1. Introduction
Hepatocellular carcinoma (HCC) is the most common liver cancer, accounting for approximately 90% [
1,
2], and it has a five-year survival rate around only 30–40% [
3]. Similar to other kinds of cancers, tumor metastasis remains the primary cause of mortality of HCC patients [
4]. It is well known that metastasis is a multistage process that requires cancer cell detachment from the primary tumor, survival in the circulation, settlement at distant sites and growth [
5]. During this process, metastatic cells go through detachment, migration, invasion and adhesion, and inhibition of any one of these steps has the potential to block the entire metastatic process [
6]. However, there is very little advancement in the area of blocking tumor metastasis [
6], therefore, new effective agents possessing anti-metastasis activities are urgently needed for better prevention and treatment of HCC and other cancers.
Recently, marine microbial polysaccharide has become an important resource for searching anti-metastasis agents because they possess numerous advantages including biocompatibility, biodegradability, nontoxicity, low cost and abundance [
7]. Heymann and colleagues found that a marine bacterial exopolysaccharide, OS-EPS, was potent in inhibiting both migration and invasiveness of osteosarcoma cell lines and was very efficient in inhibiting the establishment of lung metastases in vivo [
8]. In our previous study, we found that a novel marine bacterial polysaccharide, EPS11, showed strong cytotoxic effects on lung cancer cells through affecting cell adhesion and anoikis [
9]. Notably, filiform structures of A549 cells were markedly destroyed by EPS11 in a dose-dependent manner [
9]. Filiform structures include filopodia, invadopodia, filopodium-like protrusions and actin spikes [
10]. There is increasing evidence showing that cancer cells use filiform structures, in cell adhesion and three-dimensional cell migration, both in vitro and in vivo [
10]. Metastatic tumors cells are rich in filiform structures, and the number of which correlates with their invasiveness [
11]. Recently, filiform structures have also been implicated as critical for metastatic tumors cells to colonize in secondary tissues or organs [
12]. Hence, the formation of filiform structures represents a critical rate-limiting step for the subsequent development of macroscopic metastases and becomes a hot target for anti-cancer drug development.
Meanwhile, lots of studies have stated that many molecular factors are involved in the process of HCC metastasis, including adhesion molecules, matrix metalloproteinases, vascular growth factors, oncogenes, etc. [
13]. CD99, a glycosylated transmembrane protein, has been reported to have a marked effect on the migration, invasion, and metastasis of tumor cells through multiple and unclear action mechanisms, thereby emerging as a novel therapeutic target [
14]. However, much still needs to be understood. Nevertheless, several studies indicated that agonistic CD99 monoclonal antibodies were able to activate cell death signals and inhibit cell migration [
14]. Furthermore, CD99 is positively stained in immunohistochemical preparations of HCC tissue material and negatively stained in non-HCC tissues outside the liver or metastatic within the liver [
15], which strongly suggests that CD99 is essential in the course of cancer development and metastasis of HCC.
Based on our previous results, EPS11 preferentially killed cancer cells, including human lung cancer cells, A549, and HCV-related human liver cancer cells, Huh7.5, compared with normal cell line human embryonic lung fibroblasts, WI-38, and potentially possessed anti-metastasis activity [
9]. However, the detailed molecular mechanisms of EPS11 against liver cancer and its potential anti-metastasis activity are not clear. In this work, we first investigated the suppression of EPS11 on Huh7.5 cells’ filiform structure formation, adhesion and migration abilities. Then we chose CD99 as a reference for anti-cancer mechanism studies of EPS11 based on proteomic analysis. Finally, we verified the inhibition activity of EPS11 against metastasis in a melanoma mouse model.
3. Discussion
Numerous studies have shown that marine bacterial polysaccharides with novel chemical compositions, properties and structures have been found to have potential applications in pharmaceuticals and medicine as anti-cancer drugs, food additives and so on [
29]. EPS11, a novel marine bacterial polysaccharide, preferentially killed cancer cells including human lung cancer cells, A549, and HCV-related human liver cancer cells, Huh7.5, compared with normal cell line human embryonic lung fibroblasts, WI-38 [
9]. EPS11 was further demonstrated to inhibit lung cancer cell growth via blocking filopodia mediated adhesion and stimulating βIII-tubulin associated anoikis [
9]. However, the detailed molecular mechanisms against liver cancer and its potential anti-metastasis effect are not clear.
