
Lipofection of Single Guide RNA Targeting MMP8 Decreases Proliferation and Migration in Lung Adenocarcinoma Cells

 , , , , and
, , , , and 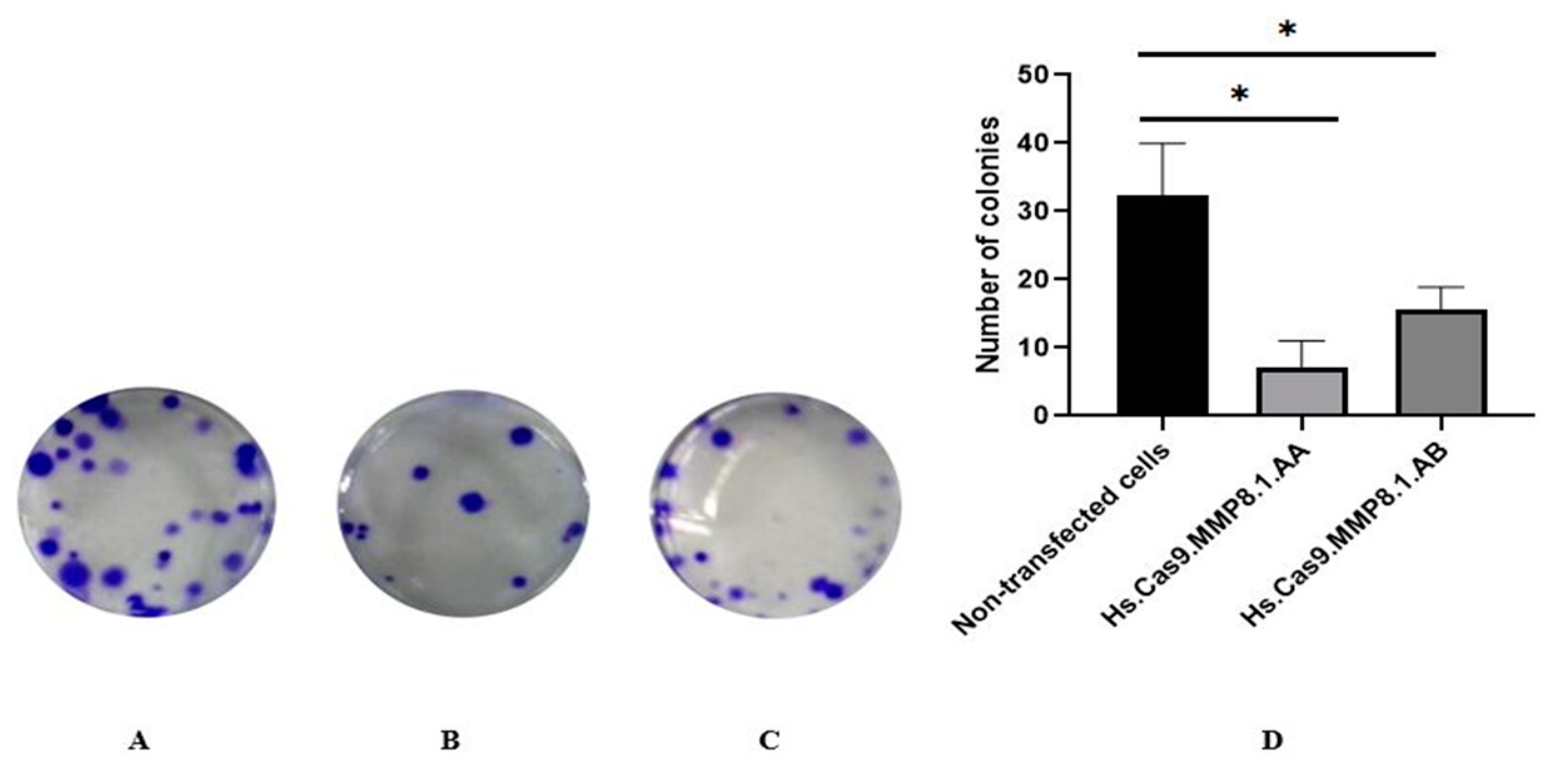{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Transfection of gRNA and Cas9 Protein Complexes into A549 Cell Line
2.3. Colony-Forming Assay
2.4. Cell Migration Assay
2.5. Cell Viability Assay
2.6. RNA Isolation and Relative mRNA Expression
2.7. Statistical Analysis
3. Results
3.1. Treated Cells Showed Reduced Cellular Adhesion and Migration Compared to Non-Transfected Cells
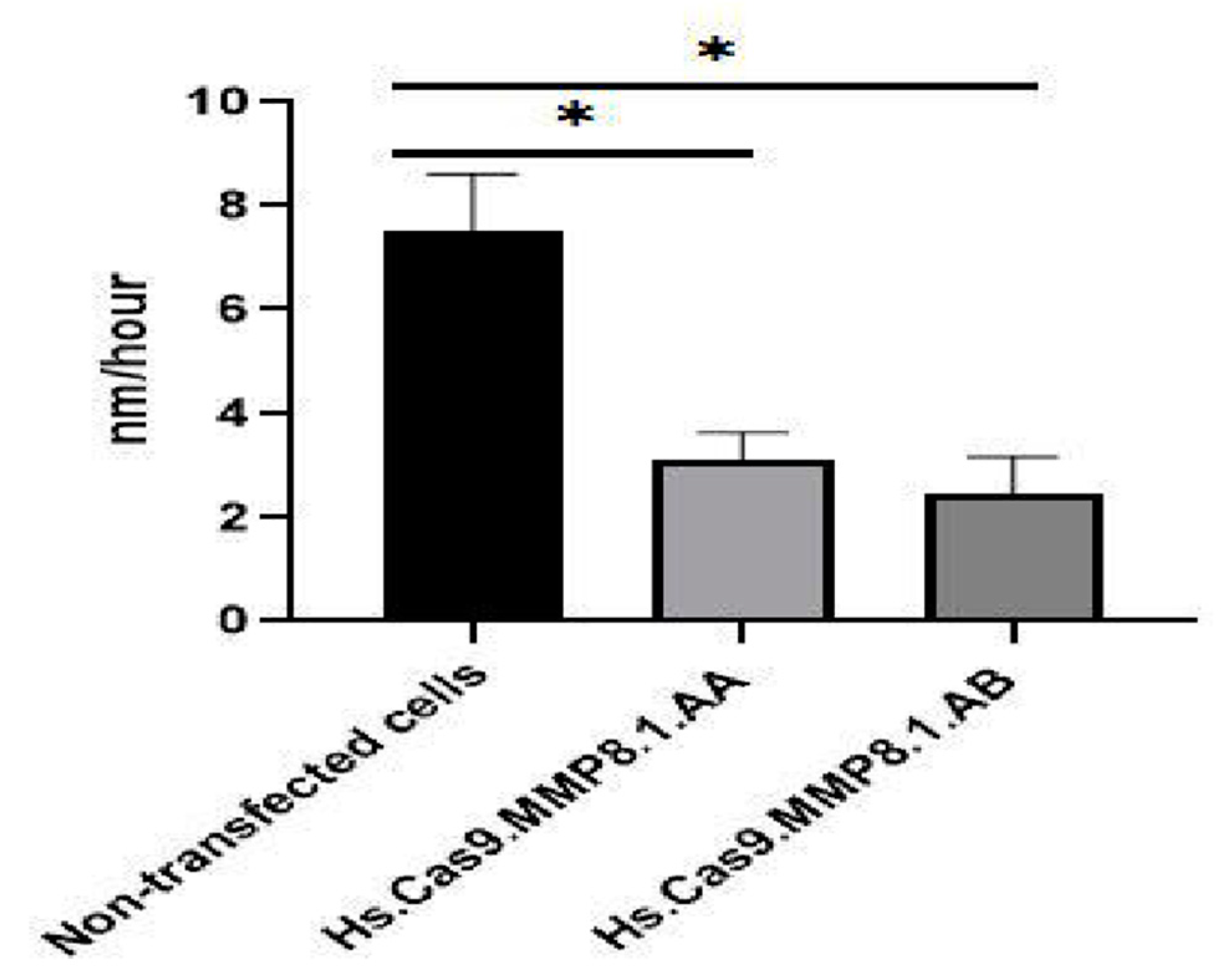
3.2. Treatment Decreased the Migration Potential Compared to Non-Transfected Cells
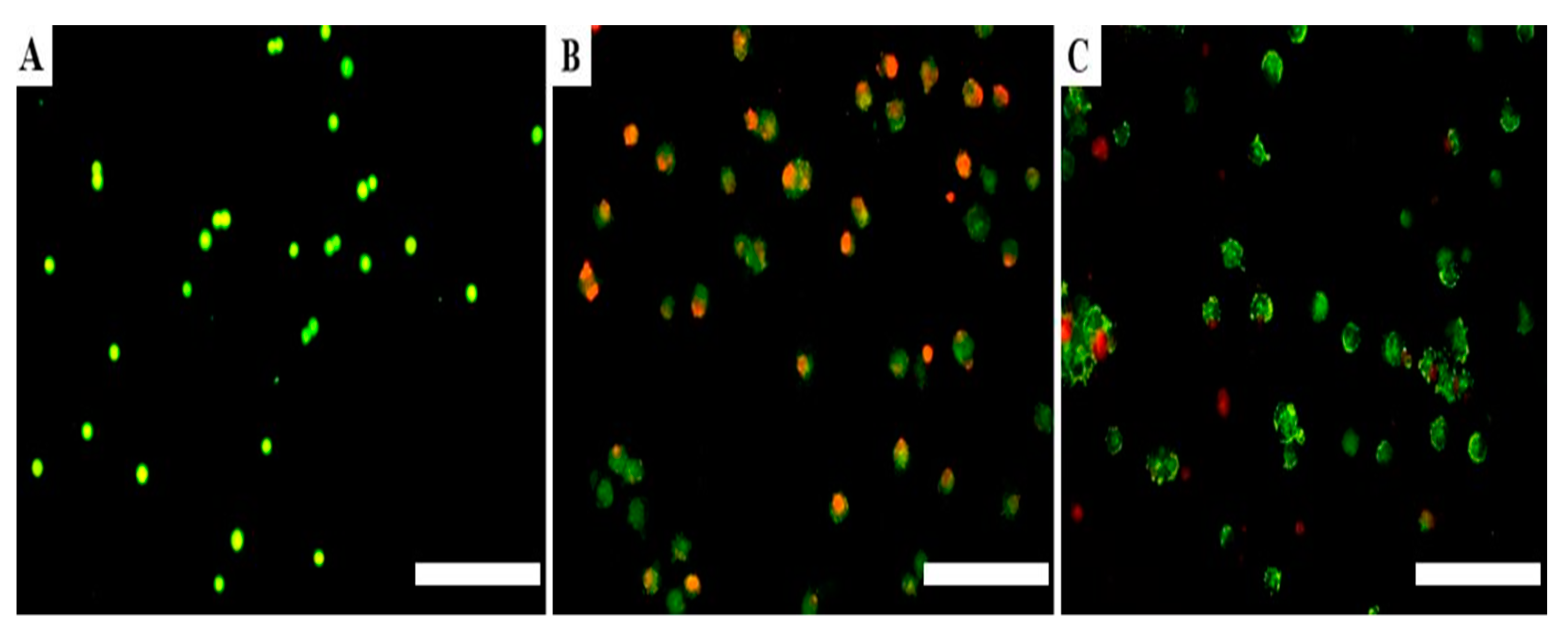
3.3. Treatment Triggered Apoptosis in Adenocarcinoma Lung Cells Compared to Non-Transfected Cells
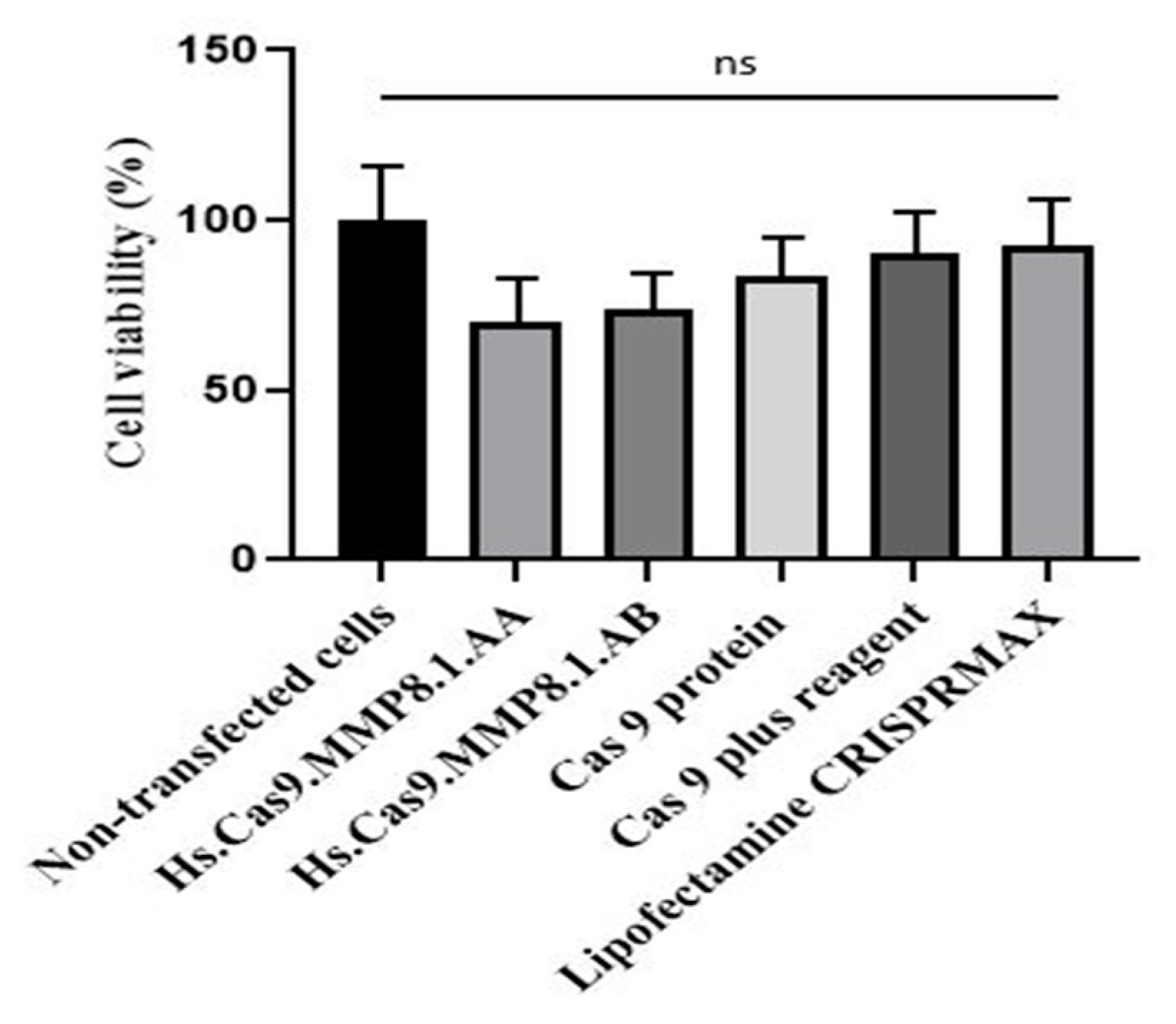
3.4. Delivery of Transfection Components Did Not Affect the Cell Viability
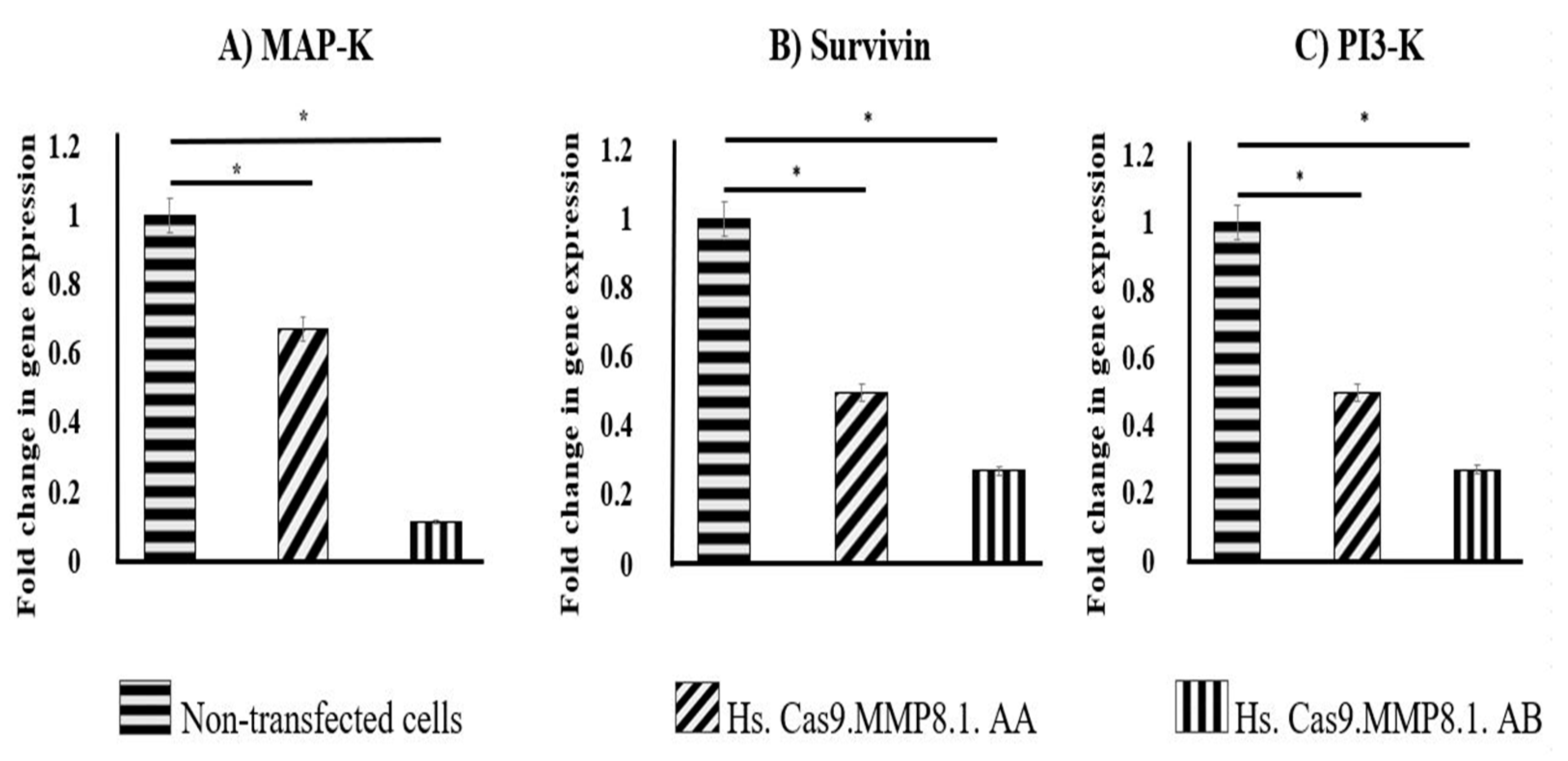
3.5. Lipofection of sgRNAs and Cas9 Protein Complexes Resulted in the Decreased Expression of MAP-K, Survivin and PI3-K in A549 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rich, A.L.; Khakwani, A.; Free, C.M.; Tata, L.J.; Stanley, R.A.; Peake, M.D.; Hubbard, R.B.; Baldwin, D.R. Non-small cell lung cancer in young adults: Presentation and survival in the