Characterization of MK8(H2) from Rhodococcus sp. B7740 and Its Potential Antiglycation Capacity Measurements


Abstract
:1. Introduction
2. Results and Discussion
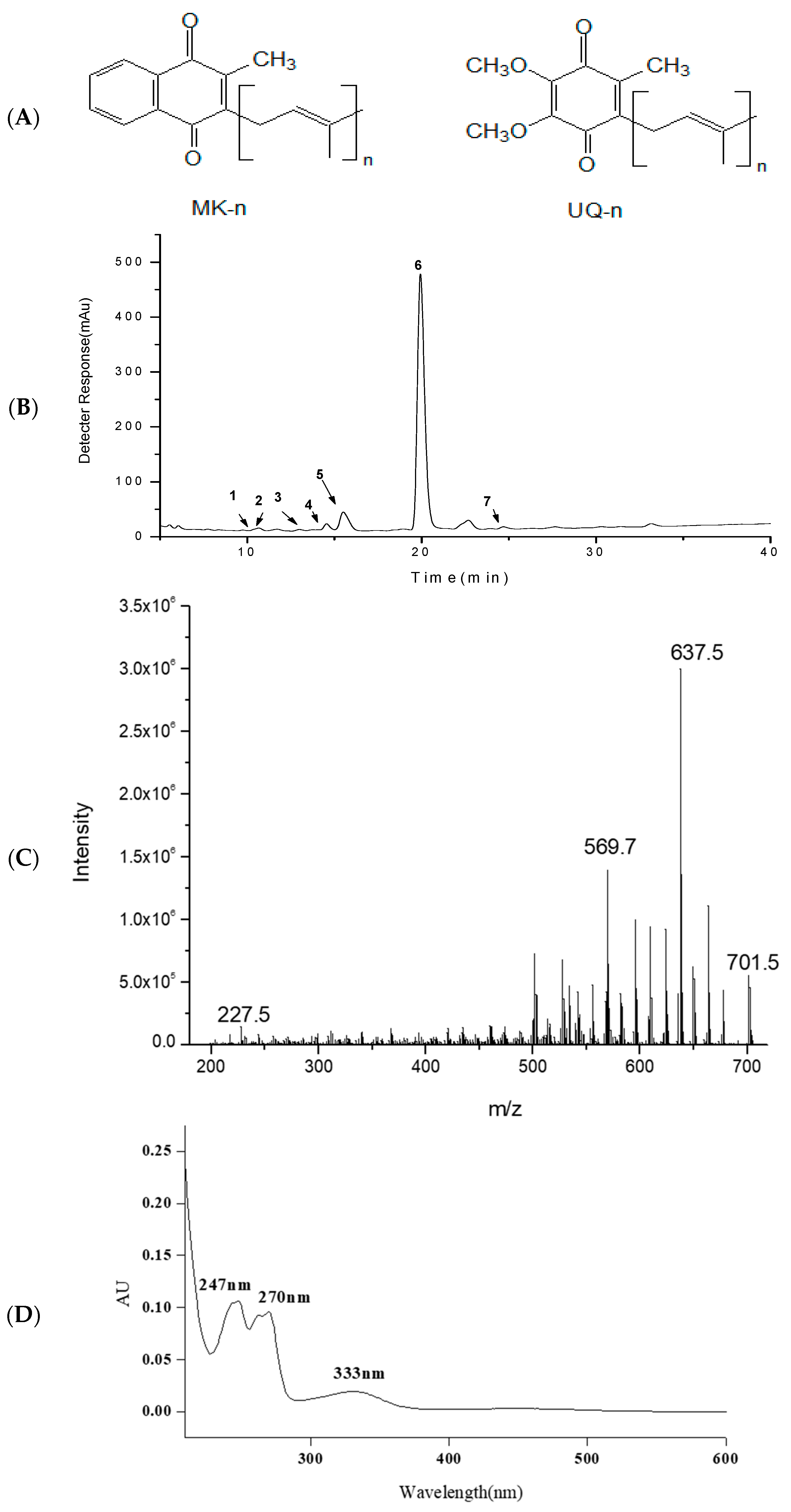
2.1. HPLC-APCI-MS/MS Analysis of MKs
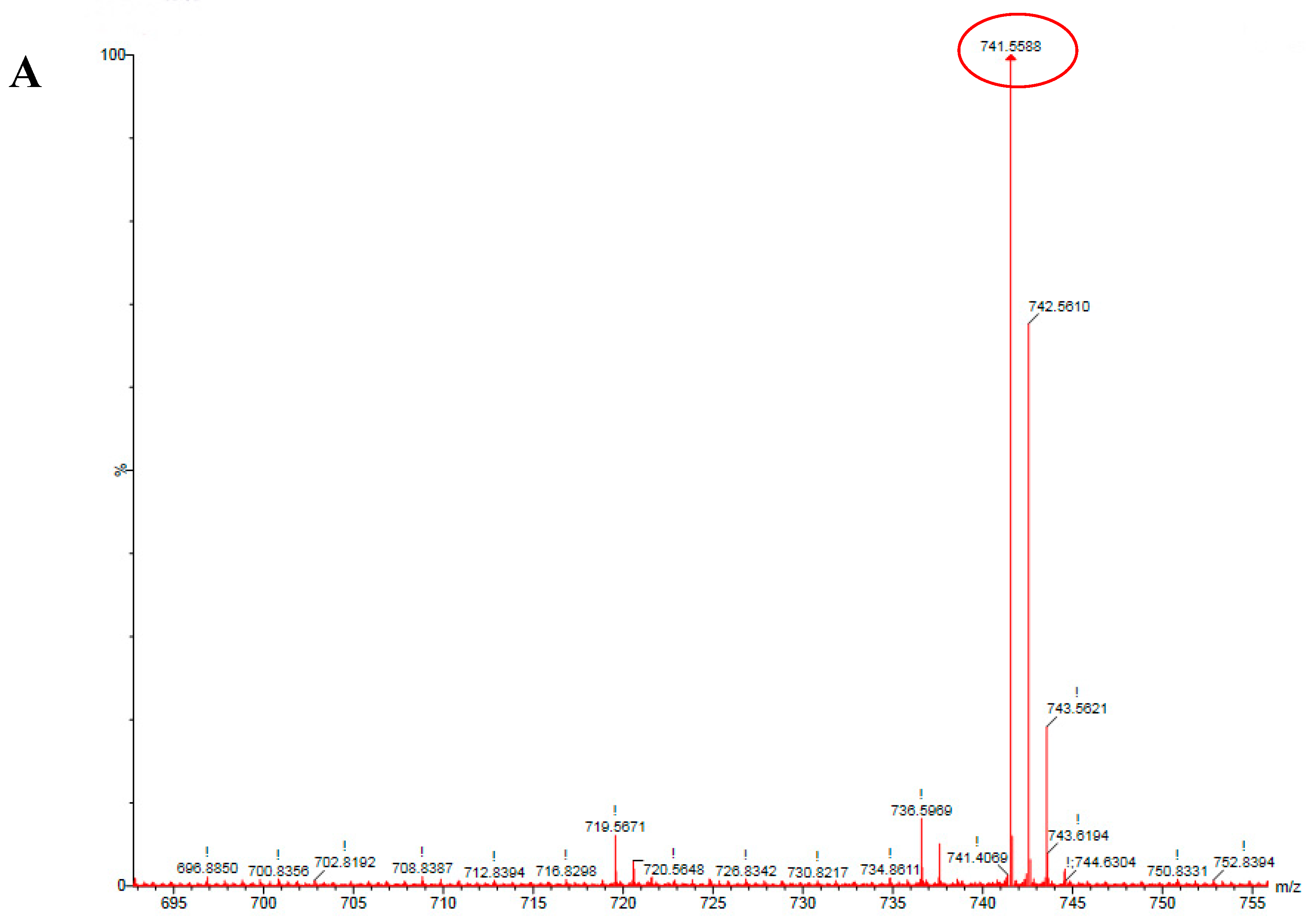
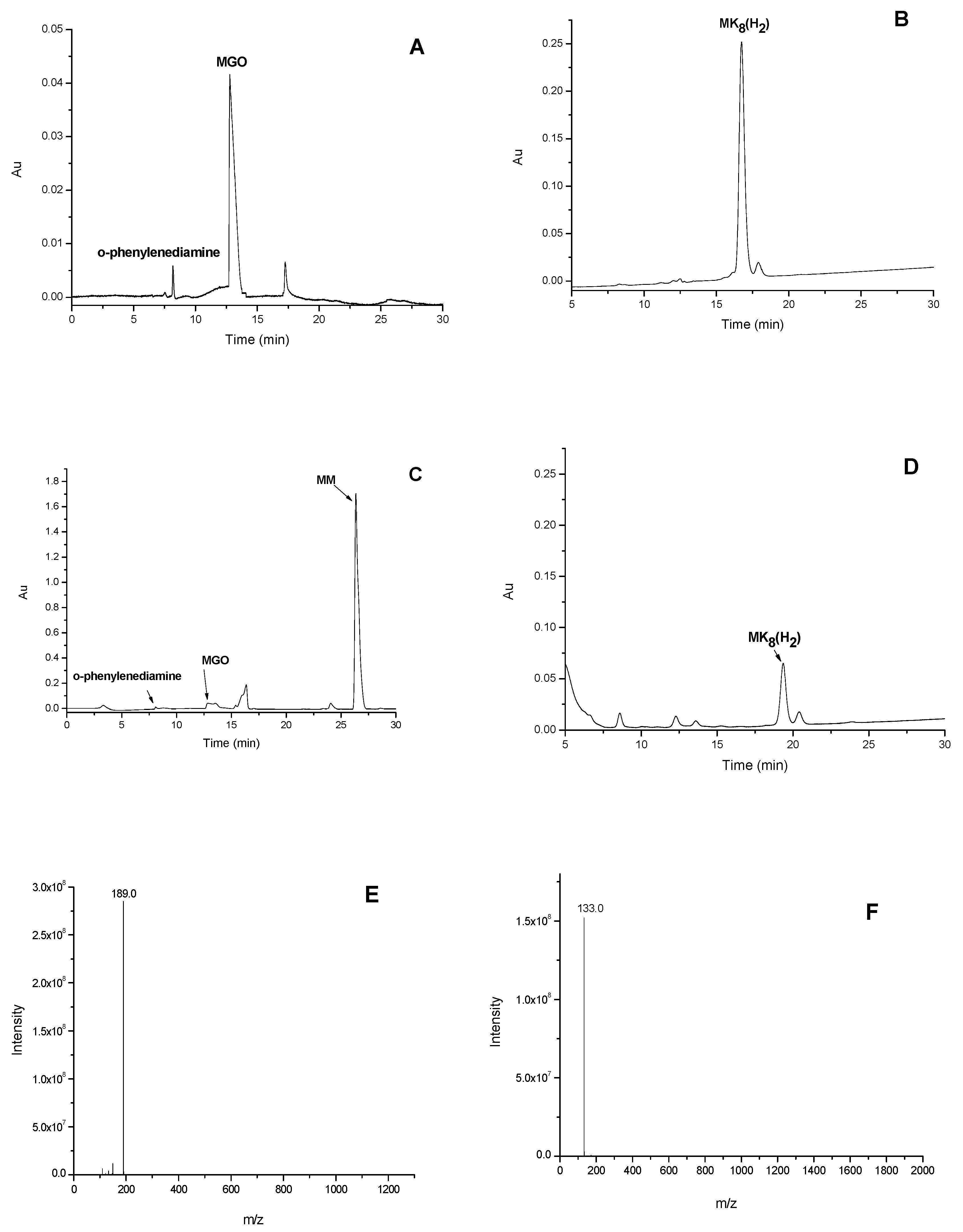
2.2. HSCCC Purification and URMS (Ultra-High Mass Spectrum) Analysis of MK8(H2)
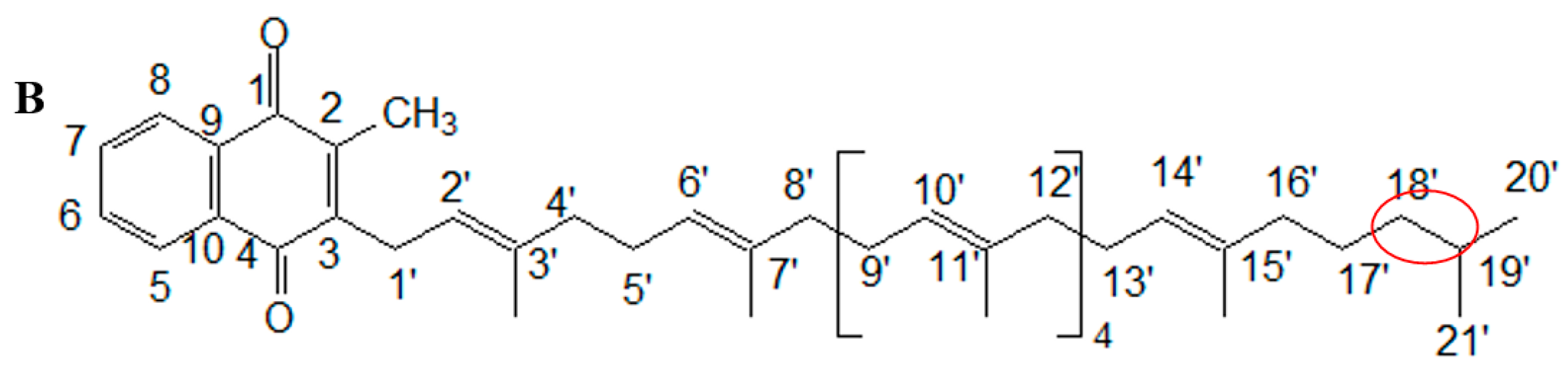
2.3. NMR Analysis of MK8(H2)
2.4. Antioxidative Effect and Antiglycation Capability
2.5. Analysis of MK8(H2)-Carbonyl Adducts by LC-MS/MS
2.6. Docking Studies
3. Materials and Methods
3.1. Extraction of MKs
3.2. LC-MS/MS Analyses of MKs
3.3. HSCCC for Purification of MKs
3.4. UPLC-ESI-QTOF-HRMS Analysis of MKs
3.5. NMR Analysis of MKs
3.6. Determination of Antioxidative Effect and Antiglycation Capability
3.7. Determination of MK8(H2)-Carbonyl Adducts by LC-MS/MS
3.8. Molecular Docking
3.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Makoto, K. Biosynthesis and applications of prenylquinones. Biosci. Biotechnol. Biochem. 2018, 82, 963–977. [Google Scholar]
- Suttie, J.W. Mechanism of action of vitamin K: Synthesis of gamma-carboxyglutamic acid. Crit. Rev. Biochem. 1980, 8, 191–233. [Google Scholar] [CrossRef]
