Discovery of Antibiofilm Activity of Elasnin against Marine Biofilms and Its Application in the Marine Antifouling Coatings

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
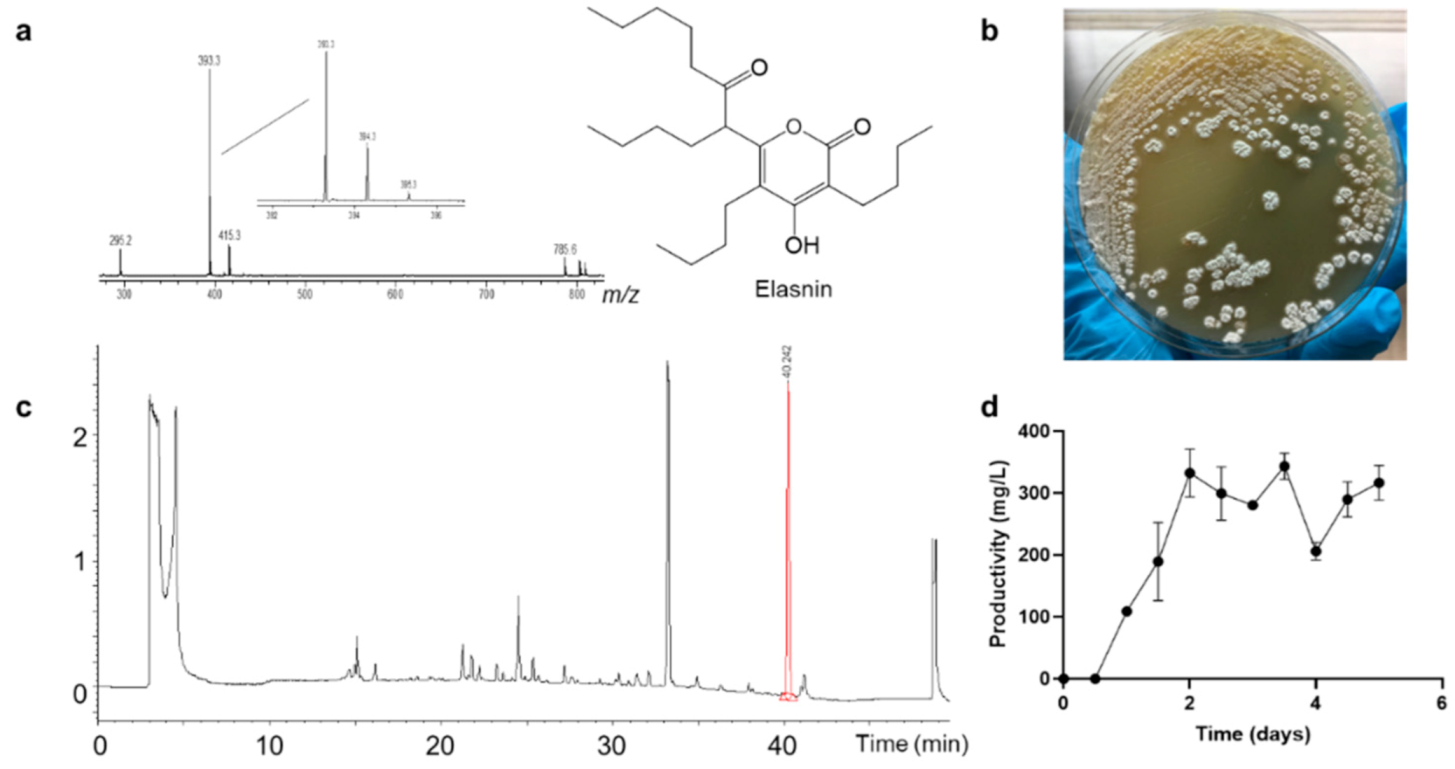
2.1. Isolation and Identification of Biofilm Inhibition Compounds
2.2. Elasnin Could Inhibit the Biofilm Formation of Multiple Strains of Bacteria Isolated from Marine Biofilms
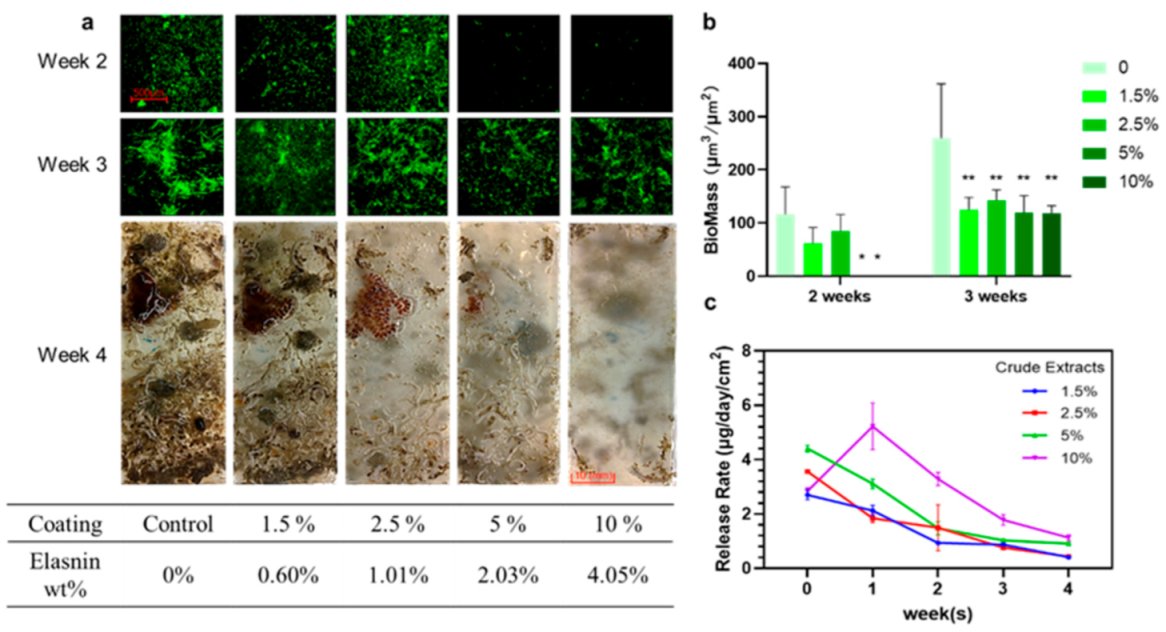
2.3. Preparation of Elasnin-Based Antibiofilm Coatings
2.4. Release Rate of Elasnin from Antibiofilm Coatings
2.5. Elasnin-Based Coatings Inhibited the Formation of Multi-Species Biofilms and the Attachment of Large Biofouling Organisms in the Marine Environment
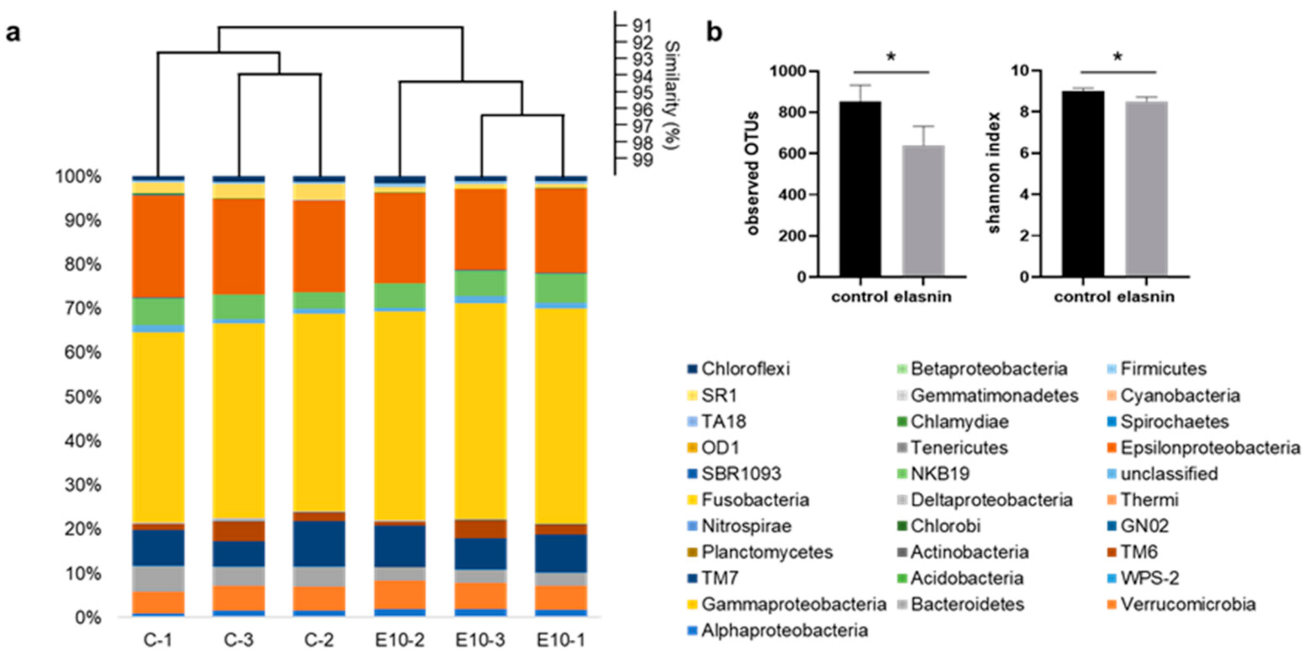
2.6. Elasnin Changed the Microbial Community Structure of Natural Marine Biofilms
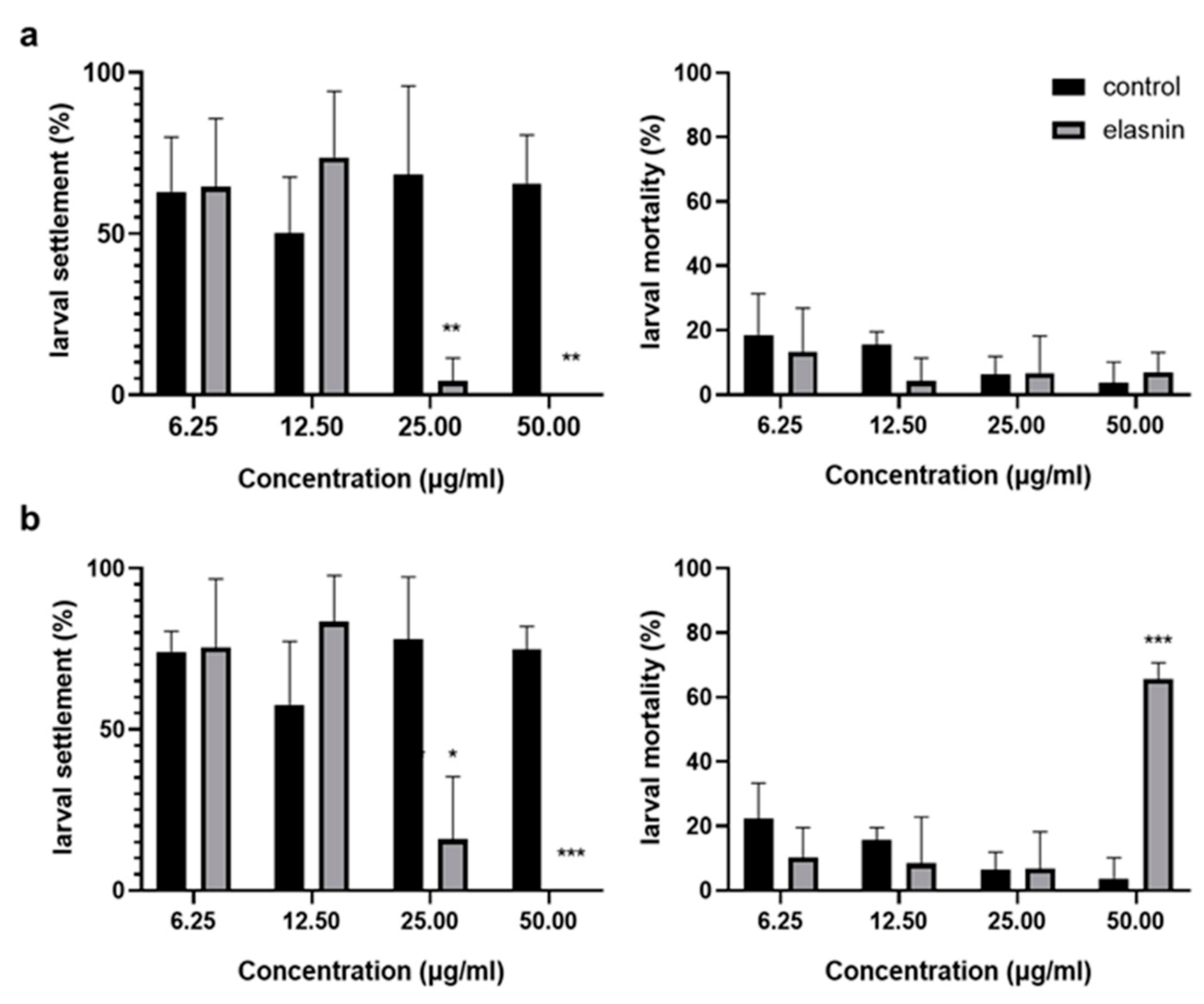
2.7. Elasnin Inhibited the Larval Settlement of Balanus Amphitrite with a Low Toxic Effect
3. Discussion
4. Materials and Methods
4.1. Strains, Culture Media, and Chemicals
4.2. Bioactive Compound Isolation and Identification
4.3. Productivity Monitoring and Extraction Efficiency Comparison
4.4. MBIC Assay and MIC Assay Against Marine Bacteria
4.5. Elasnin-Based Antibiofilm Coating Preparation
4.6. Field Test and Release Rate Determination
4.7. DNA Extraction, 16S rRNA Gene Sequencing, and Analyses
4.8. Antilarval-Settlement Assay
4.9. Statistical Analyses
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; O’Toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, L.C.M.; Ferreira, R.B.R. Biofilms and bacterial virulence. Rev. Med. Microbiol. 2011, 22, 12–16. [Google Scholar] [CrossRef]
