Inhibition of Virus-Induced Cytokine Production from Airway Epithelial Cells by the Late Addition of Budesonide

 , , , , ,
, , , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Reagents
2.2. Real-Time RT-PCR
2.3. ELISA
2.4. Statistical Analysis
2.5. Ethics Statement
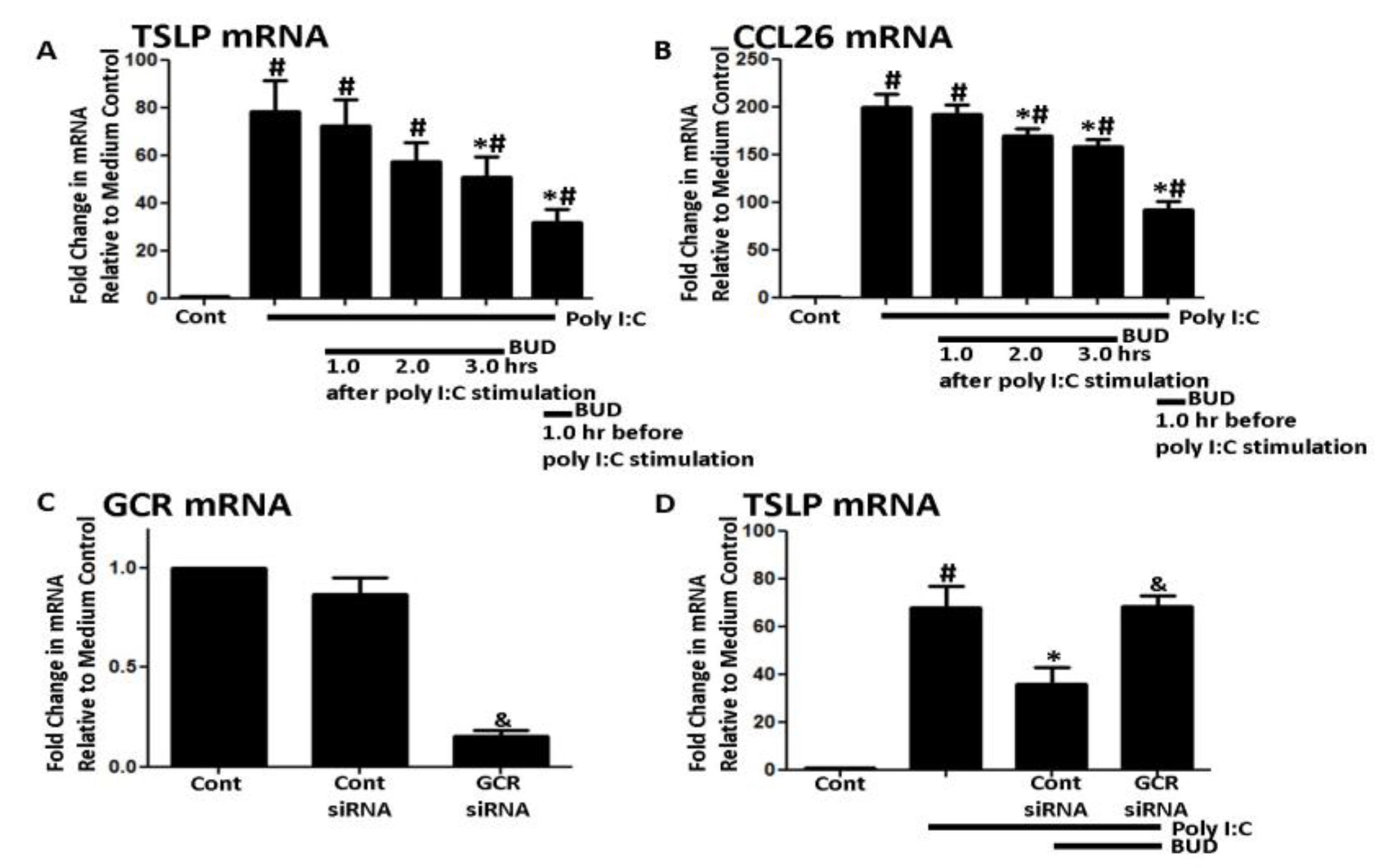
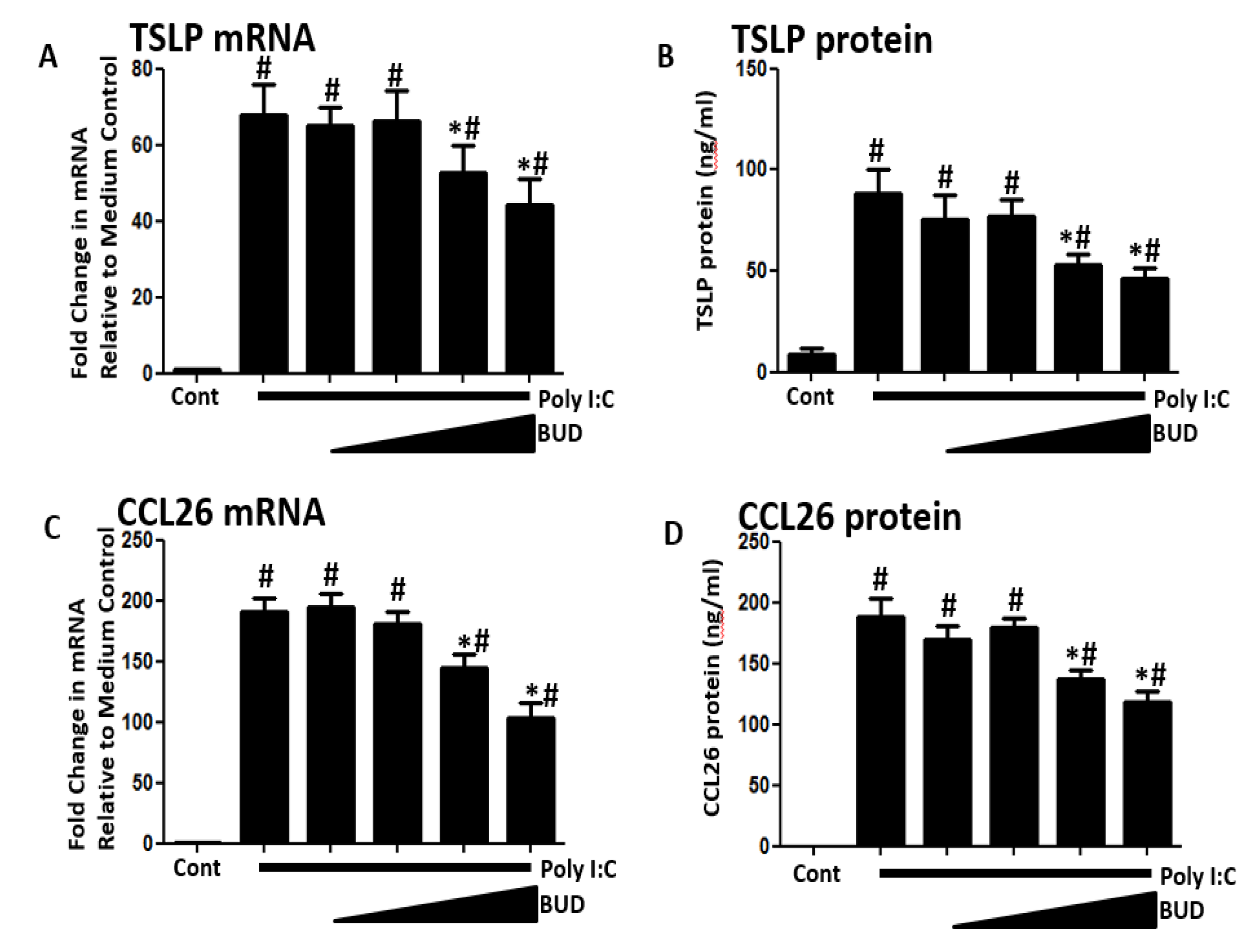
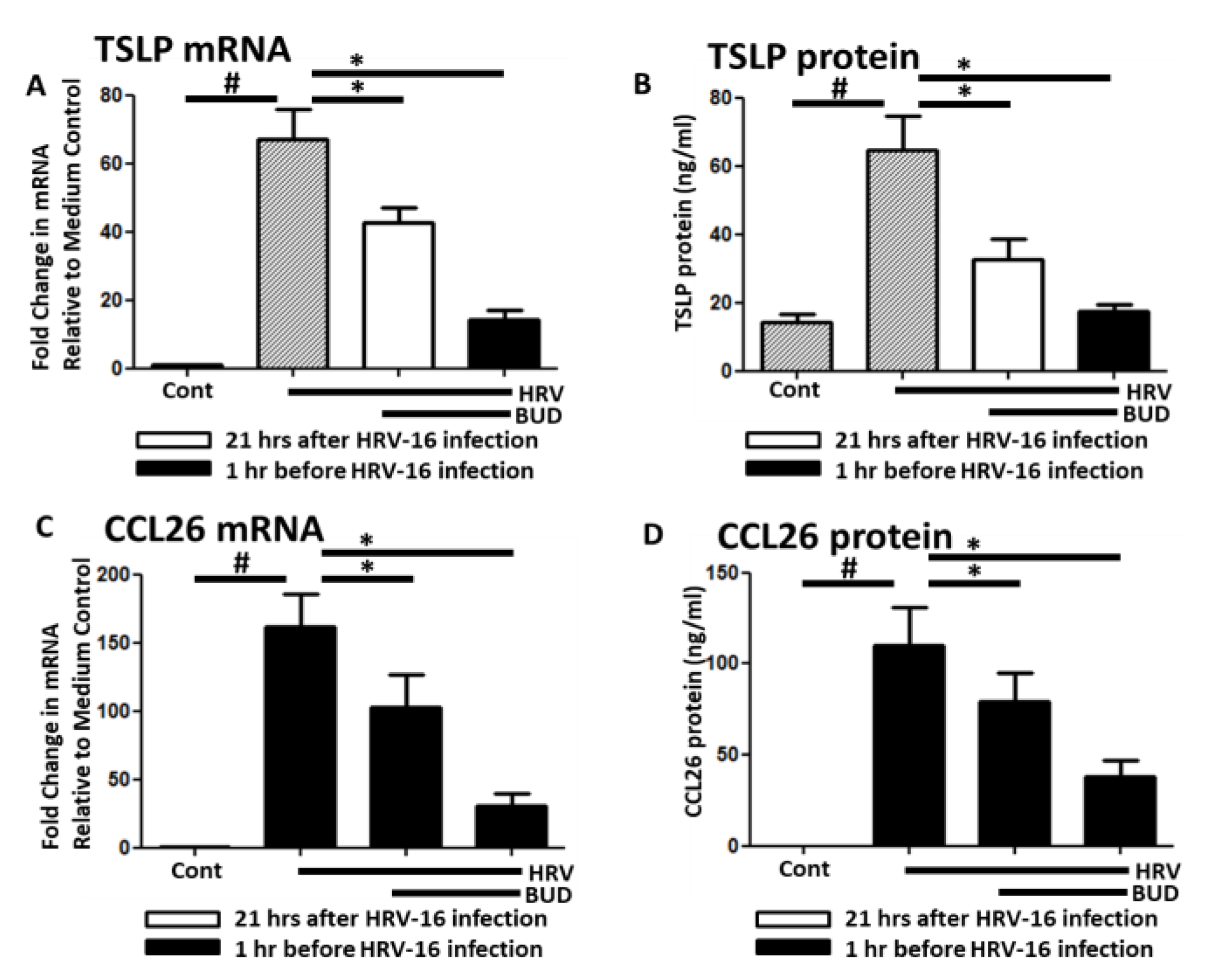
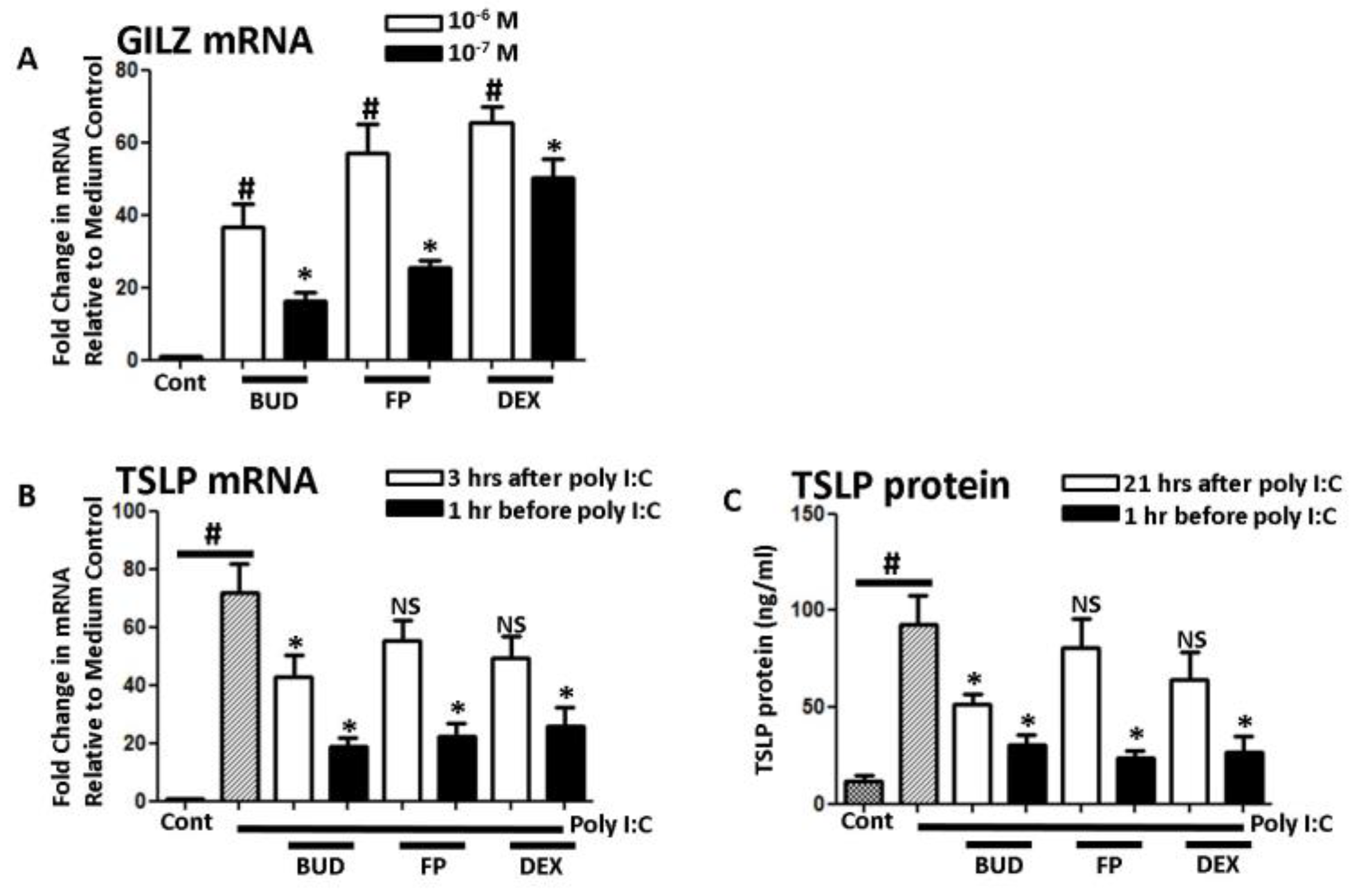
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schleimer, R.P.; Kato, A.; Kern, R.; Kuperman, D.; Avila, P.C. Epithelium: At the interface of innate and adaptive immune responses. J. Allergy Clin. Immunol. 2007, 120, 1279–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.K.; Klingler, A.I.; Poposki, J.A.; Stevens, W.W.; Peters, A.T.; Suh, L.A.; Norton, J.; Carter, R.G.; Hulse, K.E.; Harris, K.E.; et al. Heterogeneous inflammatory patterns in chronic rhinosinusitis without nasal polyps in Chicago, Illinois. J. Allergy Clin. Immunol. 2017, 139, 699–703.e7. [Google Scholar] [CrossRef] [Green Version]
- Tohidinik, H.R.; Mallah, N.; Takkouche, B. History of allergic rhinitis and risk of asthma; a systematic review and meta-analysis. World Allergy Organ. J. 2019, 12, 100069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Sullivan, S.M. Asthma death, CD8+ T cells, and viruses. Proc. Am. Thorac. Soc. 2005, 2, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. The cytokine network in asthma and chronic obstructive pulmonary disease. J. Clin. Investig. 2008, 118, 3546–3556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sha, Q.; Truong-Tran, A.Q.; Plitt, J.R.; Beck, L.A.; Schleimer, R.P. Activation of airway epithelial cells by toll-like receptor agonists. Am. J. Respir. Cell Mol. Biol. 2004, 31, 358–364. [Google Scholar] [CrossRef] [Green Version]