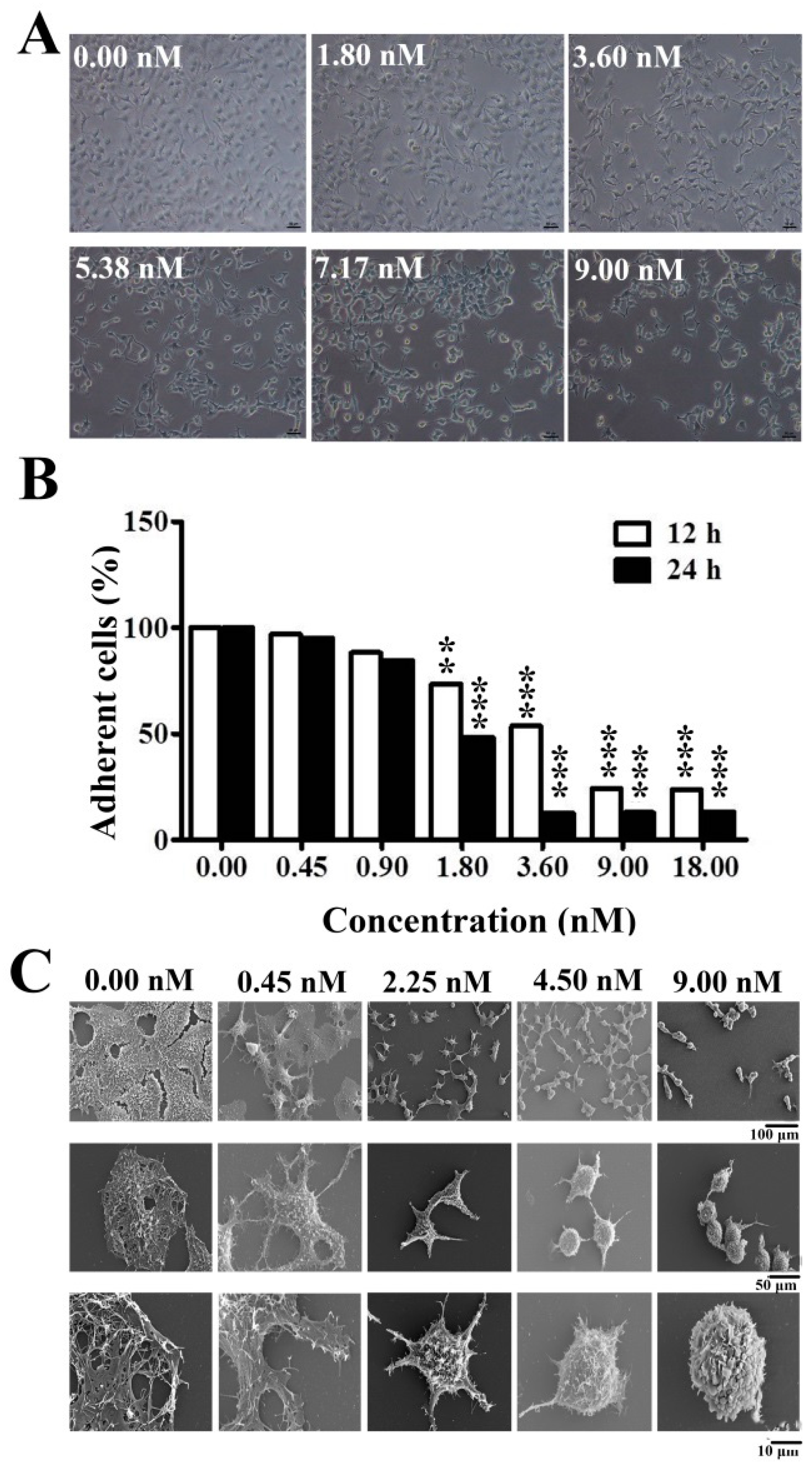
In this study, we first demonstrated that EPS11 inhibited cell growth and adhesion by destroying filiform structures in the liver cancer cell line, Huh7.5 (
Figure 2A,C), which is very similar to the effects on lung cancer cells treated with EPS11 [
9]. Filiform structures are thin, finger-like and highly dynamic actin-rich membrane protrusions that extend out from the cell edge and play a central role in modulating cell adhesion, migration and cancer cell invasion [
10]. Increased numbers of filiform structures have been implicated in the formation and reinforcement of cell-cell junctions, cell migration and invasion [
6]. In our study, the results showed that EPS11 destroyed Huh7.5 cell filiform structures in a dose-dependent manner (
Figure 2C). Meanwhile, destroying filiform structures has been associated with a reduced adhesion rate (
Figure 2B) and migration ability (
Figure 3A, B). Thus, EPS11 could inhibit Huh7.5 cell adhesion and migration ability by destroying filiform structures. Consistently, EPS11 also significantly suppressed the formation of filiform structures in three other lung cancer cell lines including A549, H1299 and H460 [
9], which strongly suggests that filiform structure is one of the key action targets of EPS11.
