
Temporomandibular Disorders Slow Down the Regeneration Process of Masticatory Muscles: Transcriptomic Analysis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients Enrollment
2.2. Tissue Collection
2.3. Total RNA Extraction and cDNA Library Preparation
2.4. Differential Expression Analysis
3. Results
3.1. Anamnestic Data
3.2. Groups Comparison Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murphy, M.K.; MacBarb, R.F.; Wong, M.E.; Athanasiou, K.A. Temporomandibular disorders: A review of etiology, clinical management, and tissue engineering strategies. Int. J. Oral Maxillofac. Implant. 2013, 28, e393–e414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emodi-Perlman, A.; Eli, I.; Smardz, J.; Uziel, N.; Wieckiewicz, G.; Gilon, E.; Grychowska, N.; Wieckiewicz, M. Temporomandibular Disorders and Bruxism Outbreak as a Possible Factor of Orofacial Pain Worsening during the COVID-19 Pandemic-Concomitant Research in Two Countries. J. Clin. Med. 2020, 9, 3250. [Google Scholar] [CrossRef] [PubMed]
- Gauer, R.L.; Semidey, M.J. Diagnosis and treatment of temporomandibular disorders. Am. Fam. Physician 2015, 91, 378–386. [Google Scholar] [PubMed]
- LeResche, L. Epidemiology of temporomandibular disorders: Implications for the investigation of etiologic factors. Crit. Rev. Oral Biol. Med. 1997, 8, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Lobbezoo, F.; Ahlberg, J.; Glaros, A.G.; Kato, T.; Koyano, K.; Lavigne, G.J.; de Leeuw, R.; Manfredini, D.; Svensson, P.; Winocur, E. Bruxism defined and graded: An international consensus. J. Oral Rehabil. 2013, 40, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Ohlmann, B.; Waldecker, M.; Leckel, M.; Bomicke, W.; Behnisch, R.; Rammelsberg, P.; Schmitter, M. Correlations between Sleep Bruxism and Temporomandibular Disorders. J. Clin. Med. 2020, 9, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Steinkeler, A. Epidemiology, diagnosis, and treatment of temporomandibular disorders. Dent. Clin. N. Am. 2013, 57, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Cutroneo, G.; Vermiglio, G.; Centofanti, A.; Rizzo, G.; Runci, M.; Favaloro, A.; Piancino, M.G.; Bracco, P.; Ramieri, G.; Bianchi, F.; et al. Morphofunctional compensation of masseter muscles in unilateral posterior crossbite patients. Eur. J. Histochem. 2016, 60, 2605. [Google Scholar] [CrossRef]
- Vermiglio, G.; Centofanti, A.; Ramieri, G.; Tepedino, M.; Runci Anastasi, M.; Micali, A.G.; Arco, A.; Piancino, M.G. Immunofluorescence Evaluation of Myf5 and MyoD in Masseter Muscle of Unilateral Posterior Crossbite Patients. J. Funct. Morphol. Kinesiol. 2020, 5, 80. [Google Scholar] [CrossRef]
- Cascone, P.; Ramieri, V.; Arangio, P.; Vellone, V.; Tarsitano, A.; Marchetti, C. TMJ inferior compartment arthroplasty procedure through a 25-year follow-up (functional arthroplasty). Ann. Stomatol. 2016, 7, 60–64. [Google Scholar] [CrossRef]
- Graff-Radford, S.B.; Abbott, J.J. Temporomandibular Disorders and Headache. Oral Maxillofac. Surg. Clin. N. Am. 2016, 28, 335–349. [Google Scholar] [CrossRef]
- Ciancaglini, R.; Radaelli, G. The relationship between headache and symptoms of temporomandibular disorder in the general population. J. Dent. 2001, 29, 93–98. [Google Scholar] [CrossRef]