- Tsaioun, K.I. Vitamin K-dependent Proteins in the Developing and Aging Nervous System. Nutr. Rev. 1999, 57, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Littarru, G.P.; Tiano, L. Clinical aspects of coenzyme Q10: An update. Nutrition 2010, 26, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Kurosu, M.; Begari, E. Vitamin K2 in electron transport system: Are enzymes involved in vitamin K2 biosynthesis promising drug targets? Molecules 2010, 15, 1531–1553. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, Y.; Ichise, H.; Kakuda, H.; Yamaguchi, M. Intake of fermented soybean (natto) increases circulating vitamin K2 (menaquinone-7). J. Bone Miner. Metab. 2000, 18, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Taguchi, H.; Gao, Y.H.; Igarashi, A.; Tsukamoto, Y. Effect of vitamin K2 (menaquinone-7) in fermented soybean (natto) on bone loss in ovariectomized. J. Am. Soc. Hypertens. 1999, 17, 23–29. [Google Scholar]
- Mandatori, D.; Penolazzi, L.; Pipino, C.; Di Tomo, P.; Di Silvestre, S.; Di Pietro, N.; Trevisani, S.; Angelozzi, M.; Ucci, M.; Piva, R.; et al. Menaquinone-4 enhances osteogenic potential of human amniotic fluid mesenchymal stem cells cultured in 2D and 3D dynamic culture systems. J. Tissue Eng. Regen. Med. 2018, 12, 447–459. [Google Scholar] [CrossRef] [PubMed]
- McFarlin, B.K.; Henning, A.L.; Venable, A.S. Oral Consumption of Vitamin K2 for 8 Weeks Associated with Increased Maximal Cardiac Output During Exercise. Altern. Ther. Health Med. 2017, 23, 26–32. [Google Scholar]