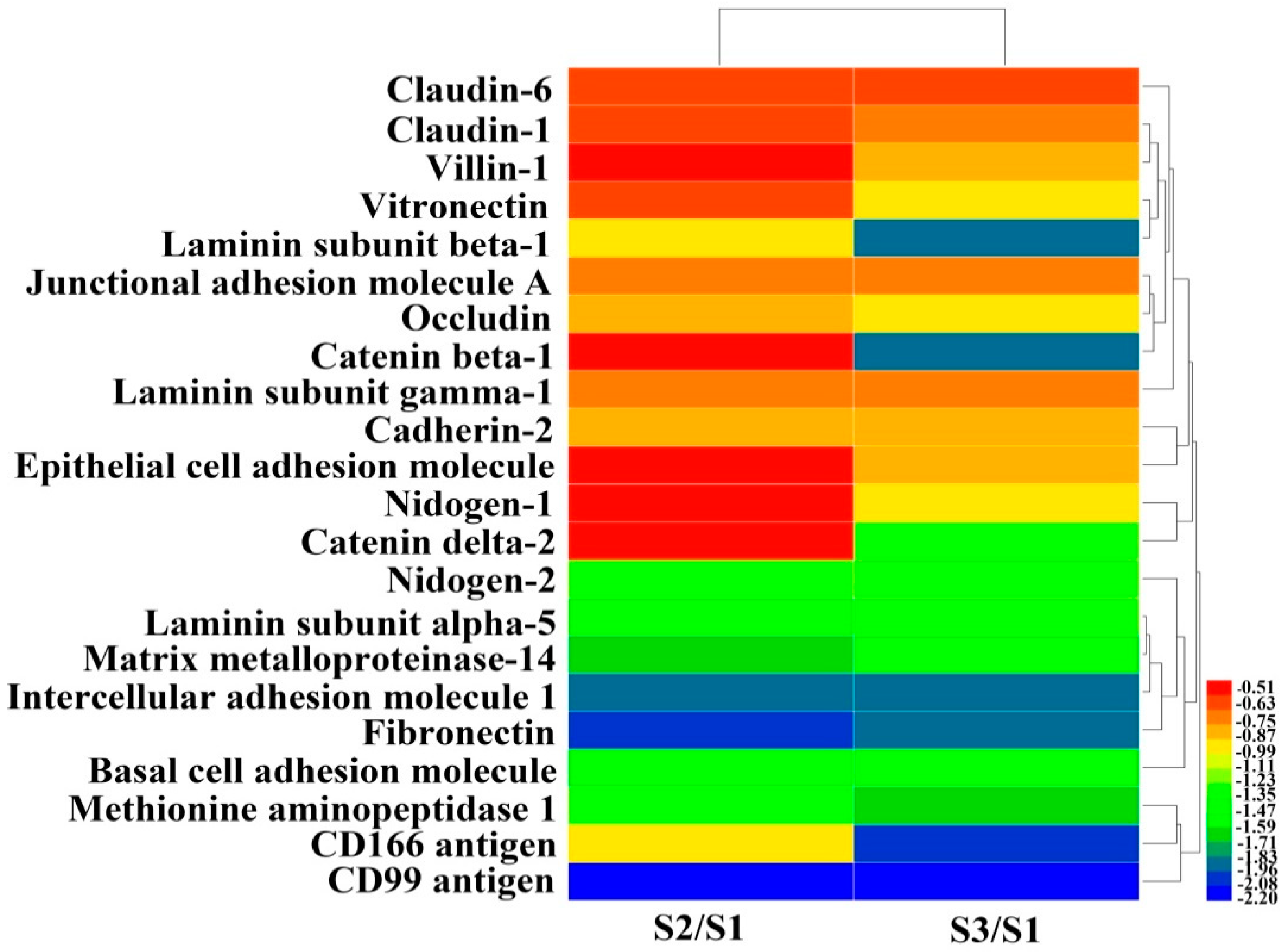
Presently, proteomic technologies not only provide a powerful tool to profile protein expression in response to various stresses in cancer cells, but are also used to reveal the possible mechanism of action of corresponding drugs [
30]. Based on the proteomic results of Huh7.5 treated with EPS11, we could see clearly that many proteins associated with cell adhesion, migration and metastasis were evidently down-regulated, which coincided with the results related to inhibition of cell adhesion and migration in Huh7.5 shown above (
Figure 2 and
Figure 3). Among these proteins, CD99 is reported to be involved in regulating tumor growth, differentiation, cell adhesion, cell migration and metastasis [
14]. High CD99 expression plays an oncogenetic role in Ewing sarcoma, lymphoblastic lymphoma, myeloid chondrosarcoma, malignant glioma and so on [
