
Potential DPP IV Inhibitory Peptides from Dry-Cured Pork Loins after Hydrolysis: An In Vitro and In Silico Study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Dry-Cured Loin Preparation
2.2. Muscle Protein Extraction and Hydrolysis
2.3. Peptide Fractions and Spectrometric Identification
2.4. Peptide Characterization and DPP IV Inhibitory Activity Determination—Chemometric Analysis
2.4.1. In Silico Bioactivity Prediction
2.4.2. In Silico Peptide–Protein Interaction Prediction
2.4.3. SwissTargetPrediction
2.4.4. ADMET Prediction
3. Results and Discussion
3.1. Peptide Presentation Based on Spectrometric Analysis
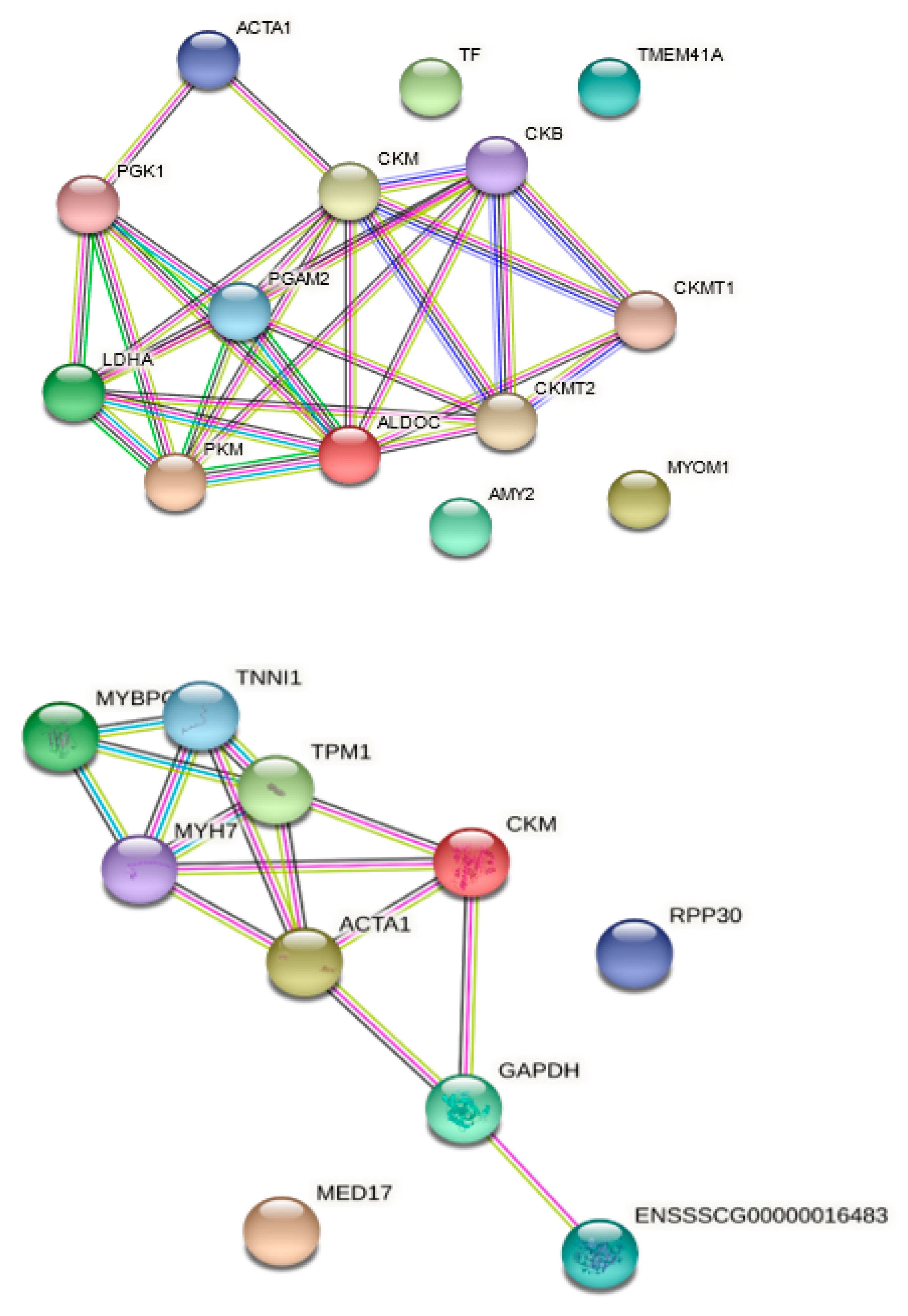
3.2. Analysis of the Protein Functional Interaction Networks
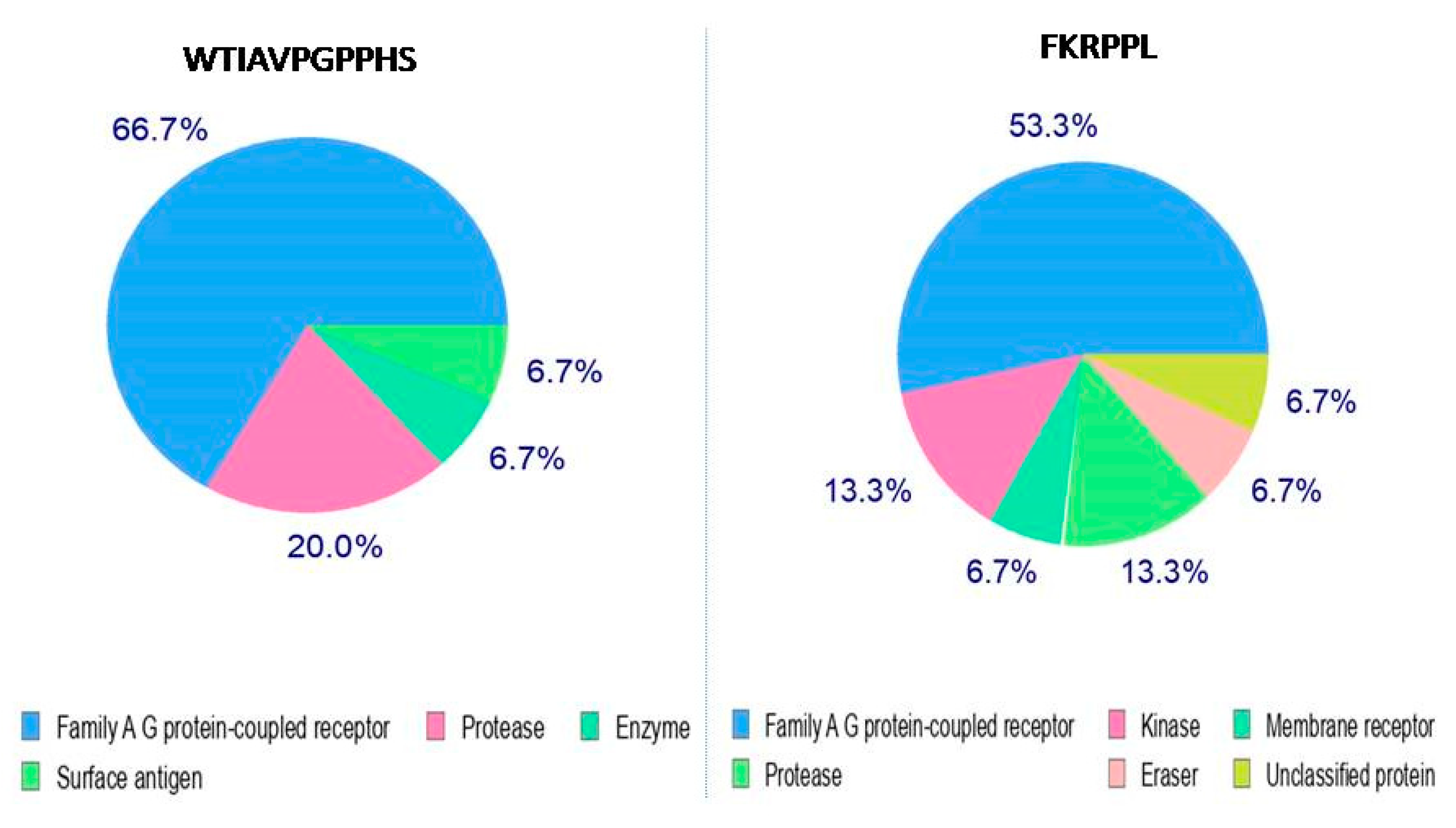
3.3. Peptides–Enzymes Interaction Prediction
3.4. ADMET Prediction
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mora, L.; Escudero, E.; Toldrá, F. Characterization of the peptide profile in Spanish Teruel, Italian Parma and Belgian dry-cured hams and its potential bioactivity. Food Res. Int. 2016, 89, 638–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogorzelska-Nowicka, E.; Atanasov, A.G.; Horbańczuk, J.; Wierzbicka, A. Bioactive compounds in functional meat products. Molecules 2018, 23, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudero, E.; Mora, L.; Toldrá, F. Stability of ACE inhibitory ham peptides against heat treatment and in vitro digestion. Food Chem. 2014, 161, 305–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kęska, P.; Stadnik, J. Stability of antiradical activity of protein extracts and hydrolysates from dry-cured pork loins with probiotic strains of LAB. Nutrients 2018, 10, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kęska, P.; Stadnik, J. Ageing-time dependent changes of angiotensin I-converting enzyme-inhibiting activity of protein hydrolysates obtained from dry-cured pork loins inoculated with probiotic lactic acid bacteria. Int. J. Pept. Res. Ther. 2019, 25, 1173–1185. [Google Scholar] [CrossRef] [Green Version]
- Gallego, M.; Mauri, L.; Aristoy, M.C.; Toldrá, F.; Mora, L. Antioxidant peptides profile in dry-cured ham as affected by gastrointestinal digestion. J. Funct. Foods 2020, 69, 103956. [Google Scholar] [CrossRef]
- Gallego, M.; Aristoy, M.C.; Toldrá, F. Dipeptidyl peptidase IV inhibitory peptides generated in Spanish dry-cured ham. Meat Sci. 2014, 96, 757–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nongonierma, A.B.; FitzGerald, R.J. Features of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from dietary proteins. J Food Biochem. 2019, 43, e12451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. 2015, 20, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Lau, J.L.; Dunn, M.K. Therapeutic peptides: Historical perspectives, current development trends, and future directions. Bioorg. Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef] [PubMed]