- Mansour, A.G.; Hariri, E.; Daaboul, Y.; Korjian, S.; El Alam, A.; Protogerou, A.D.; Kilany, H.; Karam, A.; Stephan, A.; Bahous, S.A. Vitamin K2 supplementation and arterial stiffness among renal transplant recipients-a single-arm, single-center clinical trial. J. Am. Soc. Hypertens. 2017, 11, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Vossen, L.M.; Schurgers, L.J.; van Varik, B.J.; Kietselaer, B.L.; Vermeer, C.; Meeder, J.G.; Rahel, B.M.; van Cauteren, Y.J.; Hoffland, G.A.; Rennenberg, R.J.; et al. Menaquinone-7 supplementation to reduce vascular calcification in patients with coronary artery disease: Rationale and study Protocol (VitaK-CAC Trial). Nutrients 2015, 7, 8905–8915. [Google Scholar] [CrossRef] [PubMed]
- Janssen, R.; Vermeer, C. Vitamin K deficit and elastolysis theory in pulmonary elasto-degenerative diseases. Med. Hypotheses 2017, 108, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Noda, S.; Yamada, A.; Tanabe, R.; Nakaoka, K.; Hosoi, T.; Goseki-Sone, M. Menaquinone-4 (vitamin K2) up-regulates expression of human intestinal alkaline phosphatase in Caco-2 cells. Nutr. Res. 2016, 36, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Sheetal, B.; Thomas, R.C. Vitamin K2 takes charge. Science 2012, 336, 1241–1242. [Google Scholar]
- Meganathan, R. Biosynthesis of Menaquinone (Vitamin K2) and Ubiquinone (Coenzyme Q): A Perspective on Enzymatic Mechanisms. Vitam. Horm. 2001, 61, 173–218. [Google Scholar] [PubMed]
