A Systematic Review of the Genotoxicity and Antigenotoxicity of Biologically Synthesized Metallic Nanomaterials: Are Green Nanoparticles Safe Enough for Clinical Marketing?

 , , ,
, , , 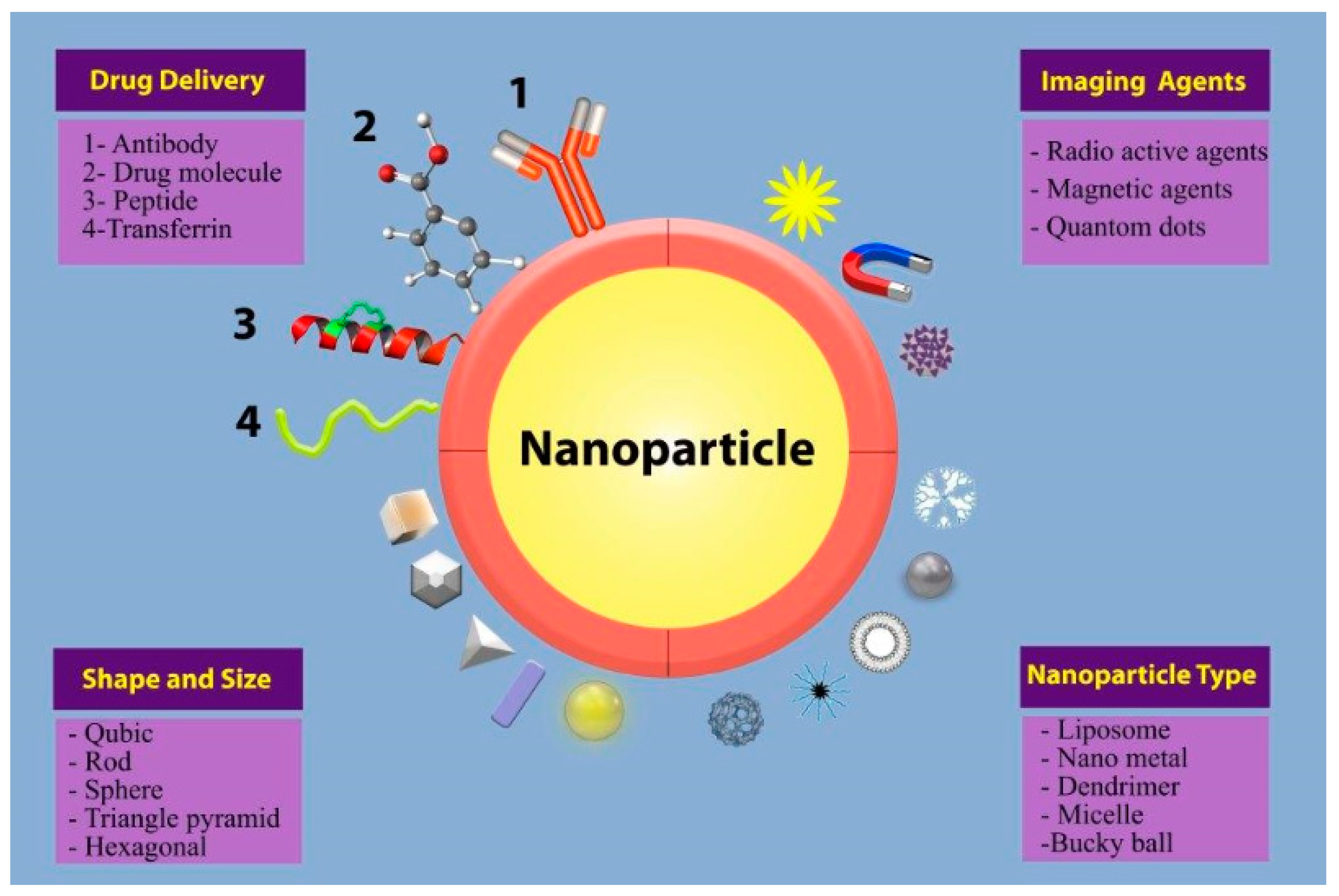{kind=link}
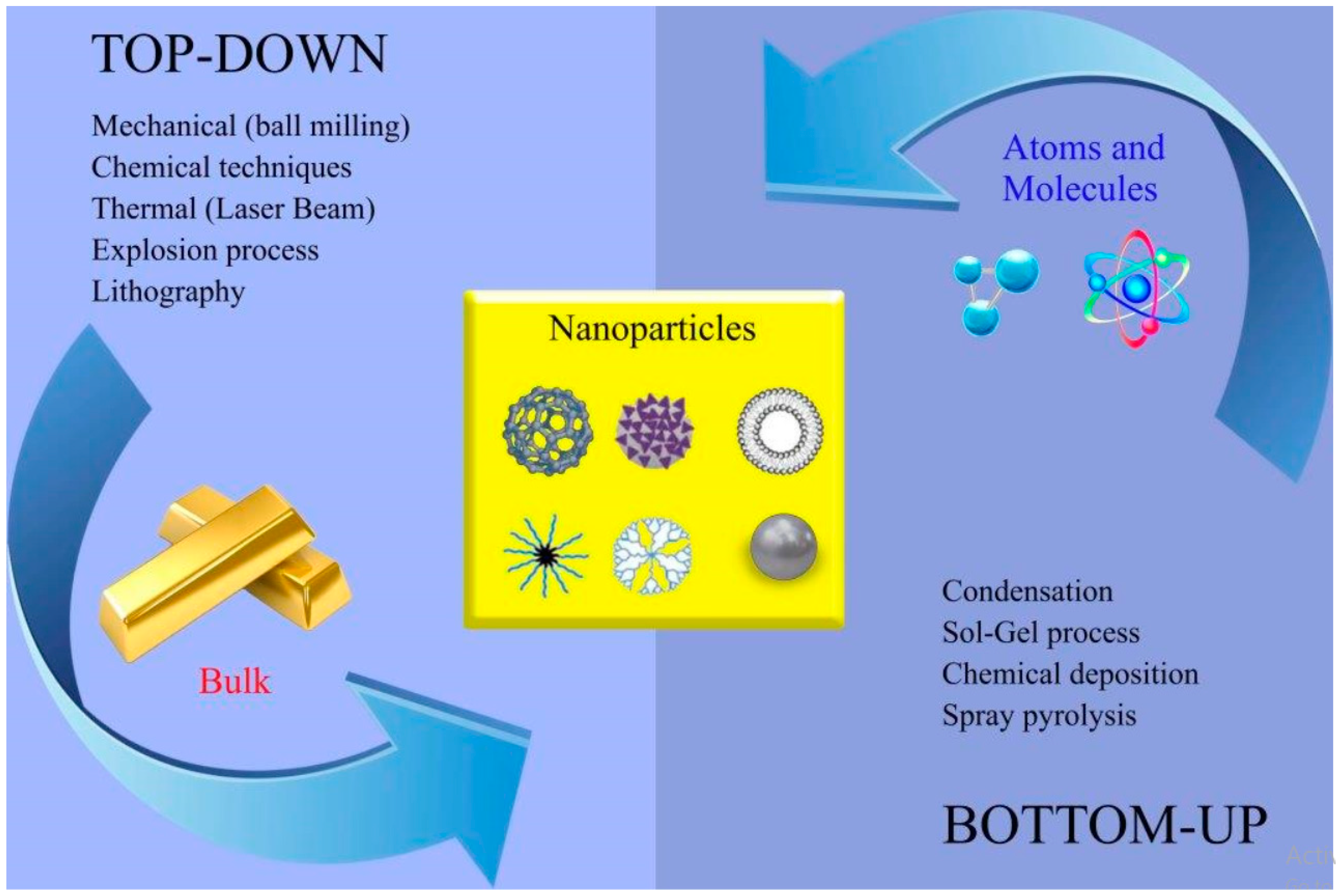{kind=link}
{kind=link}
{kind=link}
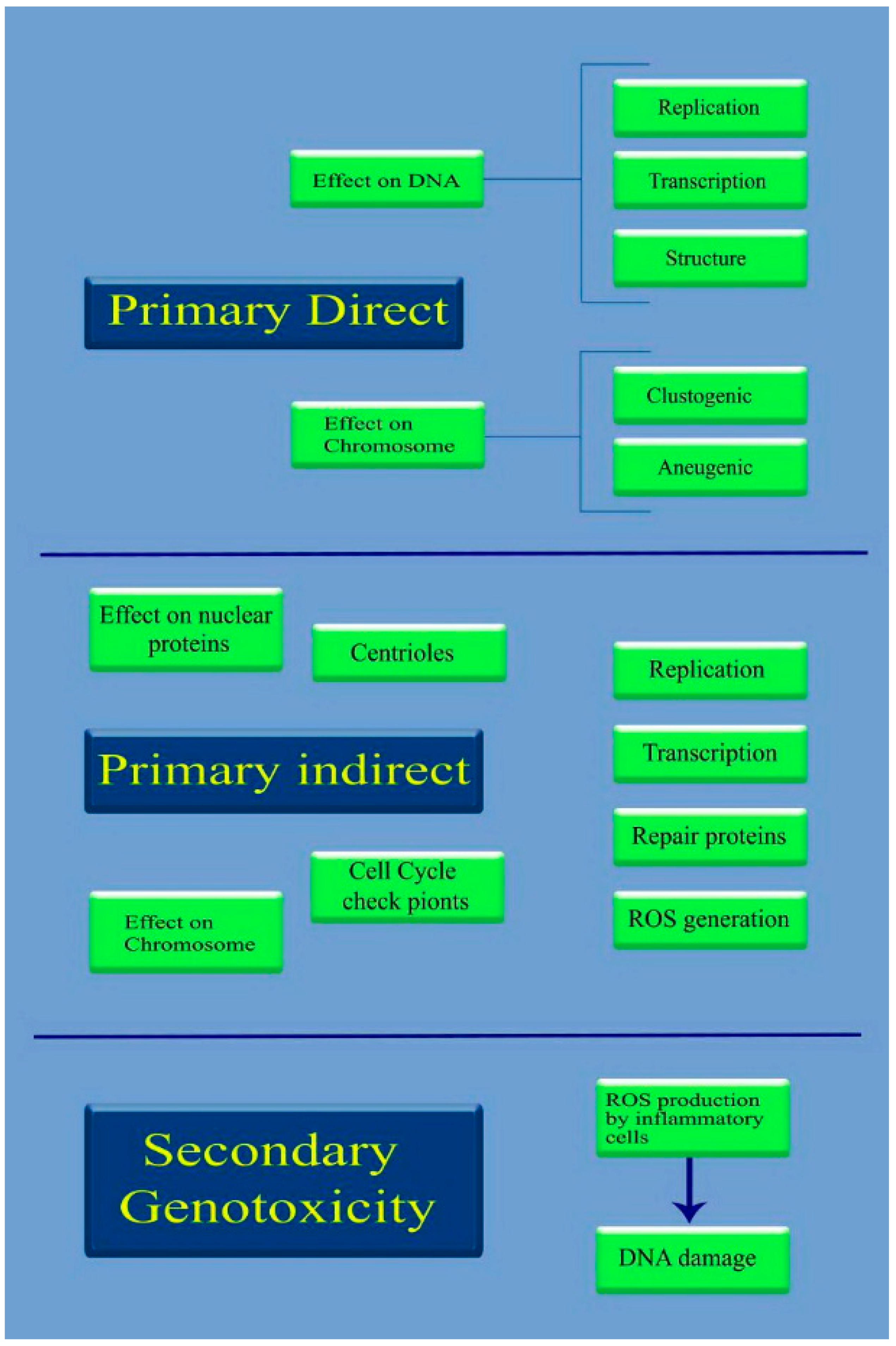{kind=link}
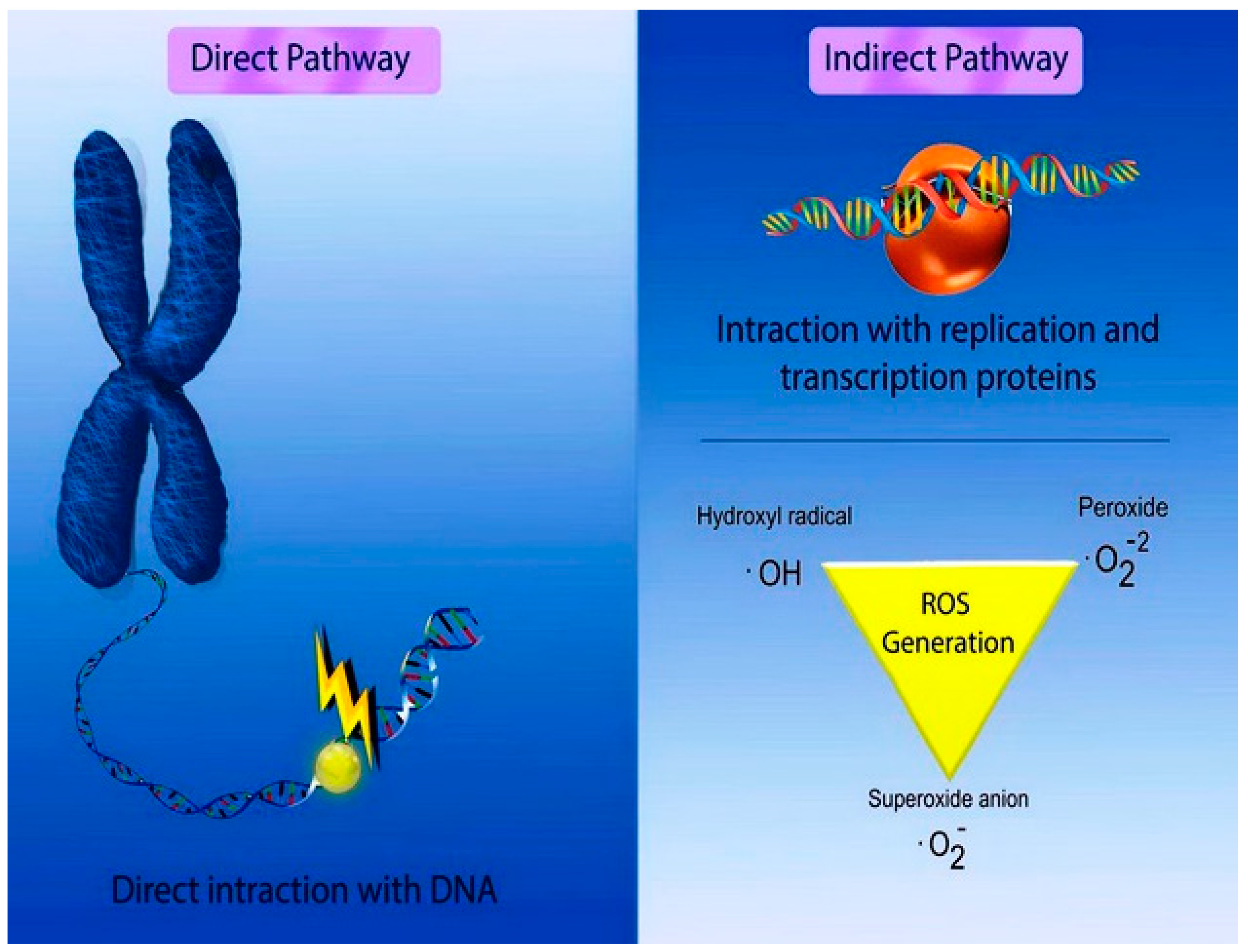{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Search Strategy
2.2. Inclusion Criteria
2.3. Exclusion Criteria
2.4. Data Collection
3. Results
4. Discussion
4.1. A Mechanistic Approach to Nanogenotoxicity
4.2. In Vitro Genotoxicity Studies of Biogenic MNPs
4.2.1. Genotoxicity Studies of Biogenic MNPs against Cancerous and Normal Cell Lines
4.2.2. Genotoxicity Studies of Biogenic MNPs against Microbes and Parasites
4.2.3. Genotoxicity Studies of Biogenic MNPs against Plants
4.3. In Vivo Genotoxicity Studies of Biogenic MNPs
4.4. Antigenotoxicity Studies of Biogenic MNPs
5. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Buzea, C.; Pacheco, I.I.; Robbie, K. Nanomaterials and nanoparticles: Sources and toxicity. Biointerphases 2007, 2, MR17–MR71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanan, M.; Asmalash, T.; Gebrekidan, A.; Gebreegziabiher, D.; Araya, T.; Hilekiros, H.; Barabadi, H.; Ramanathan, K. Nano-medicine as a newly emerging approach to combat human immunodeficiency virus (HIV). Pharm. Nanotechnol. 2018, 6, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Barabadi, H.; Mahjoub, M.A.; Tajani, B.; Ahmadi, A.; Junejo, Y.; Saravanan, M. Emerging theranostic biogenic silver nanomaterials for breast cancer: A systematic review. J. Clust. Sci. 2019, 30, 259–279. [Google Scholar] [CrossRef]
- Ovais, M.; Khalil, A.T.; Raza, A.; Islam, N.U.; Ayaz, M.; Saravanan, M.; Ali, M.; Ahmad, I.; Shahid, M.; Shinwari, Z.K. Multifunctional theranostic applications of biocompatible green-synthesized colloidal nanoparticles. Appl. Microbiol. Biotechnol. 2018, 102, 4393–4408. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xia, Y. Bottom-up and top-down approaches to the synthesis of monodispersed spherical colloids of low melting-point metals. Nano Lett. 2004, 4, 2047–2050. [Google Scholar] [CrossRef]
- Barabadi, H.; Honary, S.; Ebrahimi, P.; Alizadeh, A.; Naghibi, F.; Saravanan, M. Optimization of myco-synthesized silver nanoparticles by response surface methodology employing Box-Behnken design. Inorg. Nano Met. Chem. 2019, 49, 33–43. [Google Scholar] [CrossRef]