- Lopez, D.; Vlamakis, H.; Kolter, R. Biofilms. Cold Spring Harb. Perspect. Biol. 2010, 2, a000398. [Google Scholar] [CrossRef]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Hengzhuang, W.; Wu, H.; Ciofu, O.; Song, Z.; Hoiby, N. Pharmacokinetics/pharmacodynamics of colistin and imipenem on mucoid and nonmucoid Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2011, 55, 4469–4474. [Google Scholar] [CrossRef] [Green Version]
- Qian, P.Y.; Lau, S.C.; Dahms, H.U.; Dobretsov, S.; Harder, T. Marine biofilms as mediators of colonization by marine macroorganisms: Implications for antifouling and aquaculture. Mar. Biotechnol. 2007, 9, 399–410. [Google Scholar] [CrossRef]
- Lau, S.C.K.; Thiyagarajan, V.; Cheung, S.C.K.; Qian, P.Y. Roles of bacterial community composition in biofilms as a mediator for larval settlement of three marine invertebrates. Aquat. Microbial. Ecol. 2005, 38, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Hung, O.S.; Thiyagarajan, V.; Wu, R.S.S.; Qian, P.Y. Effect of ultraviolet radiation on biofilms and subsequent larval settlement of Hydroides elegans. Mar. Ecol. Prog. Ser. 2005, 304, 155–166. [Google Scholar] [CrossRef]
- Dobretsov, S.; Qian, P.-Y. Facilitation and inhibition of larval attachment of the bryozoan Bugula neritina in association with mono-species and multi-species biofilms. J. Exp. Mar. Biol. Ecol. 2006, 333, 263–274. [Google Scholar] [CrossRef]