- FDA. Direct food substances affirmed as generally recognized as safe. Code Fed. Regul. 2003, 21, 184. [Google Scholar]
- Schaafsma, G. Safety of protein hydrolysates, fractions thereof and bioactive peptides in human nutrition. Eur. J. Clin. Nutr. 2009, 63, 1161–1168. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Yu, Z.; Zhao, W.; Ding, L.; Zheng, F.; Li, J.; Liu, J. Identification and molecular mechanism of angiotensin-converting enzyme inhibitory peptides from Larimichthyscrocea titin. Food Sci. Hum. Wellness 2020, 9, 257–263. [Google Scholar] [CrossRef]
- Molina, I.; Toldrá, F. Detection of proteolytic activity in microorganisms isolated from dry-cured ham. J. Food Sci. 1992, 57, 1308–1310. [Google Scholar] [CrossRef]
- Fadda, S.; Sanz, Y.; Vignolo, G.; Aristoy, M.C.; Oliver, G.; Toldrá, F. Characterization of muscle sarcoplasmic and myofibrillar protein hydrolysis caused by Lactobacillus plantarum. App. Environ. Microbiol. 1999, 65, 3540–3546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Escudero, E.; Sentandreu, M.A.; Toldra, F. Characterization of peptides released by in vitro digestion of pork meat. J. Agric. Food Chem. 2010, 58, 5160–5165. [Google Scholar] [CrossRef]
- Kęska, P.; Rohn, S.; Halagarda, M.; Wójciak, K.M. Peptides from different carcass elements of organic and conventional pork—Potential source of antioxidant activity. Antioxidants 2020, 9, 835. [Google Scholar] [CrossRef]
- BIOPEP-UWM Database. Available online: http://www.uwm.edu.pl/biochemia/index.php/pl/biopep (accessed on 1 February 2021).
- STRING Database. Available online: https://string-db.org (accessed on 1 February 2021).
- SwissTargetPrediction Website. Available online: http://www.swisstargetprediction.ch/ (accessed on 1 February 2021).
- ADMETlab Website. Available online: http://admet.scbdd.com/ (accessed on 1 February 2021).
- Stachniuk, A.; Sumara, A.; Montowska, M.; Fornal, E. Peptide markers for distinguishing guinea fowl meat from that of other species using liquid chromatography–mass spectrometry. Food Chem. 2021, 345, 128810. [Google Scholar] [CrossRef]
- Kęska, P.; Stadnik, J.; Wójciak, K.M.; Neffe-Skocińska, K. Physico-chemical and proteolytic changes during cold storage of dry-cured pork loins with probiotic strains of LAB. Int. J. Food Sci. Technol. 2020, 55, 1069–1079. [Google Scholar] [CrossRef]
- Huang, C.; Hou, C.; Ijaz, M.; Yan, T.; Li, X.; Li, Y.; Zhang, D. Proteomics discovery of protein biomarkers linked to meat quality traits in post-mortem muscles: Current trends and future prospects: A review. Trends Food. Sci. Technol. 2020, 105, 416–432. [Google Scholar] [CrossRef]