- Barabadi, H. Nanobiotechnology: A promising scope of gold biotechnology. Cell. Mol. Biol. 2017, 63, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Barabadi, H.; Kobarfard, F.; Vahidi, H. Biosynthesis and characterization of biogenic tellurium nanoparticles by using Penicillium chrysogenum PTCC 5031: A novel approach in gold biotechnology. Iran. J. Pharm. Res. 2018, 17, 87–97. [Google Scholar] [PubMed]
- Salari, S.; Esmaeilzadeh Bahabadi, S.; Samzadeh-Kermani, A.; Yousefzaei, F. In vitro evaluation of antioxidant and antibacterial potential of green synthesized silver nanoparticles using Prosopis farcta fruit extract. Iran. J. Pharm. Res. 2019, 18, 430–445. [Google Scholar] [PubMed]
- Dobrucka, R. Synthesis of titanium dioxide nanoparticles using Echinacea purpurea herba. Iran. J. Pharm. Res. 2017, 16, 753–759. [Google Scholar]
- Karimi, N.; Chardoli, A.; Fattahi, A. Biosynthesis, characterization, antimicrobial and cytotoxic effects of silver nanoparticles using Nigella arvensis seed extract. Iran. J. Pharm. Res. 2017, 16, 1167–1175. [Google Scholar]
- Dahoumane, S.A.; Mechouet, M.; Wijesekera, K.; Filipe, C.D.M.; Sicard, C.; Bazylinski, D.A.; Jeffryes, C. Algae-mediated biosynthesis of inorganic nanomaterials as a promising route in nanobiotechnology—A review. Green Chem. 2017, 19, 552–587. [Google Scholar] [CrossRef]
- El-Rafie, H.M.; El-Rafie, M.H.; Zahran, M.K. Green synthesis of silver nanoparticles using polysaccharides extracted from marine macro algae. Carbohydr. Polym. 2013, 96, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Honary, S.; Barabadi, H.; Ebrahimi, P.; Naghibi, F.; Alizadeh, A. Development and optimization of biometal nanoparticles by using mathematical methodology: A microbial approach. J. Nano Res. 2015, 30, 106–115. [Google Scholar] [CrossRef]
- Rezvani Amin, Z.; Khashyarmanesh, Z.; Fazly Bazzaz, B.S.; Sabeti Noghabi, Z. Does biosynthetic silver nanoparticles are more stable with lower toxicity than their synthetic counterparts? Iran. J. Pharm. Res. 2019, 18, 210–221. [Google Scholar] [PubMed]