- Homma, T.; Kato, A.; Sakashita, M.; Norton, J.E.; Suh, L.A.; Carter, R.G.; Schleimer, R.P. Involvement of Toll-like receptor 2 and epidermal growth factor receptor signaling in epithelial expression of airway remodeling factors. Am. J. Respir. Cell Mol. Biol. 2015, 52, 471–481. [Google Scholar] [CrossRef] [Green Version]
- Homma, T.; Kato, A.; Sakashita, M.; Takabayashi, T.; Norton, J.E.; Suh, L.A.; Carter, R.G.; Harris, K.E.; Peters, A.T.; Grammer, L.C.; et al. Potential Involvement of the Epidermal Growth Factor Receptor Ligand Epiregulin and Matrix Metalloproteinase-1 in Pathogenesis of Chronic Rhinosinusitis. Am. J. Respir. Cell Mol. Biol. 2017, 57, 334–345. [Google Scholar] [CrossRef]
- Min, J.Y.; Ocampo, C.J.; Stevens, W.W.; Price, C.P.E.; Thompson, C.F.; Homma, T.; Huang, J.H.; Norton, J.E.; Suh, L.A.; Pothoven, K.L.; et al. Proton pump inhibitors decrease eotaxin-3/CCL26 expression in patients with chronic rhinosinusitis with nasal polyps: Possible role of the nongastric H,K-ATPase. J. Allergy Clin. Immunol. 2017, 139, 130–141.e11. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.J.; Makrinioti, H.; Rana, B.M.; Shamji, B.W.; Trujillo-Torralbo, M.B.; Footitt, J.; Del-Rosario, J.; Telcian, A.G.; Nikonova, A.; Zhu, J.; et al. IL-33-dependent type 2 inflammation during rhinovirus-induced asthma exacerbations in vivo. Am. J. Respir. Crit. Care Med. 2014, 190, 1373–1382. [Google Scholar] [CrossRef] [Green Version]