- Sayd, T.; Chambon, C.; Santé-Lhoutellier, V. Quantification of peptides released during in vitro digestion of cooked meat. Food Chem. 2016, 197, 1311–1323. [Google Scholar] [CrossRef]
- Mora, L.; Fraser, P.D.; Toldrá, F. Proteolysis follow-up in dry-cured meat products through proteomic approaches. Food Res. Int. 2013, 54, 1292–1297. [Google Scholar] [CrossRef]
- Mora, L.; Gallego, M.; Escudero, E.; Reig, M.; Aristoy, M.C.; Toldrá, F. Small peptides hydrolysis in dry-cured meats. Int. J. Food Microbiol. 2015, 212, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilling, M.W.; Suman, S.P.; Zhang, X.; Nair, M.N.; Desai, M.A.; Cai, K.; Allen, P.J. Proteomic approach to characterize biochemistry of meat quality defects. Meat Sci. 2017, 132, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Mora, L.; Gallego, M.; Aristoy, M.C.; Toldrá, F. Evaluation of main post-translational modifications occurring in naturally generated peptides during the ripening of Spanish dry-cured ham. Food Chem. 2020, 332, 127388. [Google Scholar] [CrossRef]
- Gfeller, D.; Grosdidier, A.; Wirth, M.; Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: A web server for target prediction of bioactive small molecules. Nucleic Acids Res. 2014, 42, W32–W38. [Google Scholar] [CrossRef]
- Gallego, M.; Grootaert, C.; Mora, L.; Aristoy, M.C.; Van Camp, J.; Toldrá, F. Transepithelial transport of dry-cured ham peptides with ACE inhibitory activity through a Caco-2 cell monolayer. J. Funct. Foods 2016, 21, 388–395. [Google Scholar] [CrossRef]
- López-Barrios, L.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bioactive peptides and hydrolysates from pulses and their potential use as functional ingredients. J. Food Sci. 2014, 79, R273–R283. [Google Scholar] [CrossRef]
- Marambe, P.W.M.L.H.K.; Wanasundara, J.P.D. Seed storage proteins as sources of bioactive peptides. In Bioactive Molecules in Plant Foods; Urakapa, F.O., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2012; pp. 49–80. [Google Scholar]
- Ma, S.; McGann, M.; Enyedy, I.J. The influence of calculated physicochemical properties of compounds on their ADMET profiles. Bioorg. Med. Chem. Lett. 2021, 36, 127825. [Google Scholar] [CrossRef]
- Veligeti, R.; Madhu, R.B.; Anireddy, J.; Pasupuleti, V.R.; Avula, V.K.R.; Ethiraj, K.S.; Uppalanchi, S.; Kasturi, S.; Perumal, Y.; Zyryanov, G.V.; et al. Synthesis of novel cytotoxic tetracyclic acridone derivatives and study of their molecular docking, ADMET, QSAR, bioactivity and protein binding properties. Sci. Rep. 2020, 10, 20720. [Google Scholar] [CrossRef] [PubMed]