- Barabadi, H.; Alizadeh, A.; Ovais, M.; Ahmadi, A.; Shinwari, Z.K.; Saravanan, M. Efficacy of green nanoparticles against cancerous and normal cell lines: A systematic review and meta-analysis. IET Nanobiotechnol. 2018, 12, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Mason, M.M.; Wise, J.P. Genotoxicity of metal nanoparticles. Rev. Environ. Health 2011, 26, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Shah, S. Importance of Genotoxicity & S2A guidelines for genotoxicity testing for pharmaceuticals. IOSR J. Pharm. Biol. Sci. 2012, 1, 43–54. [Google Scholar] [CrossRef]
- Seukep, A.J.; Noumedem, J.A.K.; Djeussi, D.E.; Kuete, V. 9—Genotoxicity and Teratogenicity of African Medicinal Plants. In Toxicological Survey of African Medicinal Plants; Kuete, V., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 235–275. [Google Scholar]
- Mohamed, S.; Sabita, U.; Rajendra, S.; Raman, D. Genotoxicity: Mechanisms, Testing Guidelines and Methods. Glob. J. Pharm. Pharm. Sci. 2017, 1, 1–6. [Google Scholar] [CrossRef]
- Rama Rao, P.; Kaul, C.; Jena, G. Genotoxicity testing, a regulatory requirement for drug discovery and development: Impact of ICH guidelines. Indian J. Pharmacol. 2002, 34, 86–89. [Google Scholar]
- Amoabediny, G.; Naderi, A.; Malakootikhah, J.; Koohi, M.K.; Mortazavi, S.A.; Naderi, M.; Rashedi, H. Guidelines for safe handling, use and disposal of nanoparticles. J. Phys. Conf. Ser. 2009, 170, 012037. [Google Scholar] [CrossRef]
- Rasmussen, K.; González, M.; Kearns, P.; Sintes, J.R.; Rossi, F.; Sayre, P. Review of achievements of the OECD Working Party on Manufactured Nanomaterials’ Testing and Assessment Programme. From exploratory testing to test guidelines. Regul. Toxicol. Pharmacol. 2016, 74, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Sarac, N.; Baygar, T.; Ugur, A. In vitro mutagenic and anti-mutagenic properties of green synthesised silver nanoparticles. IET Nanobiotechnol. 2018, 12, 230–233. [Google Scholar] [CrossRef]
- Fierascu, I.; Georgiev, M.I.; Ortan, A.; Fierascu, R.C.; Avramescu, S.M.; Ionescu, D.; Sutan, A.; Brinzan, A.; Ditu, L.M. Phyto-mediated metallic nano-architectures via Melissa officinalis L.: Synthesis, characterization and biological properties. Sci. Rep. 2017, 7, 12428. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.S.; Amin, T.; Katuva, S.; Kumari, M.; Selvaraj, K. Synthesis of water soluble CdS nanoparticles and study of their DNA damage activity. Arab. J. Chem. 2017, 10, S3929–S3935. [Google Scholar] [CrossRef] [Green Version]