- Matsukura, S.; Kurokawa, M.; Homma, T.; Watanabe, S.; Suzuki, S.; Ieki, K.; Takeuchi, H.; Notomi, K.; Schleimer, R.P.; Kawaguchi, M.; et al. Basic research on virus-induced asthma exacerbation: Inhibition of inflammatory chemokine expression by fluticasone propionate. Int. Arch. Allergy Immunol. 2013, 161 (Suppl. 2), 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mochizuki, H.; Kusuda, S.; Okada, K.; Yoshihara, S.; Furuya, H.; Simões, E.A.F.; Scientific Committee for Elucidation of Infantile Asthma. Palivizumab Prophylaxis in Preterm Infants and Subsequent Recurrent Wheezing. Six-Year Follow-up Study. Am. J. Respir. Crit. Care Med. 2017, 196, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Matsukura, S.; Kokubu, F.; Kurokawa, M.; Kawaguchi, M.; Ieki, K.; Kuga, H.; Odaka, M.; Suzuki, S.; Watanabe, S.; Takeuchi, H.; et al. Synthetic double-stranded RNA induces multiple genes related to inflammation through Toll-like receptor 3 depending on NF-kappaB and/or IRF-3 in airway epithelial cells. Clin. Exp. Allergy 2006, 36, 1049–1062. [Google Scholar] [CrossRef]
- Homma, T.; Matsukura, S.; Hirose, T.; Ohnishi, T.; Kimura, T.; Kurokawa, M.; Ieki, K.; Odaka, M.; Suzuki, S.; Watanabe, S.; et al. Cooperative activation of CCL5 expression by TLR3 and tumor necrosis factor-alpha or interferon-gamma through nuclear factor-kappaB or STAT-1 in airway epithelial cells. Int. Arch. Allergy Immunol. 2010, 152 (Suppl. 1), 9–17. [Google Scholar] [CrossRef] [Green Version]
- Sethi, S.K.; Bianco, A.; Allen, J.T.; Knight, R.A.; Spiteri, M.A. Interferon-gamma (IFN-gamma) down-regulates the rhinovirus-induced expression of intercellular adhesion molecule-1 (ICAM-1) on human airway epithelial cells. Clin. Exp. Immunol. 1997, 110, 362–369. [Google Scholar] [CrossRef]