- Speciali, J.G.; Dach, F. Temporomandibular dysfunction and headache disorder. Headache 2015, 55 (Suppl. 1), 72–83. [Google Scholar] [CrossRef]
- Zhou, Z.; Kang, H.; Li, S.; Zhao, X. Understanding the neurotropic characteristics of SARS-CoV-2: From neurological manifestations of COVID-19 to potential neurotropic mechanisms. J. Neurol. 2020, 267, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-de-las-Penas, C.; Svensson, P. Myofascial Temporomandibular Disorder. Curr. Rheumatol. Rev. 2016, 12, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Costa, Y.M.; Ariji, Y.; Ferreira, D.; Bonjardim, L.R.; Conti, P.C.R.; Ariji, E.; Svensson, P. Muscle hardness and masticatory myofascial pain: Assessment and clinical relevance. J. Oral Rehabil. 2018, 45, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.K.; Ryu, J.W.; Choi, J.H.; Merrill, R.L.; Kim, S.T. Application of ICHD-II criteria for headaches in a TMJ and orofacial pain clinic. Cephalalgia 2010, 30, 37–41. [Google Scholar] [CrossRef]
- Sung, Y.T.; Wu, J.S. The Visual Analogue Scale for Rating, Ranking and Paired-Comparison (VAS-RRP): A new technique for psychological measurement. Behav. Res. Methods 2018, 50, 1694–1715. [Google Scholar] [CrossRef] [PubMed]
- Luedtke, K.; Allers, A.; Schulte, L.H.; May, A. Efficacy of interventions used by physiotherapists for patients with headache and migraine-systematic review and meta-analysis. Cephalalgia 2016, 36, 474–492. [Google Scholar] [CrossRef]
- Silvestro, S.; Chiricosta, L.; Gugliandolo, A.; Pizzicannella, J.; Diomede, F.; Bramanti, P.; Trubiani, O.; Mazzon, E. Extracellular Vesicles Derived from Human Gingival Mesenchymal Stem Cells: A Transcriptomic Analysis. Genes 2020, 11, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.A., Jr.; Brandao, K.V.; Faleiros, B.E.; Tavares, R.M.; Lara, R.P.; Januzzi, E.; Carvalho, A.B.; Carvalho, E.M.; Gomes, J.B.; Leite, F.M.; et al. Temporo-mandibular disorders are an important comorbidity of migraine and may be clinically difficult to distinguish them from tension-type headache. Arq. Neuropsiquiatr. 2014, 72, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florencio, L.L.; de Oliveira, A.S.; Carvalho, G.F.; Dach, F.; Bigal, M.E.; Fernandez-de-Las-Penas, C.; Bevilaqua-Grossi, D. Association Between Severity of Temporomandibular Disorders and the Frequency of Headache Attacks in Women With Migraine: A Cross-Sectional Study. J. Manip. Physiol. Ther. 2017, 40, 250–254. [Google Scholar] [CrossRef]
- Kuijpers, M.A.; Grefte, S.; Bronkhorst, E.M.; Carels, C.E.; Kiliaridis, S.; Von den Hoff, J.W. Reduced masticatory function is related to lower satellite cell numbers in masseter muscle. Eur. J. Orthod. 2014, 36, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grefte, S.; Kuijpers-Jagtman, A.M.; Torensma, R.; Von den Hoff, J.W. Skeletal muscle development and regeneration. Stem Cells Dev. 2007, 16, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Alhendi, A.M.N.; Patrikakis, M.; Daub, C.O.; Kawaji, H.; Itoh, M.; de Hoon, M.; Carninci, P.; Hayashizaki, Y.; Arner, E.; Khachigian, L.M. Promoter Usage and Dynamics in Vascular Smooth Muscle Cells Exposed to Fibroblast Growth Factor-2 or Interleukin-1beta. Sci. Rep. 2018, 8, 13164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miano, J.M.; Vlasic, N.; Tota, R.R.; Stemerman, M.B. Localization of Fos and Jun proteins in rat aortic smooth muscle cells after vascular injury. Am. J. Pathol. 1993, 142, 715–724. [Google Scholar] [PubMed]