- Barua, S.; Banerjee, P.P.; Sadhu, A.; Sengupta, A.; Chatterjee, S.; Sarkar, S.; Barman, S.; Chattopadhyay, A.; Battacharya, S.; Mondal, N.C.; et al. Silver nanoparticles as antibacterial and anticancer materials against human breast, cervical and oral cancer cells. J. Nanosci. Nanotechnol. 2017, 17, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Chetia, B.; Prasanth, R.; Madhavan, J.; Singaravelu, G.; Benelli, G.; Murugan, K. Green nanosynthesis and functionalization of gold nanoparticles as PTP 1B inhibitors. J. Clust. Sci. 2017, 28, 2269–2277. [Google Scholar] [CrossRef]
- Perde-Schrepler, M.; David, L.; Olenic, L.; Potara, M.; Fischer-Fodor, E.; Virag, P.; Imre-Lucaci, F.; Brie, I.; Florea, A. Gold nanoparticles synthesized with a polyphenols-rich extract from cornelian cherry (Cornus mas) fruits: Effects on human skin cells. J. Nanomater. 2016, 2016, 6986370. [Google Scholar] [CrossRef]
- Kalangi, S.K.; Dayakar, A.; Gangappa, D.; Sathyavathi, R.; Maurya, R.S.; Narayana Rao, D. Biocompatible silver nanoparticles reduced from Anethum graveolens leaf extract augments the antileishmanial efficacy of miltefosine. Exp. Parasitol. 2016, 170, 184–192. [Google Scholar] [CrossRef]
- Krishnaraj, C.; Harper, S.L.; Choe, H.S.; Kim, K.P.; Yun, S.I. Mechanistic aspects of biologically synthesized silver nanoparticles against food- and water-borne microbes. Bioprocess Biosyst. Eng. 2015, 38, 1943–1958. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, M.; Halder, D.; Mitra, A. Mechanistic study of antibacterial activity of biologically synthesized silver nanocolloids. Colloids Surf. A Physicochem. Eng. Asp. 2014, 449, 82–86. [Google Scholar] [CrossRef]
- Syed Ali, M.; Anuradha, V.; Sripriya, V.; Yogananth, N.; Sheeba, H. Evaluation of anti-genotoxicity of THE TiO2 nanoparticle biosynthesized from Turbinaria conoides. Int. J. Curr. Res. 2017, 9, 59085–59091. [Google Scholar]
- Vijaya, P.P.; Rekha, B.; Mathew, A.T.; Ali, M.S.; Yogananth, N.; Anuradha, V.; Parveen, P.K. Antigenotoxic effect of green-synthesised silver nanoparticles from Ocimum sanctum leaf extract against cyclophosphamide induced genotoxicity in human lymphocytes-in vitro. Appl. Nanosci. 2014, 4, 415–420. [Google Scholar] [CrossRef]
- Prasad, K.S.; Selvaraj, K. Biogenic synthesis of selenium nanoparticles and their effect on as(III)-induced toxicity on human lymphocytes. Biol. Trace Elem. Res. 2014, 157, 275–283. [Google Scholar] [CrossRef]