English National Lung Cancer Audit. QJM Int. J. Med. 2015, 108, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Roh, M.S. Molecular pathology of lung cancer: Current status and future directions. Tuberc. Respir. Dis. 2014, 77, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Santini, D.; Barni, S.; Intagliata, S.; Falcone, A.; Ferraù, F.; Galetta, D.; Moscetti, L.; La Verde, N.; Ibrahim, T.; Petrelli, F. Natural history of non-small-cell lung cancer with bone metastases. Sci. Rep. 2015, 5, 18670. [Google Scholar] [CrossRef] [Green Version]
- Riihimäki, M.; Hemminki, A.; Fallah, M.; Thomsen, H.; Sundquist, K.; Sundquist, J.; Hemminki, K. Metastatic sites and survival in lung cancer. Lung Cancer 2014, 86, 78–84. [Google Scholar] [CrossRef] [PubMed]
- dos Reis Oliveira, M.B.; de Queiroz Mello, F.C.; Paschoal, M.E.M. The relationship between lung cancer histology and the clinicopathological characteristics of bone metastases. Lung Cancer 2016, 96, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Lee, H.; Um, S.-W.; Kim, K.; Zo, J.I.; Shim, Y.M.; Kwon, O.J.; Lee, K.S.; Ahn, M.-J.; Kim, H. Incidence of brain metastasis in lung adenocarcinoma at initial diagnosis on the basis of stage and genetic alterations. Lung Cancer 2019, 129, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Radisky, E.S.; Radisky, D.C. Matrix metalloproteinase-induced epithelial-mesenchymal transition in breast cancer. J. Mammary Gland. Biol. Neoplasia 2010, 15, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.E.