- Haruo, S.Y.J.; Tomoshige, H.; Miwako, F.; Kazuo, F.N.I.; Tohru, D. Studies on a New Biosynthetic Pathway for Menaquinone. J. Am. Chem. Soc. 2008, 130, 5614–5615. [Google Scholar]
- Nowicka, B.; Kruk, J. Occurrence, biosynthesis and function of isoprenoid quinones. Biochim. Biophys. Acta 2010, 1797, 1587–1605. [Google Scholar] [CrossRef] [PubMed]
- Berenjian, A.; Mahanama, R.; Talbot, A.; Regtop, H.; Kavanagh, J.; Dehghani, F. Designing of an intensification process for biosynthesis and recovery of menaquinone-7. Appl. Biochem. Biotechnol. 2014, 172, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Min, K.K.; Pyung, C.L. Metabolic Engineering of Menaquinone-8 Pathway of Escherichia coli as a Microbial Platform for Vitamin K Production. Biotechnol. Bioeng. 2011, 108, 1997–2002. [Google Scholar]
- Chen, Y.; Xie, B.; Yang, J.; Chen, J.; Sun, Z. Identification of microbial carotenoids and isoprenoid quinones from Rhodococcus sp. B7740 and its stability in the presence of iron in model gastric conditions. Food Chem. 2018, 240, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Gast, G.C.M.; de Roos, N.M.; Sluijs, I.; Bots, M.L.; Beulens, J.W.; Geleijnse, J.M.; Witteman, J.C.; Grobbee, D.E.; Peeters, P.H.; van der Schouw, Y.T. A high menaquinone intake reduces the incidence of coronary heart disease. Nutr. Metab. Cardiovasc. 