- Iwaniak, A.; Minkiewicz, P.; Pliszka, M.; Mogut, D.; Darewicz, M. Characteristics of biopeptides released in silico from collagens using quantitative parameters. Foods 2020, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Borawska-Dziadkiewicz, J.; Darewicz, M.; Tarczyńska, A.S. Properties of peptides released from salmon and carp via simulated human-like gastrointestinal digestion described applying quantitative parameters. PLoS ONE 2021, 16, e0255969. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Fan, Y.; Zhao, W.; Ding, L.; Li, J.; Liu, J. Novel Angiotensin-Converting Enzyme Inhibitory Peptides Derived from Oncorhynchus mykiss Nebulin: Virtual Screening and In Silico Molecular Docking Study. J. Food Sci. 2018, 83, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhang, D.; Yu, Z.; Ding, L.; Liu, J. Novel membrane peptidase inhibitory peptides with activity against angiotensin converting enzyme and dipeptidyl peptidase IV identified from hen eggs. J. Funct. Foods 2020, 64, 103649. [Google Scholar] [CrossRef]
- Çağlar, A.F.; Çakır, B.; Gülseren, İ. LC-Q-TOF/MS based identification and in silico verification of ACE-inhibitory peptides in Giresun (Turkey) hazelnut cakes. Eur. Food Res. Technol. 2021, 247, 1189–1198. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N.; Yadav, T.C.; Pruthi, V. Quantum chemical. ADMET and molecular docking studies of ferulic acid amide derivatives with a novel anticancer drug target. Med. Chem. Res. 2017, 26, 1822–1834. [Google Scholar] [CrossRef]
- Lin, S.H.; Huang, K.J.; Weng, C.F.; Shiuan, D. Exploration of natural product ingredients as inhibitors of human HMG-CoA reductase through structure-based virtual screening. Drug Des. Devel. Ther. 2015, 9, 3313. [Google Scholar]
- Majerova, P.; Hanes, J.; Olesova, D.; Sinsky, J.; Pilipcinec, E.; Kovac, A. Novel Blood–Brain Barrier Shuttle Peptides Discovered through the Phage Display Method. Molecules 2020, 25, 874. [Google Scholar] [CrossRef] [Green Version]
- Nisha, C.M.; Kumar, A.; Nair, P.; Gupta, N.; Silakari, C.; Tripathi, T.; Kumar, A. Molecular docking and in silico ADMET study reveals acylguanidine 7a as a potential inhibitor of β-secretase. Adv. Bioinform. 2016, 2016, 9258578. [Google Scholar]
- Scheife, R.T. Protein binding: What does it mean? Dicp 1989, 23, S27–S31. [Google Scholar] [CrossRef] [PubMed]
- Ghafourian, T.; Amin, Z. QSAR models for the prediction of plasma protein binding. Bioimpacts 2013, 3, 21–27. [Google Scholar] [PubMed]
- Guengerich, F.P. Cytochrome p450 and chemical toxicology. Chem. Res. Toxicol. 2008, 21, 70–83. [Google Scholar] [CrossRef]
- Dong, J.; Wang, N.N.; Yao, Z.J.; Zhang, L.; Cheng, Y.; Ouyang, D.; Lu, A.-P.; Cao, D.S. ADMETlab: A platform for systematic ADMET evaluation based on a comprehensively collected ADMET database. J. Cheminformat. 2018, 10, 29. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. | Peptide Sequences | Protein Source (UNIPROT ID) | Days | A Parameter | DPP-IV Inhibiting Peptides [Position in Sequences] | ||||