- Prasad, K.S.; Patel, H.; Patel, T.; Patel, K.; Selvaraj, K. Biosynthesis of Se nanoparticles and its effect on UV-induced DNA damage. Colloids Surf. Bbiointerfaces 2013, 103, 261–266. [Google Scholar] [CrossRef]
- Oberdorster, G. Safety assessment for nanotechnology and nanomedicine: Concepts of nanotoxicology. J. Intern. Med. 2010, 267, 89–105. [Google Scholar] [CrossRef]
- Singh, N.; Manshian, B.; Jenkins, G.J.; Griffiths, S.M.; Williams, P.M.; Maffeis, T.G.; Wright, C.J.; Doak, S.H. NanoGenotoxicology: The DNA damaging potential of engineered nanomaterials. Biomaterials 2009, 30, 3891–3914. [Google Scholar] [CrossRef]
- Magdolenova, Z.; Collins, A.; Kumar, A.; Dhawan, A.; Stone, V.; Dusinska, M. Mechanisms of genotoxicity. A review of in vitro and in vivo studies with engineered nanoparticles. Nanotoxicology 2014, 8, 233–278. [Google Scholar] [CrossRef]
- Mahaye, N.; Thwala, M.; Cowan, D.A.; Musee, N. Genotoxicity of metal based engineered nanoparticles in aquatic organisms: A review. Mutat. Res. 2017, 773, 134–160. [Google Scholar] [CrossRef]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 1195–1214. [Google Scholar] [CrossRef]
- Huang, S.; Chueh, P.J.; Lin, Y.W.; Shih, T.S.; Chuang, S.M. Disturbed mitotic progression and genome segregation are involved in cell transformation mediated by nano-TiO2 long-term exposure. Toxicol. Appl. Pharmacol. 2009, 241, 182–194. [Google Scholar] [CrossRef]
- Schins, R.P. Mechanisms of genotoxicity of particles and fibers. Inhal. Toxicol. 2002, 14, 57–78. [Google Scholar] [CrossRef]
- Stone, V.; Johnston, H.; Schins, R.P. Development of in vitro systems for nanotoxicology: Methodological considerations. Crit. Rev. Toxicol. 2009, 39, 613–626. [Google Scholar] [CrossRef]
- Sharifi, S.; Behzadi, S.; Laurent, S.; Forrest, M.L.; Stroeve, P.; Mahmoudi, M. Toxicity of nanomaterials. Chem. Soc. Rev. 2014, 41, 2323–2343. [Google Scholar] [CrossRef]
- Snyder, R.D.; Green, J.W. A review of the genotoxicity of marketed pharmaceuticals. Mutat. Res. Rev. Mutat. Res. 2001, 488, 151–169. [Google Scholar] [CrossRef]
- Bolt, H.M.; Stewart, J.D.; Hengstler, J.G. A comprehensive review about micronuclei: Mechanisms of formation and practical aspects in genotoxicity testing. Arch. Toxicol. 2011, 85, 861–862. [Google Scholar] [CrossRef]
- Speit, G.; Henderson, L. Review of the in vivo genotoxicity tests performed with styrene. Mutat. Res. Rev. Mutat. Res. 2005, 589, 67–79. [Google Scholar] [CrossRef]