- Homma, T.; Kato, A.; Bhushan, B.; Norton, J.E.; Suh, L.A.; Carter, R.G.; Gupta, D.S.; Schleimer, R.P. Role of Aspergillus fumigatus in Triggering Protease-Activated Receptor-2 in Airway Epithelial Cells and Skewing the Cells toward a T-helper 2 Bias. Am. J. Respir. Cell Mol. Biol. 2016, 54, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Bhushan, B.; Homma, T.; Norton, J.E.; Sha, Q.; Siebert, J.; Gupta, D.S.; Schroeder, J.W., Jr.; Schleimer, R.P. Suppression of epithelial signal transducer and activator of transcription 1 activation by extracts of Aspergillus fumigatus. Am. J. Respir. Cell Mol. Biol. 2015, 53, 87–95. [Google Scholar] [CrossRef]
- Skevaki, C.L.; Christodoulou, I.; Spyridaki, I.S.; Tiniakou, I.; Georgiou, V.; Xepapadaki, P.; Kafetzis, D.A.; Papadopoulos, N.G. Budesonide and formoterol inhibit inflammatory mediator production by bronchial epithelial cells infected with rhinovirus. Clin. Exp. Allergy 2009, 39, 1700–1710. [Google Scholar] [CrossRef]
- Kato, A. Group 2 Innate Lymphoid Cells in Airway Diseases. Chest 2019, 156, 141–149. [Google Scholar] [CrossRef]
- Corren, J.; Parnes, J.R.; Wang, L.; Mo, M.; Roseti, S.L.; Griffiths, J.M.; van der Merwe, R. Tezepelumab in Adults with Uncontrolled Asthma. N. Engl. J. Med. 2017, 377, 936–946. [Google Scholar] [CrossRef]
- Caminati, M.; Polk, B.; Rosenwasser, L.J. What have recent advances in therapy taught us about severe asthma disease mechanisms? Expert Rev. Clin. Immunol. 