- Ramachandran, A.; Gong, E.M.; Pelton, K.; Ranpura, S.A.; Mulone, M.; Seth, A.; Gomez, P., 3rd; Adam, R.M. FosB regulates stretch-induced expression of extracellular matrix proteins in smooth muscle. Am. J. Pathol. 2011, 179, 2977–2989. [Google Scholar] [CrossRef]
- Gaut, L.; Robert, N.; Delalande, A.; Bonnin, M.A.; Pichon, C.; Duprez, D. EGR1 Regulates Transcription Downstream of Mechanical Signals during Tendon Formation and Healing. PLoS ONE 2016, 11, e0166237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Chen, J.; Li, L.; Ren, X.; Cheng, T.; Lu, S.; Lawal, R.A.; Nie, Q.; Zhang, X.; Hanotte, O. c-Myc inhibits myoblast differentiation and promotes myoblast proliferation and muscle fibre hypertrophy by regulating the expression of its target genes, miRNAs and lincRNAs. Cell Death Differ. 2019, 26, 426–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Estevez, M.; Staats, J.; Paatela, E.; Munson, D.; Katoku-Kikyo, N.; Yuan, C.; Asakura, Y.; Hostager, R.; Kobayashi, H.; Asakura, A.; et al. Promotion of Myoblast Differentiation by Fkbp5 via Cdk4 Isomerization. Cell Rep. 2018, 25, 2537–2551.e2538. [Google Scholar] [CrossRef] [Green Version]
- Meyfour, A.; Ansari, H.; Pahlavan, S.; Mirshahvaladi, S.; Rezaei-Tavirani, M.; Gourabi, H.; Baharvand, H.; Salekdeh, G.H. Y Chromosome Missing Protein, TBL1Y, May Play an Important Role in Cardiac Differentiation. J. Proteome Res. 2017, 16, 4391–4402. [Google Scholar] [CrossRef] [PubMed]
- Abdelhaleem, M. RNA helicases: Regulators of differentiation. Clin. Biochem. 2005, 38, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Posey, J.E.; Rosenfeld, J.A.; Bacino, C.A.; Scaglia, F.; Immken, L.; Harris, J.M.; Hickey, S.E.; Mosher, T.M.; Slavotinek, A.; et al. Phenotypic expansion in DDX3X-a common cause of intellectual disability in females. Ann. Clin. Transl. Neurol. 2018, 5, 1277–1285. [Google Scholar] [CrossRef]
- Ravel-Chapuis, A.; Klein Gunnewiek, A.; Belanger, G.; Crawford Parks, T.E.; Cote, J.; Jasmin, B.J. Staufen1 impairs stress granule formation in skeletal muscle cells from myotonic dystrophy type 1 patients. Mol. Biol. Cell 2016, 27, 1728–1739. [Google Scholar] [CrossRef] [PubMed]
- Bergen, A.W.; Pratt, M.; Mehlman, P.T.; Goldman, D. Evolution of RPS4Y. Mol. Biol. Evol. 1998, 15, 1412–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakay, M.; Zhao, P.; Chen, J.; Hoffman, E.P. A web-accessible complete transcriptome of normal human and DMD muscle. Neuromuscul. Disord. 2002, 12 (Suppl. 1), S125–S141. [Google Scholar] [CrossRef]
- Gazova, I.; Lengeling, A.; Summers, K.M. Lysine demethylases KDM6A and UTY: The X and Y of histone demethylation. Mol. Genet. Metab. 2019, 127, 31–44. [Google Scholar] [CrossRef]
- Meyfour, A.; Pahlavan, S.; Ansari, H.; Baharvand, H.; Salekdeh, G.H. Down-Regulation of a Male-Specific H3K4 Demethylase, KDM5D, Impairs Cardiomyocyte Differentiation. J. Proteome Res. 2019, 18, 4277–4282. [Google Scholar] [CrossRef]