- Brendler-Schwaab, S.; Hartmann, A.; Pfuhler, S.; Speit, G. The in vivo comet assay: Use and status in genotoxicity testing. Mutagenesis 2005, 20, 245–254. [Google Scholar] [CrossRef]
- Maity, P.; Bepari, M.; Pradhan, A.; Baral, R.; Roy, S.; Maiti Choudhury, S. Synthesis and characterization of biogenic metal nanoparticles and its cytotoxicity and anti-neoplasticity through the induction of oxidative stress, mitochondrial dysfunction and apoptosis. Colloids Surf. B Biointerfaces 2018, 161, 111–120. [Google Scholar] [CrossRef]
- Remya, R.R.; Rajasree, S.R.R.; Suman, T.Y.; Aranganathan, L.; Gayathri, S.; Gobalakrishnan, M.; Karthih, M.G. Laminarin based AgNPs using brown seaweed Turbinaria ornata and its induction of apoptosis in human retinoblastoma Y79 cancer cell lines. Mater. Res. Express 2018, 5. [Google Scholar] [CrossRef]
- Moteriya, P.; Chanda, S. Biosynthesis of silver nanoparticles formation from Caesalpinia pulcherrima stem metabolites and their broad spectrum biological activities. J. Genet. Eng. Biotechnol. 2018, 16, 105–113. [Google Scholar] [CrossRef]
- Moteriya, P.; Chanda, S. Synthesis and characterization of silver nanoparticles using Caesalpinia pulcherrima flower extract and assessment of their in vitro antimicrobial, antioxidant, cytotoxic, and genotoxic activities. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1556–1567. [Google Scholar] [CrossRef]
- Subbaiya, R.; Masilamani Selvam, M.; Sundar, K. Biological synthesis of silver nanorods from Nocardia mediterranei-5016 and its antitumor activity against non-small cell lung carcinoma cell line. Int. J. PharmTech Res. 2015, 8, 298–307. [Google Scholar]
- Lv, Q.; Zhang, B.; Xing, X.; Zhao, Y.; Cai, R.; Wang, W.; Gu, Q. Biosynthesis of copper nanoparticles using Shewanella loihica PV-4 with antibacterial activity: Novel approach and mechanisms investigation. J. Hazard. Mater. 2018, 347, 141–149. [Google Scholar] [CrossRef]
- Syed, B.; Bisht, N.; Bhat, P.S.; Nikhil Karthik, R.; Prasad, A.; Dhananjaya, B.L.; Satish, S.; Prasad, H.; Nagendra Prasad, N.N. Phytogenic synthesis of nanoparticles from Rhizophora mangle and their bactericidal potential with DNA damage activity. Nano Struct. Nano Objects 2017, 10, 112–115. [Google Scholar] [CrossRef]