2009, 19, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Degli, E.M. A journey across genomes uncovers the origin of ubiquinone in cyanobacteria. Genome Biol. Evol. 2017, 9, 3039–3053. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, P.; Geyer, R.; Surmann, P.; Fuhrmann, H. LC-MS method for screening unknown microbial carotenoids and isoprenoid quinones. J. Microbiol. Methods 2012, 88, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, L.; Zhao, G.; Fang, Z.; Wu, H.; Wang, P.; Zheng, Z. Extraction, purification and identification of menaquinones from Flavobacterium meningosepticum fermentation medium. Process. Biochem. 2018, 66, 245–253. [Google Scholar] [CrossRef]
- Mesías, M.; Navarro, M.; Martínez-Saez, N.; Ullate, M.; del Castillo, M.D.; Morales, F.J. Antiglycative and carbonyl trapping properties of the water soluble fraction of coffee silverskin. Food. Res. Int. 2014, 62, 1120–1126. [Google Scholar] [CrossRef] [Green Version]
- Harris, C.S.; Beaulieu, L.P.; Fraser, M.H.; McIntyre, K.L.; Owen, P.L.; Martineau, L.C. Inhibition of advanced glycation end product formation by medicinal plant extracts correlates with phenolic metabolites and antioxidant activity. Planta Med. 2011, 77, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Nattha, J.; Sirintorn, Y.; Sirichai, A. Inhibition of advanced glycation end products by red grape skin extract and its antioxidant. BMC Complement. Altern. Med. 2013, 13, 171–179. [Google Scholar]