- Hausburg, M.A.; Doles, J.D.; Clement, S.L.; Cadwallader, A.B.; Hall, M.N.; Blackshear, P.J.; Lykke-Andersen, J.; Olwin, B.B. Post-transcriptional regulation of satellite cell quiescence by TTP-mediated mRNA decay. eLife 2015, 4, e03390. [Google Scholar] [CrossRef]
- Bye, A.J.H.; Pugazhendhi, D.; Woodhouse, S.; Brien, P.; Watson, R.; Turner, M.; Pell, J. The RNA-binding proteins Zfp36l1 and Zfp36l2 act redundantly in myogenesis. Skelet. Muscle 2018, 8, 37. [Google Scholar] [CrossRef]
- Karu, I.; Tahepold, P.; Ruusalepp, A.; Reimann, E.; Koks, S.; Starkopf, J. Exposure to sixty minutes of hyperoxia upregulates myocardial humanins in patients with coronary artery disease-a pilot study. J. Physiol. Pharmacol. 2015, 66, 899–906. [Google Scholar] [PubMed]
- Ponsuksili, S.; Siengdee, P.; Du, Y.; Trakooljul, N.; Murani, E.; Schwerin, M.; Wimmers, K. Identification of common regulators of genes in co-expression networks affecting muscle and meat properties. PLoS ONE 2015, 10, e0123678. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.E.; Newcomb, P.V.; Holly, J.M. Multifaceted roles of TNF-alpha in myoblast destruction: A multitude of signal transduction pathways. J. Cell. Physiol. 2004, 198, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, W.; Zhang, Y.; Zhang, F.; Huang, K. PARP1 promote autophagy in cardiomyocytes via modulating FoxO3a transcription. Cell Death Dis. 2018, 9, 1047. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Z.; Tan, Y.; Li, Q.; Yang, H.; Wang, P.; Lu, J.; Liu, P. PARP1 interacts with STAT3 and retains active phosphorylated-STAT3 in nucleus during pathological myocardial hypertrophy. Mol. Cell. Endocrinol. 2018, 474, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, X.; Chen, L.; Chen, W.; Zhang, Y.; Chen, J.; Wu, X.; Zhao, Y.; Wu, X.; Sun, G. The long noncoding RNA XIST protects cardiomyocyte hypertrophy by targeting miR-330-3p. Biochem. Biophys. Res. Commun. 2018, 505, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Marti-Pamies, I.; Canes, L.; Alonso, J.; Rodriguez, C.; Martinez-Gonzalez, J. The nuclear receptor NOR-1/NR4A3 regulates the multifunctional glycoprotein vitronectin in human vascular smooth muscle cells. FASEB J. 2017, 31, 4588–4599. [Google Scholar] [CrossRef] [Green Version]
- Ferran, B.; Marti-Pamies, I.; Alonso, J.; Rodriguez-Calvo, R.; Aguilo, S.; Vidal, F.; Rodriguez, C.; Martinez-Gonzalez, J. The nuclear receptor NOR-1 regulates the small muscle protein, X-linked (SMPX) and myotube differentiation. Sci. Rep. 2016, 6, 25944. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Jiang, M.; Wang, Z. TPM4 promotes cell migration by modulating F-actin formation in lung cancer. Onco Targets Ther. 2019, 12, 4055–4063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matyushenko, A.M.; Shchepkin, D.V.; Kopylova, G.V.; Bershitsky, S.Y.; Koubassova, N.A.; Tsaturyan, A.K.; Levitsky, D.I. Functional role of the core gap in the middle part of tropomyosin. FEBS J. 2018, 285, 871–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlahovich, N.; Schevzov, G.; Nair-Shaliker, V.; Ilkovski, B.; Artap, S.T.; Joya, J.E.; Kee, A.J.; North, K.N.; Gunning, P.W.; Hardeman, E.C. Tropomyosin 4 defines novel filaments in skeletal muscle associated with muscle remodelling/regeneration in normal and diseased muscle. Cell Motil. Cytoskelet. 2008, 65, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Frade, A.F.; Teixeira, P.C.; Ianni, B.M.; Pissetti, C.W.; Saba, B.; Wang, L.H.; Kuramoto, A.; Nogueira, L.G.; Buck, P.; Dias, F.; et al. Polymorphism in the alpha cardiac muscle actin 1 gene is associated to susceptibility to chronic inflammatory cardiomyopathy. PLoS ONE 2013, 8, e83446. [Google Scholar] [CrossRef] [PubMed]