- De Carvalho, C.C.C.R. Marine biofilms: A successful microbial strategy with economic implications. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Lehaitre, M.; Delauney, L.; Compère, C. Biofouling and underwater measurements. In Real-Time Observation Systems for Ecosystem Dynamics and Harmful Algal Blooms: Theory, Instrumentation and Modelling; Unesco Publishing: Paris, France, 2008; pp. 463–493. [Google Scholar]
- Plaza, G.; Achal, V. Biosurfactants: Eco-friendly and innovative biocides against biocorrosion. Int. J. Mol. Sci. 2020, 21, 2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Yu, L.; Mou, J.; Wu, D.; Xu, M.; Zhou, P.; Ren, Y. Research strategies to develop environmentally friendly marine antifouling coatings. Mar. Drugs 2020, 18, 371. [Google Scholar] [CrossRef]
- Dang, H.; Li, T.; Chen, M.; Huang, G. Cross-ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl. Environ. Microbiol. 2008, 74, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Moffat, J.G.; Vincent, F.; Lee, J.A.; Eder, J.; Prunotto, M. Opportunities and challenges in phenotypic drug discovery: An industry perspective. Nat. Rev. Drug Discov. 2017, 16, 531–543. [Google Scholar] [CrossRef]
- Scannell, J.W.; Bosley, J. When quality beats quantity: Decision theory, drug discovery, and the reproducibility crisis. PLoS ONE 2016, 11, e0147215. [Google Scholar] [CrossRef] [Green Version]
- Pollet, T.; Berdjeb, L.; Garnier, C.; Durrieu, G.; Le Poupon, C.; Misson, B.; Jean-Francois, B. Prokaryotic community successions and interactions in marine biofilms: The key role of Flavobacteriia. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [Green Version]