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez Sanchez, M.E. Role of matrix metalloproteinases in angiogenesis and cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.F.; Zhu, L.J.; Zhang, W.; Yuan, W.; Song, N.H.; Zuo, L.; Mi, Y.Y.; Wang, Z.J. MMP-8 C-799 T, Lys460Thr, and Lys87Glu variants are not related to risk of cancer. BMC Med. Genet. 2019, 20, 162. [Google Scholar] [CrossRef]
- Juurikka, K.; Butler, G.S.; Salo, T.; Nyberg, P.; Åström, P. The role of MMP8 in cancer: A systematic review. Int. J. Mol. Sci. 2019, 20, 4506. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.A.; Spinale, F.G.; Ikonomidis, J.S.; Stroud, R.E.; Chang, E.I.; Reed, C.E. Differential matrix metalloproteinase levels in adenocarcinoma and squamous cell carcinoma of the lung. J. Thorac. Cardiovasc. Surg. 2010, 139, 984–990. [Google Scholar] [CrossRef] [Green Version]
- Ramanujum, R.; Lin, Y.L.; Liu, J.K.; He, S. Regulatory expression of MMP-8/MMP-9 and inhibition of proliferation, migration and invasion in human lung cancer A549 cells in the presence of HGF variants. Kaohsiung J. Med. Sci. 2013, 29, 530–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, T.C.; Hsia, T.C.; Chao, C.