- Boutilier, J.K.; Taylor, R.L.; Ram, R.; McNamara, E.; Nguyen, Q.; Goullee, H.; Chandler, D.; Mehta, M.; Balmer, L.; Laing, N.G.; et al. Variable cardiac alpha-actin (Actc1) expression in early adult skeletal muscle correlates with promoter methylation. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Laing, N.G.; Dye, D.E.; Wallgren-Pettersson, C.; Richard, G.; Monnier, N.; Lillis, S.; Winder, T.L.; Lochmuller, H.; Graziano, C.; Mitrani-Rosenbaum, S.; et al. Mutations and polymorphisms of the skeletal muscle alpha-actin gene (ACTA1). Hum. Mutat. 2009, 30, 1267–1277. [Google Scholar] [CrossRef] [Green Version]
- Ito, N.; Ruegg, U.T.; Takeda, S. ATP-Induced Increase in Intracellular Calcium Levels and Subsequent Activation of mTOR as Regulators of Skeletal Muscle Hypertrophy. Int. J. Mol. Sci. 2018, 19, 2804. [Google Scholar] [CrossRef] [Green Version]
- Raffaello, A.; Milan, G.; Masiero, E.; Carnio, S.; Lee, D.; Lanfranchi, G.; Goldberg, A.L.; Sandri, M. JunB transcription factor maintains skeletal muscle mass and promotes hypertrophy. J. Cell Biol. 2010, 191, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Peris-Moreno, D.; Taillandier, D.; Polge, C. MuRF1/TRIM63, Master Regulator of Muscle Mass. Int. J. Mol. Sci. 2020, 21, 6663. [Google Scholar] [CrossRef] [PubMed]
- Sacheck, J.M.; Ohtsuka, A.; McLary, S.C.; Goldberg, A.L. IGF-I stimulates muscle growth by suppressing protein breakdown and expression of atrophy-related ubiquitin ligases, atrogin-1 and MuRF1. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E591–E601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mevissen, T.E.; Hospenthal, M.K.; Geurink, P.P.; Elliott, P.R.; Akutsu, M.; Arnaudo, N.; Ekkebus, R.; Kulathu, Y.; Wauer, T.; El Oualid, F.; et al. OTU deubiquitinases reveal mechanisms of linkage specificity and enable ubiquitin chain restriction analysis. Cell 2013, 154, 169–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustamante, M.; Fernandez-Verdejo, R.; Jaimovich, E.; Buvinic, S. Electrical stimulation induces IL-6 in skeletal muscle through extracellular ATP by activating Ca(2+) signals and an IL-6 autocrine loop. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E869–E882. [Google Scholar] [CrossRef] [Green Version]
- Serrano, A.L.; Baeza-Raja, B.; Perdiguero, E.; Jardi, M.; Munoz-Canoves, P. Interleukin-6 is an essential regulator of satellite cell-mediated skeletal muscle hypertrophy. Cell Metab. 2008, 7, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Swiderski, K.; Thakur, S.S.; Naim, T.; Trieu, J.; Chee, A.; Stapleton, D.I.; Koopman, R.; Lynch, G.S. Muscle-specific deletion of SOCS3 increases the early inflammatory response but does not affect regeneration after myotoxic injury. Skelet. Muscle 2016, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Castrillon, E.E.; Cairns, B.E.; Wang, K.; Arendt-Nielsen, L.; Svensson, P. Comparison of glutamate-evoked pain between the temporalis and masseter muscles in men and women. Pain 2012, 153, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Mathe, G.; Couvreur, P.; Tew, K.D., II. Glutamine and glutamate. Biomed. Pharmacother. 2002, 56, 446–457. [Google Scholar] [CrossRef]