- Salta, M.; Wharton, J.A.; Blache, Y.; Stokes, K.R.; Briand, J.F. Marine biofilms on artificial surfaces: Structure and dynamics. Environ. Microbiol. 2013, 15, 2879–2893. [Google Scholar] [CrossRef] [PubMed]
- Ohno, H.; Saheki, T.; Awaya, J.; Nakagawa, A.; Omura, S. Isolation and characterization of elasnin, a new human granulocyte elastase inhibitor produced by a strain of Streptomyces. J. Antibiot. 1978, 31, 1116–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Ding, W.; Long, L.; Lan, Y.; Saha, S.; Wong, Y.H.; Sun, J.; Li, Y.; Zhang, W.; Qian, P.-Y. Exploring the influence of signal molecules on marine biofilm development. Front. Microbiol. 2020, 11, 2927. [Google Scholar] [CrossRef]
- Yin, Q.; Liang, J.; Zhang, W.; Zhang, L.; Hu, Z.L.; Zhang, Y.; Xu, Y. Butenolide, a marine-derived broad-spectrum antibiofilm agent against both gram-positive and gram-negative pathogenic bacteria. Mar. Biotechnol. 2019, 21, 88–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, S.; Desai, S.; Poonacha, N.; Vipra, A.; Sharma, U. Antibiofilm activity and synergistic inhibition of Staphylococcus aureus biofilms by bactericidal protein P128 in combination with antibiotics. Antimicrob. Agents Chemother. 2016, 60, 7280–7289. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Qian, P.Y.; Pechenik, J.A. Effects of larval starvation and delayed metamorphosis on juvenile survival and growth of the tube-dwelling polychaete Hydroides elegans (Haswell). J. Exp. Mar. Biol. Ecol. 1998, 227, 169–185. [Google Scholar] [CrossRef]