-Y.; Chen, W.-C.; Chen, C.-Y.; Chen, W.-C.; Lin, Y.-T.; Hsiao, C.-L.; Chang, W.-S.; Tsai, C.-W. The contribution of MMP-8 promoter polymorphisms in lung cancer. Anticancer. Res. 2017, 37, 3563–3567. [Google Scholar]
- Saito, R.; Miki, Y.; Ishida, N.; Inoue, C.; Kobayashi, M.; Hata, S.; Yamada-Okabe, H.; Okada, Y.; Sasano, H. The Significance of MMP-1 in EGFR-TKI–Resistant Lung Adenocarcinoma: Potential for Therapeutic Targeting. Int. J. Mol. Sci. 2018, 19, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Zhu, H.; Li, Y. PKCζ, MMP-2 and MMP-9 expression in lung adenocarcinoma and association with a metastatic phenotype. Mol. Med. Rep. 2017, 16, 8301–8306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, Y.W.; Chien, S.T.; Chen, P.S.; Lee, J.H.; Wu, S.H.; Yin, L.T. α-Mangostin Suppresses Phorbol 12-myristate 13-acetate-Induced MMP-2/MMP-9 Expressions via αvβ3 Integrin/FAK/ERK and NF-κB Signaling Pathway in Human Lung Adenocarcinoma A549 Cells. Cell Biochem. Biophys. 2010, 58, 31–44. [Google Scholar] [CrossRef]
- Mok, P.L.; Arun, N.K.A.; Hernandez, M.O.D.; Tong, J.; Aisha, F.; Mohammed Safwan, A.K.; Gothai, S.; Avin, K.E.H.; Badr, A. Lung development, repair and cancer: A study on the role of MMP20 Gene in Adenocarcinoma. PLoS ONE 2021, 16, e0250552. [Google Scholar] [CrossRef]