|---|---|---|---|---|---|---|---|---|---|
| 28 | 90 | 180 | 270 | 360 | |||||
| 1 | AGFAGDDAPR | Actin (P68137; ACTA1) | + | − | − | + | + | 0.5000 | FA [3–4], AP [8–9], AG [1–2] [4–5], GF [2–3] |
| 2 | AGLKPGEL | Phosphoglyceratemutase (B5KJG2; PGAM2) | + | + | + | − | − | 0.6250 | KP [4–5], GL [2–3], AG [1–2], GE [6–7], PG [5–6] |
| 3 | ALESPERPF | Phosphoglyceratekinase (F1RPH0; PGK1) | + | + | + | + | + | 0.5556 | SP [4–5], RP [7–8], AL [1–2], ES [3–4], PF [8–9] |
| 4 | DQALKPTKPM | Phosphoglyceratemutase (B5KJG2; PGAM2) | + | − | + | + | + | 0.8000 | KP [5–6] [8–9], AL [3–4], DQ [1–2], PM [9–10], PT [6–7], QA [2–3], TK [7–8] |
| 5 | ELDQALKPTKPM | Phosphoglyceratemutase (Q32KV0; PGAM2) | − | − | + | + | + | 0.6667 | KP [7–8] [10–11], AL [5–6], DQ [3–4], PM [11–12], PT [8–9], QA [4–5], TK [9–10] |
| 6 | GVDNPGHPF | Creatinekinase (Q2HYU1; CKMT2) | + | + | + | − | − | 0.8889 | HP [7–8], NP [4–5], DN [3–4], GH [6–7], GV [1–2], PF [8–9], PG [5–6], VD [2–3] |
| 7 | IFQEVIDLGGEAIK | Alpha-amylase (P00689; AMY2) | − | + | − | + | + | 0.4286 | EV [4–5], FQ [2–3], GE [10–11], GG [9–10], QE [3–4], VI [5–6] |
| 8 | IQLVEEELDRA | Tropomyosin alpha-3 chain (A1XQV4; TPM3) | + | − | − | + | + | 0.5455 | RA [10–11], DR [9–10], IQ [1–2], LV [3–4], QL [2–3], VE [4–5] |
| 9 | KDLFDPIIQDR | Creatine kinase M-type (Q5XLD3; CKM) | + | − | − | + | + | 0.5455 | DP [5–6], DR [10–11], II [7–8], IQ [8–9], PI [6–7], QD [9–10] |
| 10 | KVVDVGSKV | Pyruvate kinase PKM (P11974; PKM) | + | + | − | − | + | 0.6667 | VV [2–3], KV [1–2] [8–9], SK [7–8], VD [3–4], VG [5–6] |
| 11 | LDQALKPTKPM | Phosphoglyceratemutase (B5KJG2; PGAM2) | + | - | + | + | + | 0.7273 | KP [6–7] [9–10], AL [4–5], DQ [2–3], PM [10–11], PT [7–8], QA [3–4],TK [8–9] |
| 12 | LFDKPVSPI | Creatine kinase B-type (Q29594; CKB) | + | + | + | − | − | 0.5556 | SP [7–8], KP [4–5], PI [8–9], PV [5–6], VS [6–7] |
| 13 | LFDKPVSPL | Creatinekinase (Q2HYU1; CKMT2) | + | + | + | + | + | 0.5556 | SP [7–8], KP [4–5], PI [8–9], PV [5–6], VS [6–7] |
| 14 | MSHLGRPDGIPMPD | Phosphoglyceratekinase (F1RPH0; PGK1) | + | + | − | + | − | 0.5714 | MP [12–13], IP [10–11], RP [6–7], HL [3–4], GI [9–10], PM [11–12], SH [2–3], IPM [10–12] |
| 15 | MSHLGRPDGIPMPDK | Phosphoglyceratekinase (F1RPH0; PGK1) | + | + | + | − | + | 0.5333 | MP [12–13], IP [10–11], RP [6–7], HL [3–4], GI [9–10], PM [11–12], SH [2–3], IPM [10–12] |
| 16 | NLHPELGTDADKEH | L-lactate dehydrogenase A chain (P00339; LDHA) | + | − | − | + | + | 0.5000 | HP [3–4], AD [10–11], EH [13–14], KE [12–13], LH [2–3], NL [1–2], TD [8–9] |
| 17 | PEILPDGDHD | Fructose-bisphosphate aldolase (F1RJ25; ALDOC) | + | + | + | + | + | 0.4000 | LP [4–5], EI [2–3], HD [9–10], IL [3–4] |