- Harder, T.N.; Thiyagarajan, V.; Qian, P.Y. Effect of cyprid age on the settlement of Balanus amphitrite darwin in response to natural biofilms. Biofouling 2001, 17, 211–219. [Google Scholar] [CrossRef]
- Thiyagarajan, V.; Harder, T.; Qiu, J.-W.; Qian, P.-Y. Energy content at metamorphosis and growth rate of the early juvenile barnacle Balanus amphitrite. Mar. Biol. 2003, 143, 543–554. [Google Scholar] [CrossRef]
- Gao, C.H.; Tian, X.P.; Qi, S.H.; Luo, X.M.; Wang, P.; Zhang, S. Antibacterial and antilarval compounds from marine gorgonian-associated bacterium Bacillus amyloliquefaciens SCSIO 00856. J. Antibiot. 2010, 63, 191–193. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, L.; Wang, R.; Chiang, H.Y.; Ding, W.; Li, Y.-X.; Chen, F.; Qian, P.-Y. Discovery of Antibiofilm Activity of Elasnin against Marine Biofilms and Its Application in the Marine Antifouling Coatings. Mar. Drugs 2021, 19, 19. https://0-doi-org.brum.beds.ac.uk/10.3390/md19010019
Long L, Wang R, Chiang HY, Ding W, Li Y-X, Chen F, Qian P-Y. Discovery of Antibiofilm Activity of Elasnin against Marine Biofilms and Its Application in the Marine Antifouling Coatings. Marine Drugs. 2021; 19(1):19. https://0-doi-org.brum.beds.ac.uk/10.3390/md19010019
Chicago/Turabian StyleLong, Lexin, Ruojun Wang, Ho Yin Chiang, Wei Ding, Yong-Xin Li, Feng Chen, and Pei-Yuan Qian. 2021. "Discovery of Antibiofilm Activity of Elasnin against Marine Biofilms and Its Application in the Marine Antifouling Coatings" Marine Drugs 19, no. 1: 19. https://0-doi-org.brum.beds.ac.uk/10.3390/md19010019