31,
32,
33,
34]. It is noteworthy that CD99 has a positive expression in hepatocellular carcinoma, but in non-HCC cancer, it has a negative expression [
15,
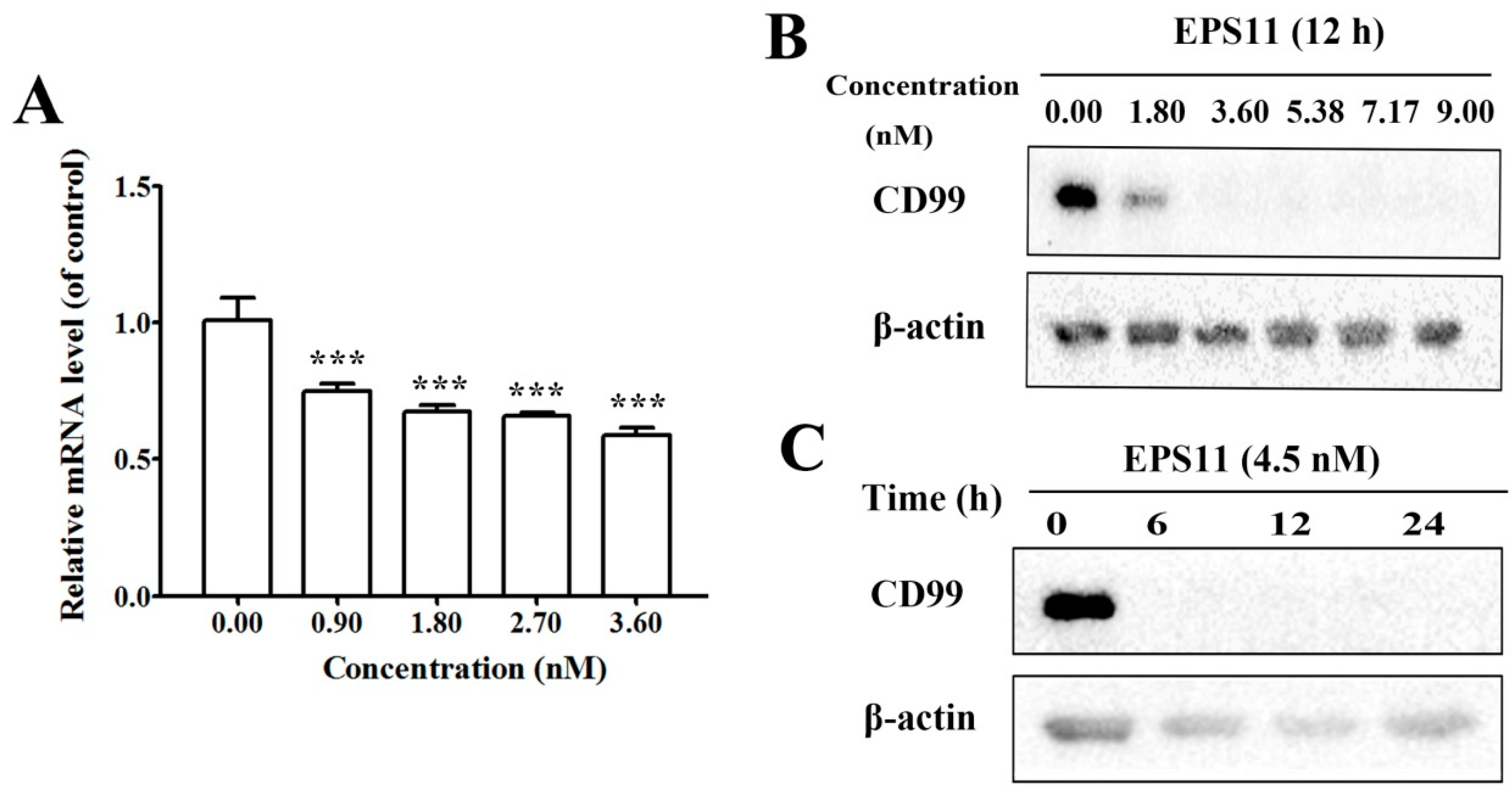
35]. Despite increasing evidence that CD99 has important functions in several aspects of cell biology, this molecule has been largely ignored by the scientific community. A number of unresolved issues remain to be clarified, particularly in terms of the mechanisms of action of CD99. In this study, EPS11 treatment significantly reduced CD99 expression in both mRNA and protein levels (
Figure 5A,B), which is consistent with the proteomic result (
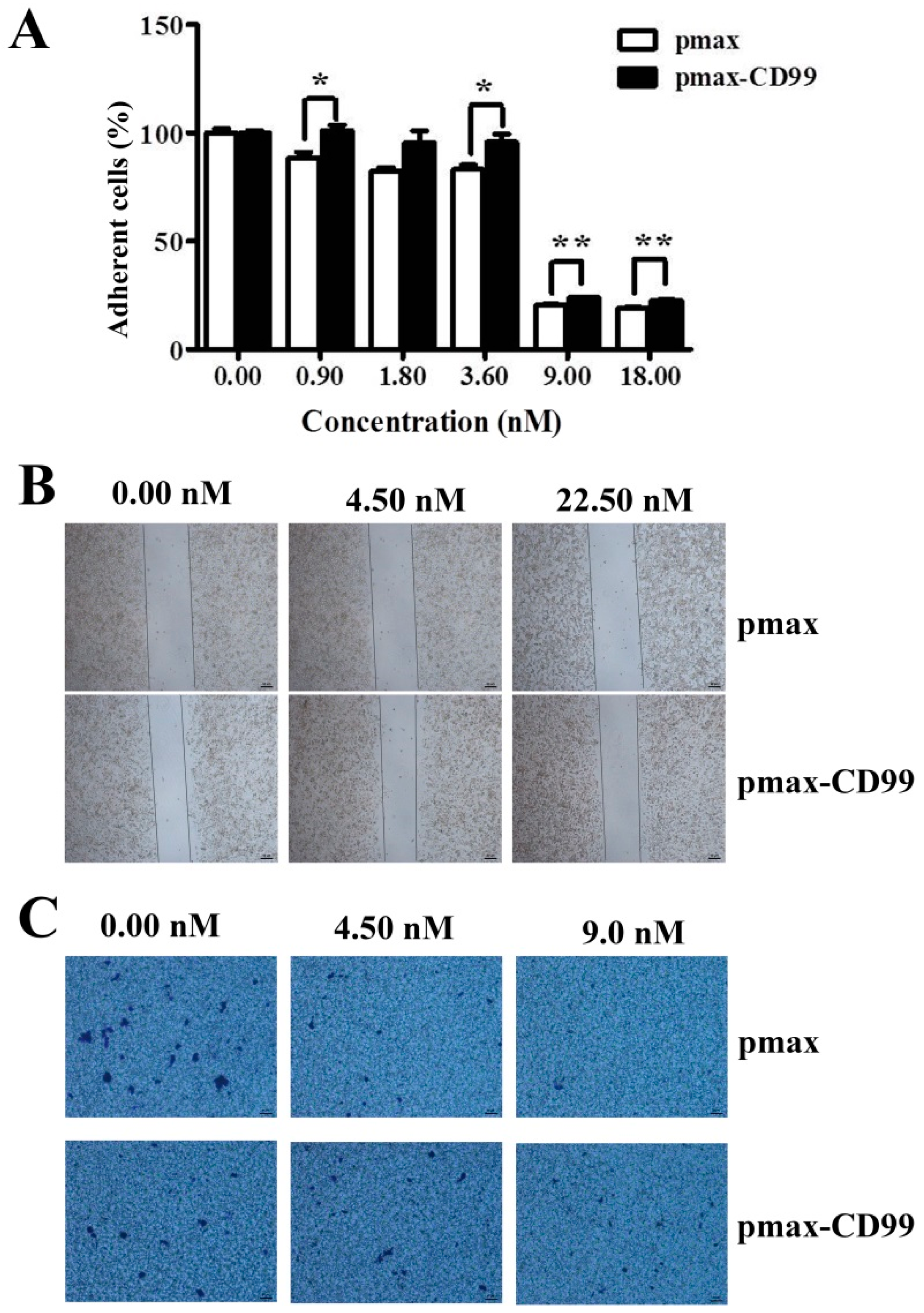
Figure 4). Furthermore, the over-expression of CD99 in Huh7.5 cells partly prevented the EPS11-mediated cell death, loss of adhesion rate and reduction of cell migration ability (
Figure 6B and
Figure 7A–C). The results suggested that the down-regulation of CD99 expression by EPS11 was partly responsible for the suppression of cell growth, cell adhesion and migration, and filiform structure formation mentioned above. In our previous results, EPS11 inhibited the growth of human non-small cell lung carcinoma via blocking filiform structure-mediated adhesion and stimulating βIII-tubulin-associated anoikis [
9]. However, the expression of βIII-tubulin in Huh7.5 was not evidently affected by EPS11, which indicates that EPS11 might inhibit cancer cell growth with slightly different mechanisms, even though it apparently attenuated different cell lines’ adhesion and led cells to aggregate.