- Muhammad, Z.; Shakeel, A.; Shazia, A.B.; Ryszard, A.; Sezai, E.; Hawa, Z.E.J. Compositional studies and biological activities of some mash bean (Vigna mungo (L.) Hepper) cultivars commonly consumed in Pakistan. Biol. Res. 2014, 47, 23–37. [Google Scholar]
- Sadowska-Bartosz, I.; Bartosz, G. Prevention of protein glycation by natural compounds. Molecules 2015, 20, 3309–3334. [Google Scholar] [CrossRef] [PubMed]
- Okano, T.; Shimomura, Y.; Yamane, M.; Suhara, Y.; Kamao, M.; Sugiura, M.; Nakagawa, K. Conversion of phylloquinone (Vitamin K1) into menaquinone-4 (Vitamin K2) in mice: Two possible routes for menaquinone-4 accumulation in cerebra of mice. J. Biol. Chem. 2008, 283, 11270–11279. [Google Scholar] [CrossRef] [PubMed]
- Eugeäne, E.N.; Sylvain, S.; Khaled, B. Anthocyanins in wild blueberries of Quebec: Extraction and identification. J. Agric. Food Chem. 2007, 55, 5626–5635. [Google Scholar]
- Wang, W.; Yagiz, Y.; Buran, T.J.; Nunes, C.d.N.; Gu, L. Phytochemicals from berries and grapes inhibited the formation of advanced glycation end-products by scavenging reactive carbonyls. Food Res. Int. 2011, 44, 2666–2673. [Google Scholar] [CrossRef]
- Shen, Y.; Xu, Z.; Sheng, Z. Ability of resveratrol to inhibit advanced glycation end product formation and carbohydrate-hydrolyzing enzyme activity, and to conjugate methylglyoxal. Food Chem. 2017, 216, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Kiran, B.U.; Khan, B.A.A.; Arunachalam, J.; Anhazhagan, V. Spectrofluorimetric