2019, 15, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Matsukura, S.; Kokubu, F.; Kurokawa, M.; Kawaguchi, M.; Ieki, K.; Kuga, H.; Odaka, M.; Suzuki, S.; Watanabe, S.; Homma, T.; et al. Role of RIG-I, MDA-5, and PKR on the expression of inflammatory chemokines induced by synthetic dsRNA in airway epithelial cells. Int. Arch. Allergy Immunol. 2007, 143 (Suppl. 1), 80–83. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y.; Shiraishi, A.; Kobayashi, T.; Kadota, Y.; Shirakata, Y.; Hashimoto, K.; Ohashi, Y. Alteration of TLR3 pathways by glucocorticoids may be responsible for immunosusceptibility of human corneal epithelial cells to viral infections. Mol. Vis. 2009, 15, 937–948. [Google Scholar] [PubMed]
- Kim, S.R.; Song, J.H.; Ahn, J.H.; Lee, G.S.; Ahn, H.; Yoon, S.I.; Kang, S.G.; Kim, P.H.; Jeon, S.M.; Choi, E.J.; et al. Antiviral and anti-inflammatory activity of budesonide against human rhinovirus infection mediated via autophagy activation. Antivir. Res. 2018, 151, 87–96. [Google Scholar] [CrossRef]
- Bird, A.D.; McDougall, A.R.; Seow, B.; Hooper, S.B.; Cole, T.J. Glucocorticoid regulation of lung development: Lessons learned from conditional GR knockout mice. Mol. Endocrinol. 2015, 29, 158–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schleimer, R.P. Glucocorticoids suppress inflammation but spare innate immune responses in airway epithelium. Proc. Am. Thorac. Soc. 2004, 1, 222–230. [Google Scholar] [CrossRef]
- Edsbäcker, S.; Wollmer, P.; Selroos, O.; Borgström, L.; Olsson, B.; Ingelf, J. Do airway clearance mechanisms influence the local and systemic effects of inhaled corticosteroids? Pulm. Pharmacol. Ther. 2008, 21, 247–258. [Google Scholar] [CrossRef]
- Tashkin, D.P.; Lipworth, B.; Brattsand, R. Benefit: Risk Profile of Budesonide in Obstructive Airways Disease. Drugs 2019, 79, 1757–1775. [Google Scholar] [CrossRef] [Green Version]