- Hayes, J.D.; Pulford, D.J. The glutathione S-transferase supergene family: Regulation of GST and the contribution of the isoenzymes to cancer chemoprotection and drug resistance. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 445–600. [Google Scholar] [CrossRef]
- Tripathi, G.M.; Kalita, J.; Misra, U.K. A study of oxidative stress in migraine with special reference to prophylactic therapy. Int. J. Neurosci. 2018, 128, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Merker, S.; Reif, A.; Ziegler, G.C.; Weber, H.; Mayer, U.; Ehlis, A.C.; Conzelmann, A.; Johansson, S.; Muller-Reible, C.; Nanda, I.; et al. SLC2A3 single-nucleotide polymorphism and duplication influence cognitive processing and population-specific risk for attention-deficit/hyperactivity disorder. J. Child Psychol. Psychiatry 2017, 58, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Gaster, M.; Beck-Nielsen, H.; Schroder, H.D. Regenerating human muscle fibres express GLUT3 protein. Pflugers. Arch. 2002, 445, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Yang, H.; Zhou, L.; Yang, F. Glucose-Sensing Transcription Factor MondoA/ChREBP as Targets for Type 2 Diabetes: Opportunities and Challenges. Int. J. Mol. Sci. 2019, 20, 5132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesari, M.; Penninx, B.W.; Lauretani, F.; Russo, C.R.; Carter, C.; Bandinelli, S.; Atkinson, H.; Onder, G.; Pahor, M.; Ferrucci, L. Hemoglobin levels and skeletal muscle: Results from the InCHIANTI study. J. Gerontol. A Biol. Sci. Med. Sci. 2004, 59, 249–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shephard, F.; Greville-Heygate, O.; Marsh, O.; Anderson, S.; Chakrabarti, L. A mitochondrial location for haemoglobins--dynamic distribution in ageing and Parkinson’s disease. Mitochondrion 2014, 14, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Oyaizu, T.; Enomoto, M.; Horie, M.; Yuasa, M.; Okawa, A.; Yagishita, K. VEGF and bFGF induction by nitric oxide is associated with hyperbaric oxygen-induced angiogenesis and muscle regeneration. Sci. Rep. 2020, 10, 2744. [Google Scholar] [CrossRef]


{kind=link}
| Variables | Patient 1 | Patient 2 | Patient 3 | Patient 4 | Patient 5 |
|---|---|---|---|---|---|
| Gender | Male | Male | Female | Female | Male |
| Age | 52 | 68 | 59 | 68 | 62 |
| Familiar headache | No | No | No | No | No |
| Head Pain Perception | Bilateral | Bilateral | Bilateral | Bilateral | - |
| VAS | 2/10 | 6/10 | 9/10 | 6/10 | 0 |
| Migraine Attack duration (without drug) | 2 h | 4 h | 24 h | 10 h | - |
| Nausea | 0 | 0 | 7/10 | 2/10 | 0 |
| Vomit | 0 | 0 | 6/10 | 2/10 | 0 |
| Photophobia | 0 | 4/10 | 8/10 | 4/10 | 0 |
| Phonophobia | 0 | 3/10 | 8/10 | 4/10 | 0 |
| Osmo-phobia | 2/10 | 0 | 8/10 | 3/10 | 0 |
| Tearing | 0 | 1/10 | 7/10 | 3/10 | 0 |
| Rhinorrhea | 0 | 0 | 6/10 | 2/10 | 0 |
| Difficult moving | 0 | 0 | 6/10 | 2/10 | 0 |
| Sensory processing disorder | 0 | 0 | 6/10 | 0 | 0 |
| Dysphasia | 0 | 0 | 4/10 | 0 | 0 |