Moreover, EPS11 significantly inhibited cell adhesion and migration, which are necessary for tumor metastasis. Metastasis is a complex, multistep process, during which tumor cells spread from the primary tumor mass to distant organs [
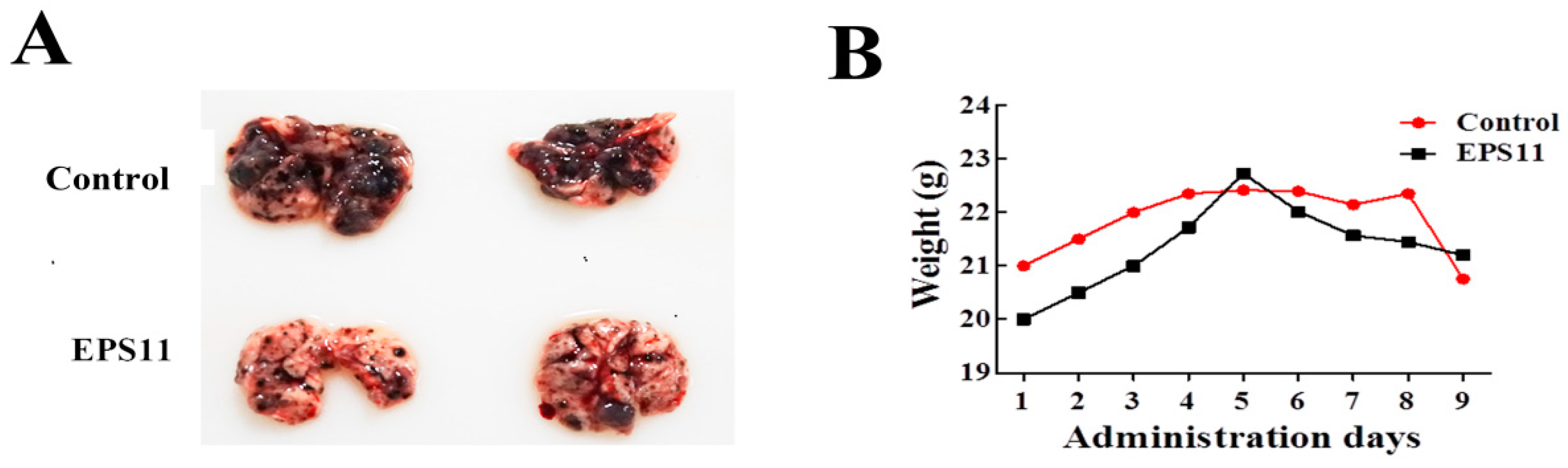
36]. The ability of cancer cells to disseminate from the primary site and form distant metastases is the main cause for cancer-related morbidity in patients with solid tumors. Notably, the melanoma tumor nodules in lung were significantly reduced after EPS11 treatment (
Figure 8A). Combining our previous and current results in different cancer cell lines, we conclude that EPS11 has potential to inhibit several key steps of metastasis, including cell adhesion, migration and filiform structure formation. Thus, it is reasonable to develop anti-cancer drugs targeting metastasis with EPS11 in the future. Moreover, our results found that several marine bacterial polysaccharides derived from costal and deep sea also dramatically attenuated the cell adhesion and migration and the formation of filiform structures in Huh7.5 (results not shown). Therefore, we believe that marine bacterial polysaccharides are a valuable resource to develop drugs blocking tumor metastasis in the future. However, there is still work needed to be done, such as the detailed mechanisms suppressing filiform structure formation, the exact action target and the elucidation of the structure of EPS11.
In summary, we have shown that EPS11 inhibited Huh7.5 cell growth and adhesion in vitro by destroying filiform structure and down-regulating the expression of CD99. Meanwhile, EPS11 had a significant anti-metastasis effect on melanoma in animal experiments. Thus, combining the anti-cancer effects of EPS11 towards human non-small cell lung carcinoma, we further confirmed that EPS11 should be a promising lead compound for novel anti-cancer drug development.
4. Materials and Methods
4.1. Cell Culture, Reagents and Antibodies
Huh7.5, B16F-10 cell lines were obtained from the American Type Culture Collection (Manassas, Virginia, USA), and they were cultured in RPMI 1640 medium supplemented with 10% FBS (all of them purchased from Gibico, Grand Island, NY, USA), penicillin at 100 units/mL, and streptomycin at 100 μg/mL (HyClone, Logan, UT, USA) in a humidified atmosphere of 5% CO2 at 37 °C. MTT, BCA kit for protein quantification were purchased from Beyotime Institute of Biotechnology (Shanghai, China), and the enhanced chemiluminescence (ECL) was purchased from Pierce (Thermo Scientific, Portsmouth, NH, USA). Anti-CD99 monoclonal was purchased from KleanAB (Sangon Biotech, Shanghai, China). Antibody against β-actin was obtained from Proteintech (Wuhan, China).
4.2. Purification and Anticancer Activity Assay of EPS11
The purification of EPS11 was carried out as previously described with minor modifications [
9]. Briefly, marine bacterium
Bacillus sp. 11 was cultured in 2216E medium (5 g/L tryptone, 1 g/L yeast extract, 1 L filtered seawater, pH adjusted to 7.4–7.6) supplemented with 1% sucrose at 150 rpm for 2 days at 28 °C. The culture supernatant was concentrated by centrifugation at a speed of 8000 rpm at 20 °C for 20 minutes, and the polysaccharide was precipitated with three volumes of 95% (v/v) ethanol. After keeping at 4 °C overnight, the precipitate was collected by centrifugation with a speed of 8000 rpm at 4 °C for 20 minutes and dissolved in distilled water. After proteins in the samples were removed by sevage reagent (chloroform/n-butyl alcohol = 5:1; v/v), the solution was dialyzed against ddH
2O with an 8000–14,000 molecular weight cutoff membrane. The crude polysaccharide was dissolved in ddH
2O and loaded onto a 5 mL HiTrapTM Q HP column (GE Healthcare, Little Chalfont, UK) equilibrated with start buffer (20 mM Tris–HCl, pH 8.0), then collected with eluent (2 M NaCl in 20 mM Tris–HCl, pH 8.0). The retained fraction was dialyzed and concentrated by lyophilized. To get high purity polysaccharide, the crude polysaccharide was further purified using HiloadTM 16/600 SephadexTM 200 column (GE Healthcare, Little Chalfont, UK) pre-equilibrated with 50 mM NaCl in 20 mM Tris-HCl (pH 9.0). The fractions were collected for 4 mL/tube, and the content of polysaccharide was analyzed by phenol-sulfuric acid assay. In phenol-sulfuric acid assay, 0.5 mL solutions from each fraction in the gel filtration were mixed with 0.5 mL 5% phenol respectively, and then 2.5 mL 95.5% sulfuric acid was added. After mixing and cooling down for 20 minutes at 20 °C, the polysaccharide content was determined at OD490 nm. The corresponding ability inhibiting Huh7.5 cells growth was detected by MTT assay described as following.