- Gurunathan, S.; Jeong, J.K.; Han, J.W.; Zhang, X.F.; Park, J.H.; Kim, J.H. Multidimensional effects of biologically synthesized silver nanoparticles in Helicobacter pylori, Helicobacter felis, and human lung (L132) and lung carcinoma A549 cells. Nanoscale Res. Lett. 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Tamboli, D.P.; Lee, D.S. Mechanistic antimicrobial approach of extracellularly synthesized silver nanoparticles against gram positive and gram negative bacteria. J. Hazard. Mater. 2013, 260, 878–884. [Google Scholar] [CrossRef]
- Koca, F.D.; Duman, F. Genotoxic and cytotoxic activity of green synthesized TiO2 nanoparticles. Appl. Nanosci. 2018. [Google Scholar] [CrossRef]
- Chowdhury, S.; Basu, A.; Kundu, S. Green synthesis of protein capped silver nanoparticles from phytopathogenic fungus Macrophomina phaseolina (Tassi) Goid with antimicrobial properties against multidrug-resistant bacteria. Nanoscale Res. Lett. 2014, 9, 365. [Google Scholar] [CrossRef]
- Neveen, M.K. Biogenic silver nanoparticles by Aspergillus terreus as a powerful nanoweapon against Aspergillus fumigatus. Afr. J. Microbiol. Res. 2013, 7, 5645–5651. [Google Scholar] [CrossRef]
- Daphedar, A.; Taranath, T.C. Green synthesis of zinc nanoparticles using leaf extract of Albizia saman (Jacq.) Merr. and their effect on root meristems of Drimia indica (Roxb.) Jessop. Caryologia 2018, 71, 93–102. [Google Scholar] [CrossRef]
- Panda, K.K.; Golari, D.; Venugopal, A.; Achary, V.M.M.; Phaomei, G.; Parinandi, N.L.; Sahu, H.K.; Panda, B.B. Green Synthesized Zinc Oxide (ZnO) Nanoparticles Induce Oxidative Stress and DNA Damage in Lathyrus sativus L. Root Bioassay System. Antioxidants 2017, 6, 35. [Google Scholar] [CrossRef]
- Abdelsalam, N.R.; Abdel-Megeed, A.; Ali, H.M.; Salem, M.Z.M.; Al-Hayali, M.F.A.; Elshikh, M.S. Genotoxicity effects of silver nanoparticles on wheat (Triticum aestivum L.) root tip cells. Ecotoxicol. Environ. Saf. 2018, 155, 76–85. [Google Scholar] [CrossRef]
- Saha, N.; Dutta Gupta, S. Low-dose toxicity of biogenic silver nanoparticles fabricated by Swertia chirata on root tips and flower buds of Allium cepa. J. Hazard. Mater. 2017, 330, 18–28. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Gurunathan, S.; Chung, I.M. Physiological, metabolic, and transcriptional effects of biologically-synthesized silver nanoparticles in turnip (Brassica rapa ssp. rapa L.). Protoplasma 2015, 252, 1031–1046. [Google Scholar] [CrossRef]