| 18 | PEILPDGDHDL | Fructose-bisphosphate aldolase (F1RJ25; ALDOC) | + | + | + | + | + | 0.3636 | LP [4–5], EI [2–3], HD [9–10], IL [3–4] |
| 19 | PEILPDGDHDLK | Fructose-bisphosphate aldolase (F1RJ25; ALDOC) | + | + | + | + | + | 0.3333 | LP [4–5], EI [2–3], HD [9–10], IL [3–4] |
| 20 | SAPILNIPI | Transmembrane protein 41A (I3LPS6; TMEM41A) | + | + | + | − | − | 0.7778 | IPI [7–9], AP [2–3], IP [7–8], IL [4–5], LN [5–6], PI [3–4] [8–9] |
| 21 | SAPILNIPIV | Transmembrane protein 41A (I3LPS6; TMEM41A) | + | + | + | − | + | 0.7778 | IPI [7–9], AP [2–3], IP [7–8], IL [4–5], LN [5–6], PI [3–4] [8–9] |
| 22 | SFDIPPPPMD | Phosphoglyceratemutase (Q32KV0; PGAM2) | − | + | + | + | + | 0.7000 | PPPP [5–8], PP [5–6] [6–7] [7–8], IP [4–5], PM [8–9], SF [1–2] |
| 23 | SFDIPPPPMDEK | Phosphoglyceratemutase (Q32KV0; PGAM2) | + | − | + | + | + | 0.6667 | PPPP [5–8], PP [5–6] [6–7] [7–8], IP [4–5], EK [11–12], PM [8–9], SF [1–2] |
| 24 | SISNSAEDPFIAIH | Alpha-amylase (F1S574; AMY2) | - | + | − | + | + | 0.4286 | IA [11–12], AE [6–7], DP [8–9], IH [13–14], PF [9–10], SI [1–2] |
| 25 | VDLPAVSEK | Multifunctional fusion protein (F1SHL9; PKM) | + | − | − | + | + | 0.6667 | PA [4–5], LP [3–4], EK [8–9], AV [5–6], VD [1–2], VS [6–7] |
| 26 | VGVNLPK | Phosphoglyceratekinase (F1RPH0; PGK1) | + | + | + | + | − | 0.8571 | LP [5–6], GV [2–3], NL [4–5], PK [6–7], VG [1–2], VN [3–4] |
| 27 | VPAPVEIPVTPPTLVSGLK | Titin (Q8WZ42; TTN) | + | - | + | − | + | 0.8421 | PP [11–12], AP [3–4], PA [2–3], VP [1–2], IP [7–8], TP [10–11], GL [17–18], EI [6–7], LV [14–15], PT [12–13], PV [4–5] [8–9], TL [13–14], VE [5–6], VS [15–16], VT [9–10] |
| 28 | WILGEHGDSSVPV | L-lactate dehydrogenase A chain (P00339; LDHA) | + | + | − | − | + | 0.5385 | VP [11–12], WI [1–2], EH [5–6], GE [4–5], IL [2–3], PV [12–13], SV [10–11] |
| 29 | WTIAVPGPPH | Myomesin 1 (F1SM75; MYOM1) | + | − | + | + | + | 0.9000 | GP [7–8], PP [8–9], VP [5–6], IA [3–4]WT [1–2], AV [4–5], PG [6–7], PH [9–10], TI [2–3] |
| 30 | WTIAVPGPPHS | Myomesin 1 (F1SM75; MYOM1) | − | − | + | + | + | 0.9091 | GP [7–8], PP [8–9], VP [5–6], IA [3–4], WT [1–2], AV [4–5], HS [10–11], PG [6–7], PH [9–10], TI [2–3] |
| 31 | YDQLPEPR | Serotransferrin (P09571; TF) | − | + | + | − | + | 0.6250 | LP [4–5], EP [6–7], DQ [2–3], QL [3–4], YD [1–2] |
| No. | Peptide Sequences | Protein Source (UNIPROT ID) | Days | A Parameter | DPP-IV Inhibiting Peptides [Position in Sequences] | ||||
|---|---|---|---|---|---|---|---|---|---|
| 28 | 90 | 180 | 270 | 360 | |||||
| 1 | AGFAGDDAPR | Actin, alpha skeletal muscle (P68137; ACTA1) | + | − | + | + | + | 0.5000 | FA [3–4], AP [8–9], AG [1–2] [4–5], GF [2–3] |
| 2 | FDKPVSPL | Creatine kinase M-type (Q5XLD3; CKM) | + | + | + | − | − | 0.6250 | SP [6–7], KP [3–4], PL [7–8], PV [4–5], VS [5–6] |
