativa L. cv. Superhongmi). J. Agric. Food Chem. 2020, 68, 742–750. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, P.; Zhang, Q.; Fu, Y.; Hou, Y.; Wei, Y.; Zheng, X.; Feng, W. Taxifolin improves disorders of glucose metabolism and water-salt metabolism in kidney via PI3K/AKT signaling pathway in metabolic syndrome rats. Life Sci. 2020, 263, 118713. [Google Scholar] [CrossRef] [PubMed]
- Tomino, Y. Lessons From the KK-Ay Mouse, a Spontaneous Animal Model for the Treatment of Human Type 2 Diabetic Nephropathy. Nephrourol. Mon. 2012, 4, 524–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Tanimoto, M.; Yamada, K.; Kaneko, S.; Matsumoto, M.; Obayashi, K.; Hagiwara, S.; Murakoshi, M.; Aoki, T.; Wakabayashi, M.; et al. Glomerular changes in the KK-Ay/Ta mouse: A possible model for human type 2 diabetic nephropathy. Nephrology (Carlton) 2006, 11, 29–35. [Google Scholar] [CrossRef]
- Ishikawa, T.; Aw, W.; Kaneko, K. Metabolic Interactions of Purine Derivatives with Human ABC Transporter ABCG2: Genetic Testing to Assess Gout Risk. Pharmaceuticals 2013, 6, 1347–1360. [Google Scholar] [CrossRef]
- Zhao, Y.; Hu, X.; Liu, Y.; Dong, S.; Wen, Z.; He, W.; Zhang, S.; Huang, Q.; Shi, M. ROS signaling under metabolic stress: Cross-talk between AMPK and AKT pathway. Mol. Cancer 2017, 16, 79. [Google Scholar] [CrossRef] [Green Version]
- Louden, E.; Chi, M.M.; Moley, K.H. Crosstalk between the AMP-activated kinase and insulin signaling pathways rescues murine blastocyst cells from insulin resistance. Reproduction 2008, 136, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Lian, X.; Hong, L.; Zou, J.; Li, Z.; Zhu, Y.; Huang, T.; Zhang, Y.; Hu, Y.; Yuan, H.; et al. High Uric Acid Activates the ROS-AMPK Pathway, Impairs CD68 Expression and Inhibits OxLDL-Induced Foam-Cell Formation in a Human Monocytic Cell Line, THP-1. Cell Physiol. Biochem. 2016, 40, 538–548. [Google Scholar] [CrossRef]
- Kimura, Y.; Yanagida, T.; Onda, A.; Tsukui, D.; Hosoyamada, M.; Kono, H. Soluble Uric Acid Promotes Atherosclerosis via AMPK (AMP-Activated Protein Kinase)-Mediated Inflammation. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 570–582. [Google Scholar] [CrossRef]
- Zhanataev, A.K.; Kulakova, A.V.; Nasonova, V.V.; Durnev, A.D. In vivo study of dihydroquercetin genotoxicity. Bull. Exp. Biol. Med. 2008, 145, 338–340. [Google Scholar] [CrossRef]
- Makena, P.S.; Pierce, S.C.; Chung, K.T.; Sinclair, S.E. Comparative mutagenic effects of structurally similar flavonoids quercetin and taxifolin on tester strains Salmonella typhimurium TA102 and Escherichia coli WP-2 uvrA. Environ. Mol. Mutagen. 2009, 50, 451–459. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement | NOR | CNT | TXF |
|---|---|---|---|
| Initial body weight (g) | 19.4 ± 0.2 ** | 28.4 ± 0.5 | 28.7 ± 0.8 |
| Final body weight (g) | 23.1 ± 0.3 ** | 37.6 ± 0.9 | 36.1 ± 1.3 |
| Weight gain (g/4 weeks) | 3.7 ± 0.2 ** | 9.2 ± 0.6 | 7.4 ± 1.0 |
| Food intake (g/4 weeks) | 89.1 ± 1.3 ** | 162.9 ± 6.0 | 154.9 ± 9.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondo, S.; Adachi, S.-i.; Yoshizawa, F.; Yagasaki, K. Antidiabetic Effect of Taxifolin in Cultured L6 Myotubes and Type 2 Diabetic Model KK-Ay/Ta Mice with Hyperglycemia and Hyperuricemia. Curr. Issues Mol. Biol. 2021, 43, 1293-1306. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030092
Kondo S, Adachi S-i, Yoshizawa F, Yagasaki K. Antidiabetic Effect of Taxifolin in Cultured L6 Myotubes and Type 2 Diabetic Model KK-Ay/Ta Mice with Hyperglycemia and Hyperuricemia. Current Issues in Molecular Biology. 2021; 43(3):1293-1306. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030092
Chicago/Turabian StyleKondo, Shinji, Shin-ichi Adachi, Fumiaki Yoshizawa, and Kazumi Yagasaki. 2021. "Antidiabetic Effect of Taxifolin in Cultured L6 Myotubes and Type 2 Diabetic Model KK-Ay/Ta Mice with Hyperglycemia and Hyperuricemia" Current Issues in Molecular Biology 43, no. 3: 1293-1306. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030092