and molecular docking investigation on the interaction of 6-azauridine, a pyrimidine nucleoside antimetabolite, with serum protein. J. Mol. Liq. 2016, 219, 602–607. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
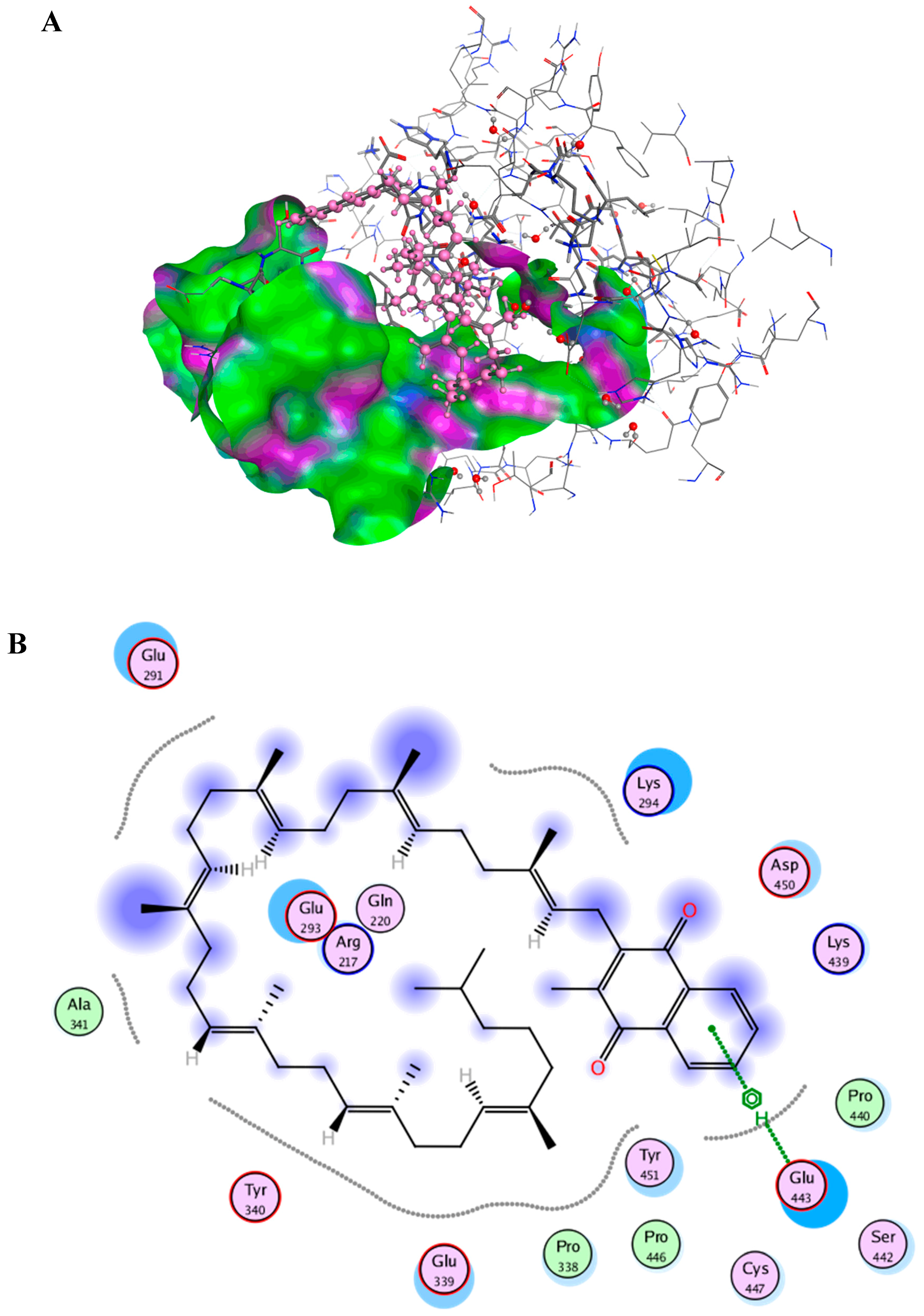{kind=link}
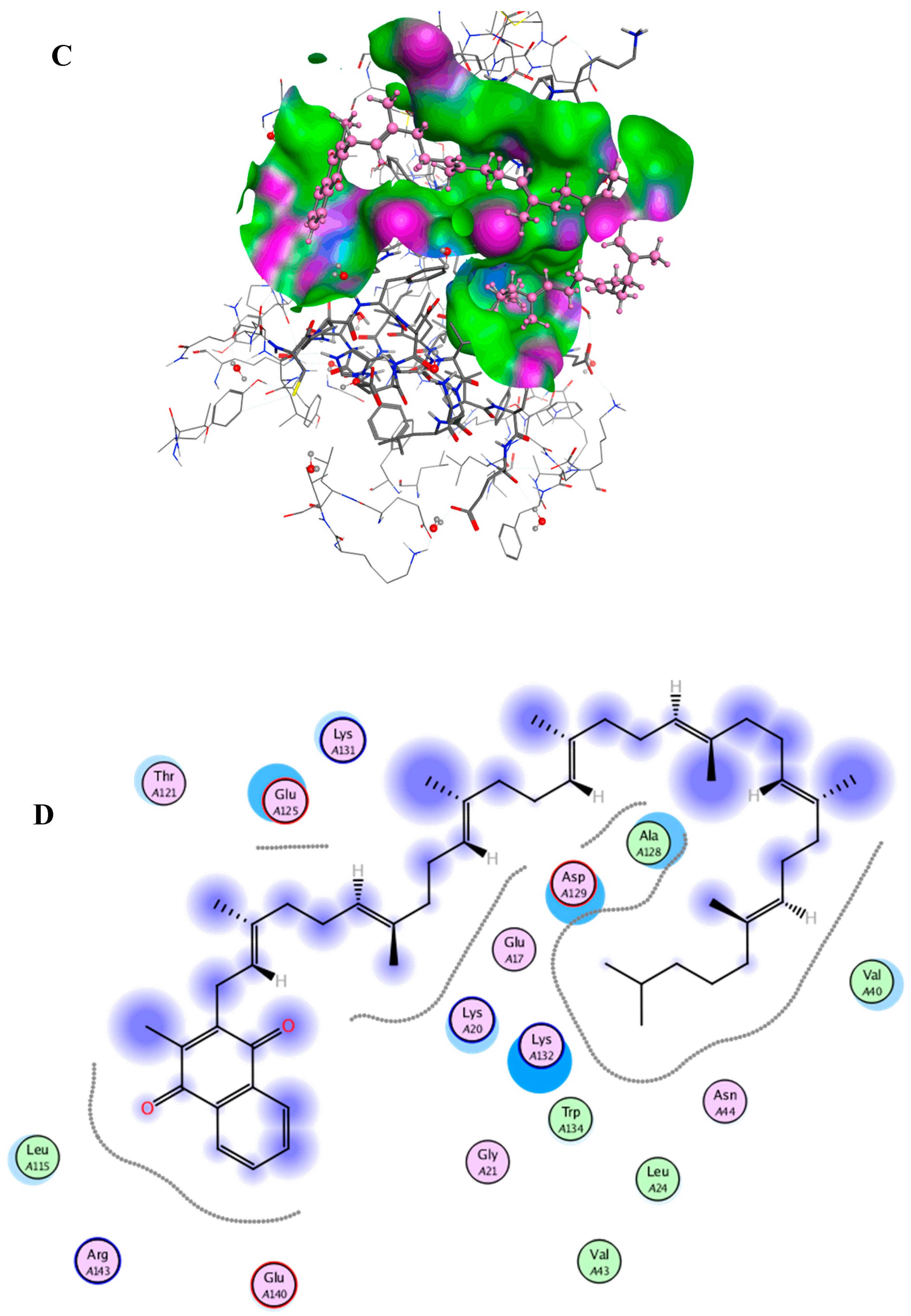{kind=link}
| Peak a | Compound b | Formula | tR (min) | λmax (nm) c | [M + H]+ (m/z) | MS/MS (m/z) |
|---|---|---|---|---|---|---|
| 1 | MK8(H2) | C51H74O2 | 10.2 | 250,264 | 719 | 701,675,227, |
| 2 | MK8 | C51H72O2 | 10.4 | 248,262 | 717 | 699,567,227 |
| 3 | MK8(H4) | C51H76O2 | 13.0 | 252 | 721 | 703,571,227 |
| 4 | MK8(H6) | C51H78O2 | 14.4 | 252 | 723 | 705,571,227 |
| 5 | MK7(H2) | C46H66O2 | 15.4 | 243,268 | 651 | 633,567,187 |
| 6 | MK8(H2) | C51H74O2 | 19.7 | 247,269 | 719 | 701,635,227 |
| 7 | MK9(H4) | C56H84O2 | 24.8 | 223,269 | 789 | 770,637,501 |
| C/H Position | 1H | 13C | C/H Position | 1H | 13C |
|---|---|---|---|---|---|
| 1 | 185.40 | 11′ | 124.28 | ||
| 2 | 143.43 | 12′ | 3.60–3.75 (m) | 35.63 | |
| 3 | 146.05 | 13′ | 3.60–3.75 (m) | 32.31 | |
| 4 | 184.50 | 14′ | 5.00 (t) | 124.27 | |
| 5 | 8.08 (q) | 126.20 | 15′ | 118.84 | |
| 6 | 7.68 (q) | 134.87 | 16′ | 3.60–3.75 (m) | 31.92 |
| 7 | 7.67 (q) | 133.34 | 17′ | 0.91–1.20(m) | 29.60 |
| 8 | 8.07 (q) | 126.12 | 18′ | 0.91–1.20(m) | 29.43 |
| 9 | 131.25 | 19′ | 1.50–2.09 (m) | 29.38 | |