| 3 | FKRPLP | Mediator of RNA polymerase II transcription subunit 17 (F1STK7; MED17) | + | + | + | − | − | 0.6667 | LP [5–6], RP [3–4], PL [4–5], KR [2–3] |
| 4 | FKRPPI | ribonuclease P/MRP subunit p30 (F1SCX5; RPP30) | + | + | + | − | − | 0.6667 | PP [4–5], RP [3–4], KR [2–3], PI [5–6] |
| 5 | FKRPPL | Troponin I (I3LB76; TTI1) | + | + | + | − | − | 0.8333 | PP [4–5], RP [3–4], PL [5–6], PRL [4–6], KR [2–3] |
| 6 | FRVPTPNVSV | Glyceraldehyde-3-phosphate dehydrogenase (P00355; GAPDH) | + | + | − | + | − | 0.8000 | VP [3–4], TP [5–6], FR [1–2], NV [7–8], PN [6–7], PT [4–5], SV [9–10], VS [8–9] |
| 7 | IIAPPER | Actin, alpha skeletal muscle (P68137; ACTA1) | + | + | + | − | − | 0.7143 | PP [4–5], AP [3–4], IA [2–3], IIAP [1–4], II [1–2] |
| 8 | KDLFDPIIQD | Creatine kinase M-type (Q5XLD3; CKM) | + | + | − | + | − | 0.5000 | DP [5–6], II [7–8], IQ [8–9], PI [6–7], QD [9–10] |
| 9 | LGEHNIDVLEGNEQFINAAK | Trypsin (P00761) | + | + | − | + | − | 0.4500 | AA [18–19], EG [10–11], EH [3–4], GE [2–3], IN [16–17], NA [17–18], NE [12–13], QF [14–15], VL [8–9] |
| 10 | LTEAPLNPK | Actin, alpha skeletal muscle (P68137; ACTA1) | + | + | − | + | − | 0.7778 | AP [4–5], NP. [7–8], PL [5–6], LN [6–7], LT [1–2], PK [8–9], TE [2–3] |
| 11 | LVIIESDLER | Tropomyosin alpha-1 chain (P42639; TPM1) | + | − | − | + | + | 0.4000 | ES [5–6], II [3–4], LV [1–2], VI [2–3] |
| 12 | PEILPDGDHDL | Fructose-bisphosphate aldolase (P09972; ALDOC) | + | + | − | + | − | 0.3636 | LP [4–5], EI [2–3], HD [9–10], IL [3–4] |
| 13 | PEILPDGDHDLK | Fructose-bisphosphate aldolase (P09972; ALDOC) | + | + | − | + | − | 0.3333 | LP [4–5], EI [2–3], HD [9–10], IL [3–4] |
| 14 | PVTIPDKPNSE | Calsequestrin-1 (P31415; CASQ) | − | + | + | + | − | 0.5455 | IP [4–5], KP [7–8], PN [8–9], PV [1–2], TI [3–4], VT [2–3] |
| 15 | PVTIPDKPNSEE | Calsequestrin-1 (P31415; CASQ) | − | + | + | + | − | 0.5000 | IP [4–5], KP [7–8], PN [8–9], PV [1–2], TI [3–4], VT [2–3] |
| 16 | RVVDVPDPPEA | Myosin binding protein C2 (F1RH19; MYBPC2) | + | + | + | - | − | 0.4545 | PP [8–9], VP [5–6], VV [2–3], DP [7–8], VD [3–4] |
| 17 | VDDLEGSLEQEK | Myosin-7 (P79293; MYH7) | + | + | − | + | − | 0.4167 | EK [11–12], SL [7–8], EG [5–6], QE [10–11], VD [1–2] |
| WTIAVPGPPHS | ||
|---|---|---|
| Sequences | SMILES Code | Probability 1 |
| GP | NCC(=O)N1[C@@]([H])(CCC1)C(=O)O | 0.1334 |
| PP | N1[C@]([H])(CCC1)C(=O)N1[C@]([H])(CCC1)C(=O)O | 0.3095 |
| VP | N[C@@]([H])(C(C)C)C(=O)N1[C@@]([H])(CCC1)C(=O)O | 0.1858 |
| IA | N[C@@]([H])([C@]([H])(CC)C)C(=O)N[C@@]([H])(C)C(=O)O | 0.0619 |
| WT | N[C@@H](Cc1c[nH]c2c1cccc2)C(=O)N[C@@]([H])([C@]([H])(O)C)C(=O)O | 0.1273 |
| AV | N[C@]([H])(C)C(=O)N[C@]([H])(C(C)C)C(=O)O | 0.1684 |