- Ryan, D.E.; Taussig, D.; Steinfeld, I.; Phadnis, S.M.; Lunstad, B.D.; Singh, M.; Vuong, X.; Okochi, K.D.; McCaffrey, R.; Olesiak, M.; et al. Improving CRISPR–Cas specificity with chemical modifications in single-guide RNAs. Nucleic Acids Res. 2017, 46, 792–803. [Google Scholar] [CrossRef]
- Cromwell, C.R.; Sung, K.; Park, J.; Krysler, A.R.; Jovel, J.; Kim, S.K.; Hubbard, B.P. Incorporation of bridged nucleic acids into CRISPR RNAs improves Cas9 endonuclease specificity. Nat. Commun. 2018, 9, 1448. [Google Scholar] [CrossRef]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research techniques made simple: Analysis of collective cell migration using the wound healing assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef] [Green Version]
- Craig, V.J.; Zhang, L.; Hagood, J.S.; Owen, C.A. Matrix metalloproteinases as therapeutic targets for idiopathic pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 2015, 53, 585–600. [Google Scholar] [CrossRef] [Green Version]
- Manicone, A.M.; McGuire, J.K. Matrix metalloproteinases as modulators of inflammation. Semin. Cell Dev. Biol. 2008, 19, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardomi, A.; Abediankenari, S. Matrix metalloproteinase 8: Could it benefit the CAR-T cell therapy of solid tumors?-a-Commentary on therapeutic potential. Cancer Microenviron. 