| Limitation Activity daily for headache | No | No | No | No | No |
| Analgesic taken in a month | 1–3 | 1–3 | 4–10 | 4–10 | 0 |
| Analgesic efficacy during migraine attack | Yes | Yes | No | Yes | - |
| Migraine attacks in the last three months | 0 | 10 | 80 | 50 | 0 |
| Drugs used for migraine attacks | NSAID | NSAID | NSAID | NSAID | - |
| Professional status | Worker | Non- worker | Non- worker | Non- worker | Worker |
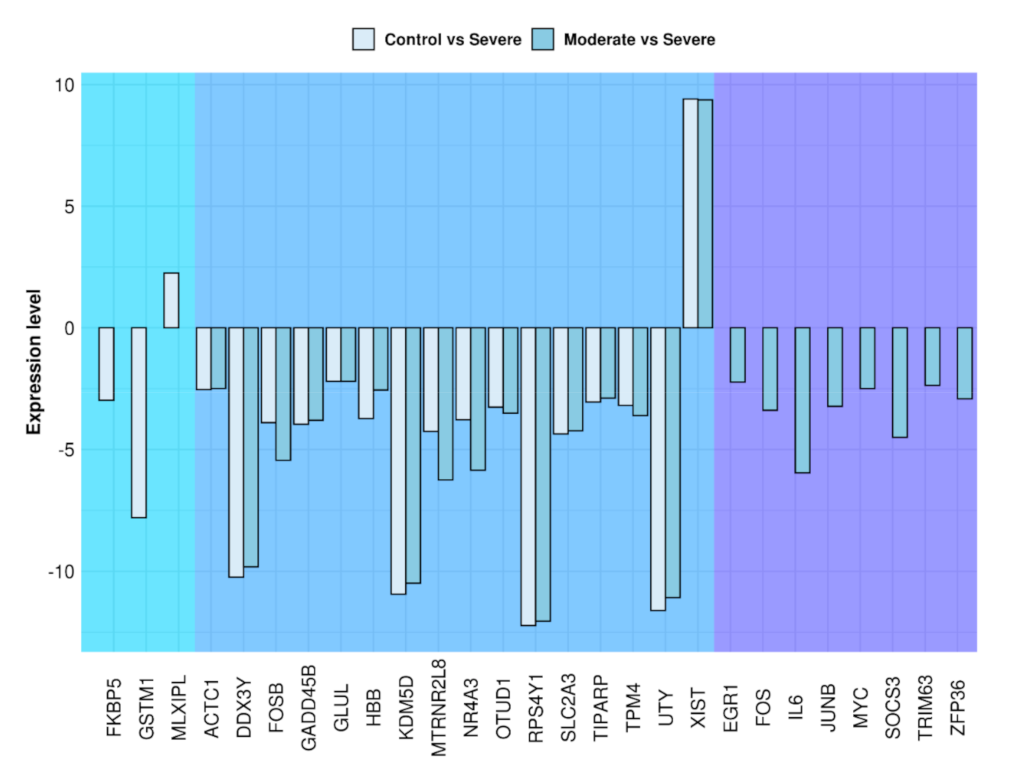
| Gene | Name | Fold Change Control vs. Severe | Fold Change Moderate vs. Severe |
|---|---|---|---|
| ACTC1 | actin alpha cardiac muscle 1 | −2.54 | −2.49 |
| DDX3Y | DEAD-box helicase 3 Y-linked | −10.24 | −9.82 |
| EGR1 | early growth response 1 | N.S. | −2.23 |
| FKBP5 | FKBP prolyl isomerase 5 | −2.98 | N.S. |
| FOS | FOS proto-oncogene, AP-1 transcription factor subunit | N.S. | −3.39 |
| FOSB | FOS proto-oncogene, AP-1 transcription factor subunit | −3.90 | −5.44 |
| GADD45B | growth arrest and DNA damage inducible beta | −3.96 | −3.80 |
| GLUL | glutamate-ammonia ligase | −2.20 | −2.20 |
| GSTM1 | glutathione S-transferase mu 1 | −7.80 | N.S. |
| HBB | hemo-globin subunit beta | −3.73 | −2.56 |
| IL6 | interleukin 6 | N.S. | −5.96 |
| JUNB | JunB proto-oncogene, AP-1 transcription factor subunit | N.S. | −3.23 |
| KDM5D | lysine demethylase 5D | −10.94 | −10.49 |
| MLXIPL | MLX interacting protein like | 2.25 | N.S. |
| MTRNR2L8 | MT-RNR2 like 8 | −4.26 | −6.25 |
| MYC | MYC proto-oncogene, bHLH transcription factor | N.S. | −2.50 |
| NR4A3 | nuclear receptor subfamily 4 group A member 3 | −3.78 | −5.85 |
| OTUD1 | OTU deubiquitinase 1 | −3.26 | −3.51 |
| RPS4Y1 | ribosomal protein S4 Y-linked 1 | −12.23 | −12.05 |
| SLC2A3 | solute carrier family 2 member 3 | −4.36 | −4.23 |
| SOCS3 | suppressor of cytokine signalling 3 | N.S. | −4.50 |
| TIPARP | TCDD inducible poly(ADP-ribose) polymerase | −3.05 | −2.89 |
| TPM4 | tropomyosin 4 | −3.19 | −3.61 |
| TRIM63 | tripartite motif containing 63 | N.S. | −2.37 |
| UTY | ubiquitously transcribed tetratricopeptide repeat containing, Y-linked | −11.61 | −11.08 |
| XIST | X inactive specific transcript | 9.41 | 9.37 |
| ZFP36 | ZFP36 ring finger protein | N.S. | −2.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sindona, C.; Runci Anastasi, M.; Chiricosta, L.; Gugliandolo, A.; Silvestro, S.; Bramanti, P.; Cascone, P.; Mazzon, E. Temporomandibular Disorders Slow Down the Regeneration Process of Masticatory Muscles: Transcriptomic Analysis. Medicina 2021, 57, 354. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57040354
Sindona C, Runci Anastasi M, Chiricosta L, Gugliandolo A, Silvestro S, Bramanti P, Cascone P, Mazzon E. Temporomandibular Disorders Slow Down the Regeneration Process of Masticatory Muscles: Transcriptomic Analysis. Medicina. 2021; 57(4):354. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57040354
Chicago/Turabian StyleSindona, Cinzia, Michele Runci Anastasi, Luigi Chiricosta, Agnese Gugliandolo, Serena Silvestro, Placido Bramanti, Piero Cascone, and Emanuela Mazzon. 2021. "Temporomandibular Disorders Slow Down the Regeneration Process of Masticatory Muscles: Transcriptomic Analysis" Medicina 57, no. 4: 354. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57040354