| HS | N[C@H](Cc1c[nH]cn1)C(=O)N[C@]([H])(CO)C(=O)O | 0.0000 |
| PG | N1[C@@]([H])(CCC1)C(=O)NCC(=O)O | 0.1501 |
| PH | N1[C@@]([H])(CCC1)C(=O)N[C@@H](Cc1c[nH]cn1)C(=O)O | 0.0000 |
| TI | N[C@@]([H])([C@]([H])(O)C)C(=O)N[C@@]([H])([C@]([H])(CC)C)C(=O)O | 0.0864 |
| FKRPPL | ||
|---|---|---|
| Sequences | SMILES Code | Probability 1 |
| PP | N1[C@]([H])(CCC1)C(=O)N1[C@]([H])(CCC1)C(=O)O | 0.3095 |
| RP | N[C@]([H])(CCCNC(=N)N)C(=O)N1[C@]([H])(CCC1)C(=O)O | 0.1120 |
| PL | N1[C@]([H])(CCC1)C(=O)N[C@]([H])(CC(C)C)C(=O)O | 0.4592 |
| RPL | N[C@@]([H])(CCCNC(=N)N)C(=O)N1[C@@]([H])(CCC1)C(=O)N[C@@]([H])(CC(C)C)C(=O)O | 0.1213 |
| KR | N[C@]([H])(CCCCN)C(=O)N[C@]([H])(CCCNC(=N)N)C(=O)O | 0.0000 |
| Sequences | A | D | M | E | T | ||||
|---|---|---|---|---|---|---|---|---|---|
| Caco-2 Permeability 1 | HIA 2 | PPB 3 | BBB 4 | VD 5 | Cyp450 2D6 6 | T1/2 7 | LD50 8 | ||
| WTIAVPGPPHS | GP | −5.144 | 0 | 13.60 | 0.629 | −0.554 | 0 | 0.990 | 1.904 |
| PP | −5.055 | 0 | 25.48 | 0.716 | −0.444 | 0 | 0.845 | 2.423 | |
| VP | −5.068 | 1 (0.504) | 38.18 | 0.624 | −0.448 | 0 | 0.924 | 2.423 | |
| IA | −5.522 | 1 (0.616) | 23.65 | 0.864 | −0.505 | 0 | 1.114 | 2.158 | |
| WT | −6.061 | 0 | 53.96 | 0.956 | −0.752 | 0 | 0.715 | 2.211 | |
| AV | −5.540 | 1 (0.553) | 28.01 | 0.932 | −0.596 | 0 | 1.229 | 2.058 | |
| HS | −1.721 | 0 | 13.87 | 0.909 | −0.765 | 0 | 0.55 | 2.401 | |
| PG | −5.372 | 0 | 15.65 | 0.899 | −0.042 | 0 | 0.989 | 2.020 | |
| PH | −5.863 | 0 | 20.78 | 0.910 | −0.510 | 0 | 0.588 | 2.535 | |
| TI | −5.786 | 0 | 35.24 | 0.749 | −0.545 | 0 | 1.124 | 2.371 | |
| FKRPPL | PP | −5.055 | 0 | 25.48 | 0.714 | −0.444 | 0 | 0.845 | 2.432 |
| RP | −6.200 | 0 | 25.84 | 0.251 | −0.604 | 0 | 0.573 | 2.450 | |
| PL | −5.376 | 1 (0.510) | 45.60 | 0.830 | −0.487 | 0 | 0.876 | 2.382 | |
| RPL | −6.248 | 0 | 33.58 | 0.256 | −0.840 | 1 (substrate) | 0.723 | 2.514 | |
| KR | −6.280 | 0 | 32.87 | 0.523 | −0.759 | 0 | 0.896 | 2.328 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kęska, P.; Stadnik, J. Potential DPP IV Inhibitory Peptides from Dry-Cured Pork Loins after Hydrolysis: An In Vitro and In Silico Study. Curr. Issues Mol. Biol. 2021, 43, 1335-1349. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030095
Kęska P, Stadnik J. Potential DPP IV Inhibitory Peptides from Dry-Cured Pork Loins after Hydrolysis: An In Vitro and In Silico Study. Current Issues in Molecular Biology. 2021; 43(3):1335-1349. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030095
Chicago/Turabian StyleKęska, Paulina, and Joanna Stadnik. 2021. "Potential DPP IV Inhibitory Peptides from Dry-Cured Pork Loins after Hydrolysis: An In Vitro and In Silico Study" Current Issues in Molecular Biology 43, no. 3: 1335-1349. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030095