2018, 11, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Fernández, A.; Fueyo, A.; Folgueras, A.R.; Garabaya, C.; Pennington, C.J.; Pilgrim, S.; Edwards, D.R.; Holliday, D.L.; Jones, J.L.; Span, P.N. Matrix metalloproteinase-8 functions as a metastasis suppressor through modulation of tumor cell adhesion and invasion. Cancer Res. 2008, 68, 2755–2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plosa, E.J.; Benjamin, J.T.; Sucre, J.M.; Gulleman, P.M.; Gleaves, L.A.; Han, W.; Kook, S.; Polosukhin, V.V.; Haake, S.M.; Guttentag, S.H.; et al. Beta1 Integrin regulates adult lung alveolar epithelial cell inflammation. JCI Insight 2020, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.B.; Nabha, S.M.; Atanaskova, N. Role of MAP kinase in tumor progression and invasion. Cancer Metastasis Rev. 2003, 22, 395–403. [Google Scholar] [CrossRef]
- Madsen, R.R. PI3K in stemness regulation: From development to cancer. Biochem. Soc. Trans. 2020, 48, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Bronger, H.; Magdolen, V.; Goettig, P.; Dreyer, T. Proteolytic chemokine cleavage as a regulator of lymphocytic infiltration in solid tumors. Cancer Metastasis Rev. 2019, 38, 417–430. [Google Scholar] [CrossRef] [Green Version]
- Farahani, E.; Patra, H.K.; Jangamreddy, J.R.; Rashedi, I.; Kawalec, M.; Rao Pariti, R.K.; Batakis, P.; Wiechec, E. Cell adhesion molecules and their relation to (cancer) cell stemness. Carcinogenesis 2014, 35, 747–759. [Google Scholar] [CrossRef] [Green Version]