4.3. Cell Proliferation Viability Assay
Viabilities of Huh7.5 cells were measured by the MTT method. Briefly, exponentially growing Huh7.5 cells (5 × 104 cell/mL) were routinely seeded into 96-well plate at 37 °C for overnight. Then the cells were treated with different concentrations of EPS11 (0–180 nM) for 24 hours and 48 hours. 30 μL of 5 mg/mL MTT (Sigma, St. Louis, MO, USA) was added into each well and incubated for 3 hours at 37 °C, and 100 μL “Triplex Solution” (10% SDS-5% isobutanol-12 mM HCl) was added to each well. After incubation overnight, the absorbance of each well was measured at a wavelength of 570 nm by a multi-detection microplate reader (Infinite M1000 Pro, TECAN, Mannedorf, Switzerland). Relative cell viability was presented as a percentage relative to the control group. All experiments were performed three times.
4.4. Scanning Electron Microscope (SEM)
Huh7.5 cells were plated onto laminin-coated glass coverslips overnight and treated with different concentrations of EPS11 (0–9.0 nM). After incubation for 6 hours, the glass coverslips were washed with PBS and fixed with 5.0% glutaradehyde in PBS and gradually dehydrated in ethanol (30%, 50%, 70%, 90% and 100% for 10 minutes at each step). Finally, the glass coverslips were observed and imaged by SEM (Hitachi S-3400N, Tokyo, Japan).
4.5. Cell Adhesion Assay
Huh7.5 cells (5 × 104/mL) were seeded into a 96-well plate at 37 °C overnight and then treated with varying concentrations of EPS11 (0–18 nM) for 12 hours and 24 hours. Cell culture medium and the non-adherent cells were discarded and washed three times with PBS, fixed with 95% ethanol for 30 minutes, and then stained with 100 μL 0.1% crystal violet for 20 minutes. While detached cells were washed off, only adhesive cells could be stained. After removing the redundant crystal violet, 100 μL acetic acid was added into each well with gentle shaking for 10 minutes. The absorbance was measured by a multi-detection microplate reader (Infinite M1000 Pro, TECAN, Mannedorf, Switzerland). Relative adhered cells were presented as a percentage relative to the control group. All experiments were performed three times.
4.6. Wound Healing Migration Assay
Huh7.5 cells migration ability treated with EPS11 was measured by wound healing assay. Briefly, Huh7.5 cells were incubated in serum-free medium overnight for synchronization and then trypsinized from culture dishes and placed into 12-well plate with a Culture-Insert 2 well. After growth to 90% confluence, the Culture-Insert 2 well was gently removed by sterile tweezers. After washing three times with PBS, the cells were exposed to a medium supplemented with 2% FBS with or without EPS11 (0, 0.9 and 1.8 nM). After incubation for 24 hours, three fields of each wound were selected and photographed with an inverted microscope (NIKON TS100, Tokyo, Japan) equipped with a digital camera.
4.7. Transwell Migration Assay
The migration ability of Huh7.5 cells was measured by a Transwell Boyden chamber (Costar; Corning Life Sciences, Lowell, MA, USA). Briefly, Huh7.5 cells were cultured in serum–free medium RPMI 1640 for overnight, then the cells were suspended in a 200 μL medium containing different concentrations of EPS11 (0–9.0 nM) that was added into the upper compartment, then 600 μL medium supplemented with 20% FBS was added into the lower compartment. After incubation at 37 °C for 8 hours, cell culture medium was discarded and the upper surface cells were gently removed by cotton swabs. After washing with PBS, cells were fixed with 95% ethanol and stained with 0.1% crystal violet. Then the non-migrated cells on the upper side of the filter were gently removed using cotton swabs and the migrated cells on the lower side of the filter were observed and counted in five random fields.
4.8. Proteomic Analysis
Proteomic analysis of Huh7.5 cells treated with EPS11 was performed by PTM Biolabs Co., Ltd. (HangZhou, Zhejiang, China). Briefly, Huh7.5 cells were treated with different concentrations of EPS11 (0, 45, 90 nM) for 24 hours, and cells were collected and lysed to obtain total cellular protein. Protein samples were then digested, labeled, separated and quantified by LC-ESI-MS/MS analysis. The bioinformatic analyses of protein annotation, functional classification, functional enrichment and cluster analyses were then performed as described previously [
37].