| 10 | 132.17 | 20′ | 0.82–0.89 (m) | 19.60 | |
| 1′ | 3.38 (d) | 70.7 | 21′ | 0.82–0.89 (m) | 14.20 |
| 2′ | 5.34 (t) | 129.35 | 2-CH3 | 2.19 (s) | 29.72 |
| 3′ | 129.85 | 3′-CH3 | 0.90–1.48 (m) | 29.34 | |
| 4′ | 3.60–3.75 (m) | 40.04 | 7′-CH3 | 27.25 | |
| 5′ | 3.60–3.75 (m) | 39.77 | 12′-CH3 | 26.71 | |
| 6′ | 5.08–5.12 (m) | 126.20 | 12′-CH3 | 26.02 | |
| 7′ | 124.85 | 12′-CH3 | 25.71 | ||
| 8′ | 3.60–3.75 (m) | 37.09 | 12′-CH3 | 25.47 | |
| 9′ | 3.60–3.75 (m) | 36.56 | 16′-CH3 | 25.26 | |
| 10′ | 5.08–5.12 (m) | 124.42 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Mu, Q.; Hu, K.; Chen, M.; Yang, J.; Chen, J.; Xie, B.; Sun, Z. Characterization of MK8(H2) from Rhodococcus sp. B7740 and Its Potential Antiglycation Capacity Measurements. Mar. Drugs 2018, 16, 391. https://0-doi-org.brum.beds.ac.uk/10.3390/md16100391
Chen Y, Mu Q, Hu K, Chen M, Yang J, Chen J, Xie B, Sun Z. Characterization of MK8(H2) from Rhodococcus sp. B7740 and Its Potential Antiglycation Capacity Measurements. Marine Drugs. 2018; 16(10):391. https://0-doi-org.brum.beds.ac.uk/10.3390/md16100391
Chicago/Turabian StyleChen, Yashu, Qin Mu, Kai Hu, Mo Chen, Jifang Yang, Jigang Chen, Bijun Xie, and Zhida Sun. 2018. "Characterization of MK8(H2) from Rhodococcus sp. B7740 and Its Potential Antiglycation Capacity Measurements" Marine Drugs 16, no. 10: 391. https://0-doi-org.brum.beds.ac.uk/10.3390/md16100391