- Devarasetty, M.; Forsythe, S.D.; Shelkey, E.; Soker, S. In Vitro Modeling of the Tumor Microenvironment in Tumor Organoids. Tissue Eng. Regen. Med. 2020, 17, 759–771. [Google Scholar] [CrossRef]
- Owen, C.A.; Hu, Z.; Lopez-Otin, C.; Shapiro, S.D. Membrane-bound matrix metalloproteinase-8 on activated polymorphonuclear cells is a potent, tissue inhibitor of metalloproteinase-resistant collagenase and serpinase. J. Immunol. 2004, 172, 7791–7803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Kim, C.H.; An, B.; Ha, K.S.; Hong, S.H.; Kim, K.S. Effects of clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR-associated (Cas) protein 9 system-Based Deletion of miR-451 in Mouse Embryonic Stem Cells on Their Self-Renewal and Hematopoietic Differentiation. Tissue Eng. Regen. Med. 2017, 14, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.R.; Collins, K.H.; Lee, J.W.; Kang, H.J.; Guilak, F. Genome engineering for osteoarthritis: From designer cells to disease-modifying drugs. Tissue Eng. Regen. Med. 2019, 16, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Im, G.I.; Cho, C.S. Updates in Cartilage Tissue Regeneration. Tissue Eng. Regen. Med. 2019, 16, 325–326. [Google Scholar] [CrossRef] [Green Version]