- Adiguzel, A.O.; Adiguzel, S.K.; Mazmanci, B.; Tuncer, M.; Mazmanci, M.A. Silver nanoparticle biosynthesis from newly isolated streptomyces genus from soil. Mater. Res. Express. 2018, 5, 045402. [Google Scholar] [CrossRef]
- Pandiarajan, J.; Balaji, S. Genotoxic effect of silver nanoparticles in silk worm Bombyx mori. Austin J. Biotechnol. Bioeng. 2018, 5, 1096. [Google Scholar]
- Ishwarya, R.; Vaseeharan, B.; Shanthi, S.; Ramesh, S.; Manogari, P.; Dhanalakshmi, K.; Vijayakumar, S.; Benelli, G. Green synthesized silver nanoparticles: Toxicity against Poecilia reticulata fishes and Ceriodaphnia cornuta crustaceans. J. Clust. Sci. 2017, 28, 519–527. [Google Scholar] [CrossRef]
- Krishnaraj, C.; Harper, S.L.; Yun, S.I. In vivo toxicological assessment of biologically synthesized silver nanoparticles in adult Zebrafish (Danio rerio). J. Hazard. Mater. 2016, 301, 480–491. [Google Scholar] [CrossRef]
- Sukirtha, R.; Krishnan, M.; Ramachandran, R.; Kamalakkannan, S.; Kokilavani, P.; SankarGanesh, D.; Kannan, S.; Achiraman, S. Areca catechu Linn.-derived silver nanoparticles: A novel antitumor agent against Dalton’s ascites lymphoma. Int. J. Green Nanotechnol. Biomed. 2011, 3, 1–12. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barabadi, H.; Najafi, M.; Samadian, H.; Azarnezhad, A.; Vahidi, H.; Mahjoub, M.A.; Koohiyan, M.; Ahmadi, A. A Systematic Review of the Genotoxicity and Antigenotoxicity of Biologically Synthesized Metallic Nanomaterials: Are Green Nanoparticles Safe Enough for Clinical Marketing? Medicina 2019, 55, 439. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55080439
Barabadi H, Najafi M, Samadian H, Azarnezhad A, Vahidi H, Mahjoub MA, Koohiyan M, Ahmadi A. A Systematic Review of the Genotoxicity and Antigenotoxicity of Biologically Synthesized Metallic Nanomaterials: Are Green Nanoparticles Safe Enough for Clinical Marketing? Medicina. 2019; 55(8):439. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55080439
Chicago/Turabian StyleBarabadi, Hamed, Masoud Najafi, Hadi Samadian, Asaad Azarnezhad, Hossein Vahidi, Mohammad Ali Mahjoub, Mahbobeh Koohiyan, and Amirhossein Ahmadi. 2019. "A Systematic Review of the Genotoxicity and Antigenotoxicity of Biologically Synthesized Metallic Nanomaterials: Are Green Nanoparticles Safe Enough for Clinical Marketing?" Medicina 55, no. 8: 439. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55080439