4.9. Quantitative Reverse Transcription-PCR (qRT-PCR)
Huh7.5 cells were treated with different concentrations of EPS11 (0–3.6 nM) for 12 hours. Total cellular RNA was extracted using the TRIpure reagent (Aidlad, Beijing, China) according to the manufacturer’s instructions. Quality and quantity of RNA samples were determined by NanoDrop analysis (NanoDrop 2000, Thermo, Beijing, China). RNA was reverse transcribed into cDNA by using a Prime Script® RT reagent kit (Takara Biotechnology Co., Ltd., Dalian, China). Then qRT-PCR was carried out with SYBR premix real-time PCR Reagents (Takara Biotechnology Co, Ltd., Dalian, China) by an ABI7900 real-time PCR system (Applied Biosystems, Foster City, CA, USA). Housekeeping gene β-actin was used as an internal control. The comparative 2−ΔΔCt method was used to calculate the relative expression. For CD99-F1: 5′-GCCACAGGAAAGAAGGGGAA-3′, CD99-R1: 5′-CCCTTGTTC TGCATTTTCTTTGA-3′, β-actin-F: 5′-CACGATGGAGGGCCGGACTCATC-3′, β-actin-R: 5′-TAAAGACCTCTATGCCAACACAGT-3′. All qRT-PCR runs were conducted with three biological and three technical replicates.
4.10. Western Blot Analysis
Huh7.5 cells were treated with varying concentrations of EPS11 (0–9 nM) for different times. Cells were then collected and lysed with RIPA buffer (Sigma, St. Louis, MO, USA) with protease inhibitor PMSF (Sigma, St. Louis, MO, USA). Equal amounts of protein determined by BCA kit (Beyotime, Shanghai, China) were separated by 12% SDS-PAGE gels, and then electro-transferred to nitrocellulose membranes and incubated with primary antibodies (anti-CD99, anti-β-actin) and secondary antibodies. Finally, the results were analyzed with an ECL chemiluminescence kit and a Molecular Imager® ChemiDoc™XRS system (Bio-Rad Laboratories, Inc., Pleasenton, CA, USA). Anti-β-actin was used to normalize protein loading.
4.11. DNA Constructs and Transfection
Total RNA of Huh7.5 cells was extracted with the TRIpure reagent (Aidlad, China) according to the manufacturer’s instructions, and then reverse transcribed into cDNA by using a Prime Script® RT reagent kit (Takara Biotechnology Co, Ltd, Dalian, China). The full-length cDNA sequence of CD99 was produced by polymerase chain reaction (PCR) with the primers CD99-F2 (5′-CGGGGTACCATGGCCCGCGGGGCTGC-3′) and CD99-R2 (5′- CCGCTCGAGCTATTTCTCTAAAAGAGTACG-3′). The PCR products were purified, digested with KpnI and XhoI, and ligated into the pmax vector (Lonza Colgne, Koln, Germany). The resulting plasmid (pmax-CD99) was verified by DNA sequencing.
The plasmids pmax and pmax-CD99 were transfected into Huh7.5 cells using the translipofectamine (Bioino, Qingdao, China) according to the manufacture’s protocol. After transfection, cells were collected at 48 hours post-transfection, and proteins were extracted. Protein samples were resolved on 12% SDS-PAGE gels, electro-transferred to nitrocellulose membranes and incubated with primary antibodies against CD99, β-actin and secondary antibodies, and finally detected by enhanced chemiluminescence.
4.12. Pulmonary Colonization Assay
C57BL/6 mice (7-week-old males) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd (Beijing, China). All studies on mice were approved by IOCAS (Institute of Oceanology, Chinese Academy of Sciences) Laboratory Animal Care and Ethics Committee in accordance with the animal care and use guidelines. All animal procedures were conducted in accordance with all appropriate regulatory standards under protocol Haifajizi 2013-3 (approval date: 2013-12-09). Metastases were established by giving lateral tail vein injection of 4 × 105 B16F-10 melanoma cells into mice and divided into 2 groups. Mice were simultaneously administrated with EPS11 dose at 200 mg/kg (0.9 μmol/L/kg on average) once every two days by intraperitoneal injection, while equivoluminal normal saline was injected for the control group. Drug-treatment lasted for 18 days. Body weight was measured every two days with a balance for the average value. After 18 days treatment, the mice were euthanized to excise the lungs. The lungs were used for morphological examination of melanoma metastatic tumor nodules.
4.13. Statistical Analysis
All data are expressed as means ± SD. Statistical analysis was performed using GraphPad’s Prism v5.0 (San Diego, USA). Differences of p < 0.05 were considered statistically significant (*p < 0.05, **p < 0.01).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