- Pryzhkova, M.V.; Jordan, P.W. Adaptation of Human Testicular Niche Cells for Pluripotent Stem Cell and Testis Development Research. Tissue Eng. Regen. Med. 2020, 17, 223–235. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maradiaga, O.D.H.; Mok, P.L.; Sivapragasam, G.; Samrot, A.V.; Ali Khan, M.S.; Farhana, A.; Alzahrani, B.; Tong, J.; Karuppiah, T.; Joseph, N.M.S.; et al. Lipofection of Single Guide RNA Targeting MMP8 Decreases Proliferation and Migration in Lung Adenocarcinoma Cells. Medicina 2021, 57, 710. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57070710
Maradiaga ODH, Mok PL, Sivapragasam G, Samrot AV, Ali Khan MS, Farhana A, Alzahrani B, Tong J, Karuppiah T, Joseph NMS, et al. Lipofection of Single Guide RNA Targeting MMP8 Decreases Proliferation and Migration in Lung Adenocarcinoma Cells. Medicina. 2021; 57(7):710. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57070710
Chicago/Turabian StyleMaradiaga, Oscar David Hernandez, Pooi Ling Mok, Gothai Sivapragasam, Antony V. Samrot, Mohammed Safwan Ali Khan, Aisha Farhana, Badr Alzahrani, Jiabei Tong, Thilakavathy Karuppiah, Narcisse M. S. Joseph, and et al. 2021. "Lipofection of Single Guide RNA Targeting MMP8 Decreases Proliferation and Migration in Lung Adenocarcinoma Cells" Medicina 57, no. 7: 710. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57070710