- Ek, A.; Larsson, K.; Siljerud, S.; Palmberg, L. Fluticasone and budesonide inhibit cytokine release in human lung epithelial cells and alveolar macrophages. Allergy 1999, 54, 691–699. [Google Scholar] [CrossRef]
- Dalby, C.; Polanowski, T.; Larsson, T.; Borgström, L.; Edsbäcker, S.; Harrison, T.W. The bioavailability and airway clearance of the steroid component of budesonide/formoterol and salmeterol/fluticasone after inhaled administration in patients with COPD and healthy subjects: A randomized controlled trial. Respir. Res. 2009, 10, 104. [Google Scholar] [CrossRef] [Green Version]
- Heijink, I.H.; Jonker, M.R.; de Vries, M.; van Oosterhout, A.J.M.; Telenga, E.; Ten Hacken, N.H.T.; Postma, D.S.; van den Berge, M. Budesonide and fluticasone propionate differentially affect the airway epithelial barrier. Respir. Res. 2016, 17, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Berge, M.; Jonker, M.R.; Miller-Larsson, A.; Postma, D.S.; Heijink, I.H. Effects of fluticasone propionate and budesonide on the expression of immune defense genes in bronchial epithelial cells. Pulm. Pharmacol. Ther. 2018, 50, 47–56. [Google Scholar] [CrossRef]
- Beasley, R.; Holliday, M.; Reddel, H.K.; Braithwaite, I.; Ebmeier, S.; Hancox, R.J.; Harrison, T.; Houghton, C.; Oldfield, K.; Papi, A.; et al. Controlled Trial of Budesonide-Formoterol as Needed for Mild Asthma. N. Engl. J. Med. 2019, 23, 2020–2030. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.J.; Porritt, R.A.; Hertzog, P.J.; Bardin, P.G.; Tate, M.D. Glucocorticosteroids enhance replication of respiratory viruses: Effect of adjuvant interferon. Sci. Rep. 2014, 4, 7176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaya, M.; Nishimura, H.; Nadine, L.; Kubo, H.; Nagatomi, R. Formoterol and budesonide inhibit rhinovirus infection and cytokine production in primary cultures of human tracheal epithelial cells. Respir. Investig. 2014, 52, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Bochkov, Y.A.; Busse, W.W.; Brockman-Schneider, R.A.; Evans, M.D.; Jarjour, N.N.; McCrae, C.; Miller-Larsson, A.; Gern, J.E. Budesonide and formoterol effects on rhinovirus replication and epithelial cell cytokine responses. Respir. Res. 2013, 14, 98. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Homma, T.; Fukuda, Y.; Uchida, Y.; Uno, T.; Jinno, M.; Kishino, Y.; Yamamoto, M.; Sato, H.; Akimoto, K.; Kaneko, K.; et al. Inhibition of Virus-Induced Cytokine Production from Airway Epithelial Cells by the Late Addition of Budesonide. Medicina 2020, 56, 98. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina56030098
Homma T, Fukuda Y, Uchida Y, Uno T, Jinno M, Kishino Y, Yamamoto M, Sato H, Akimoto K, Kaneko K, et al. Inhibition of Virus-Induced Cytokine Production from Airway Epithelial Cells by the Late Addition of Budesonide. Medicina. 2020; 56(3):98. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina56030098
Chicago/Turabian StyleHomma, Tetsuya, Yosuke Fukuda, Yoshitaka Uchida, Tomoki Uno, Megumi Jinno, Yasunari Kishino, Mayumi Yamamoto, Hiroki Sato, Kaho Akimoto, Keisuke Kaneko, and et al. 2020. "Inhibition of Virus-Induced Cytokine Production from Airway Epithelial Cells by the Late Addition of Budesonide" Medicina 56, no. 3: 98. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina56030098