Bioactive Lipids of Marine Microalga Chlorococcum sp. SABC 012504 with Anti-Inflammatory and Anti-Thrombotic Activities

 ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
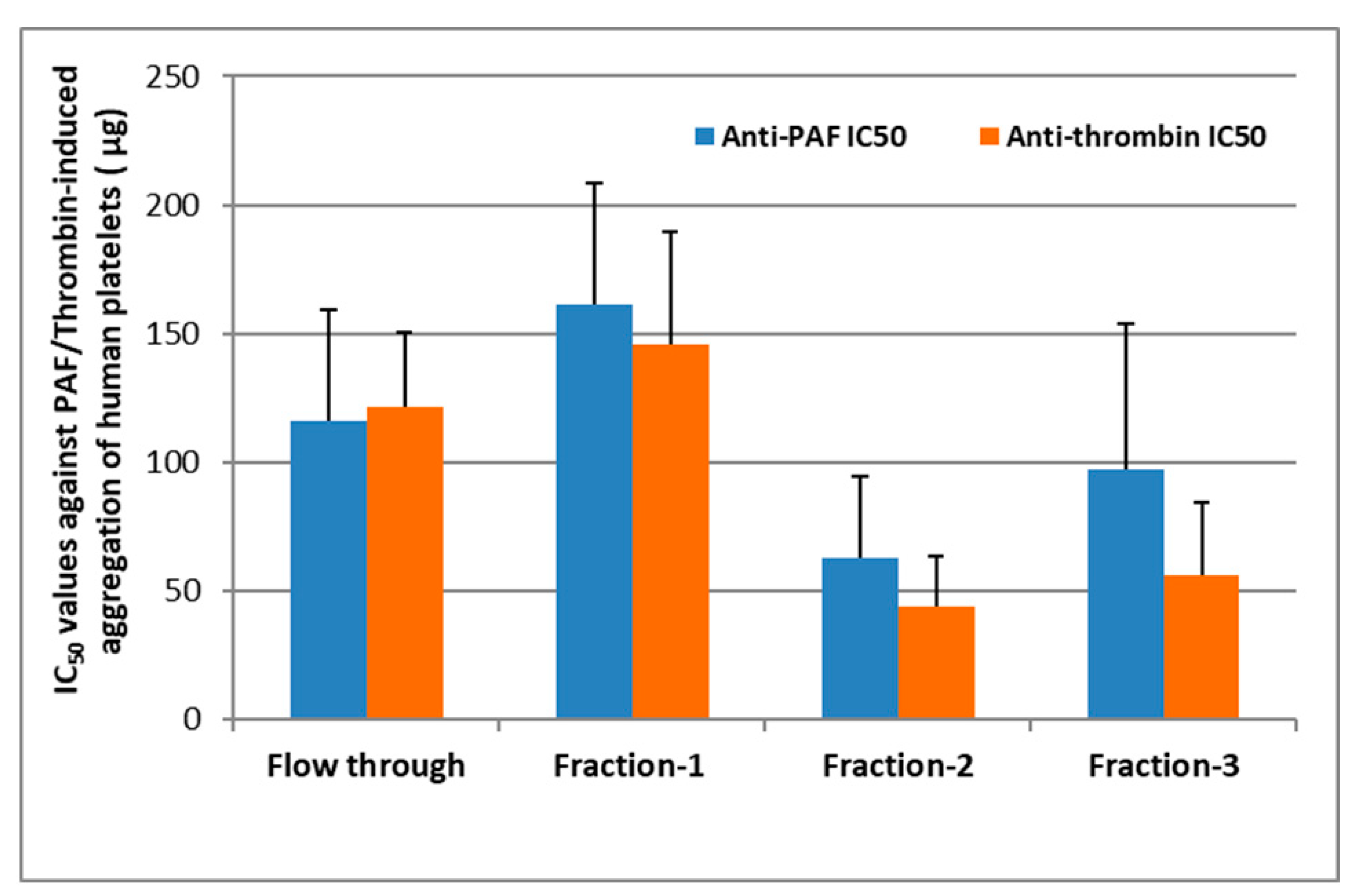
2.1. Bioactivity of Lipid Fractions
2.2. Fatty Acids Analysis
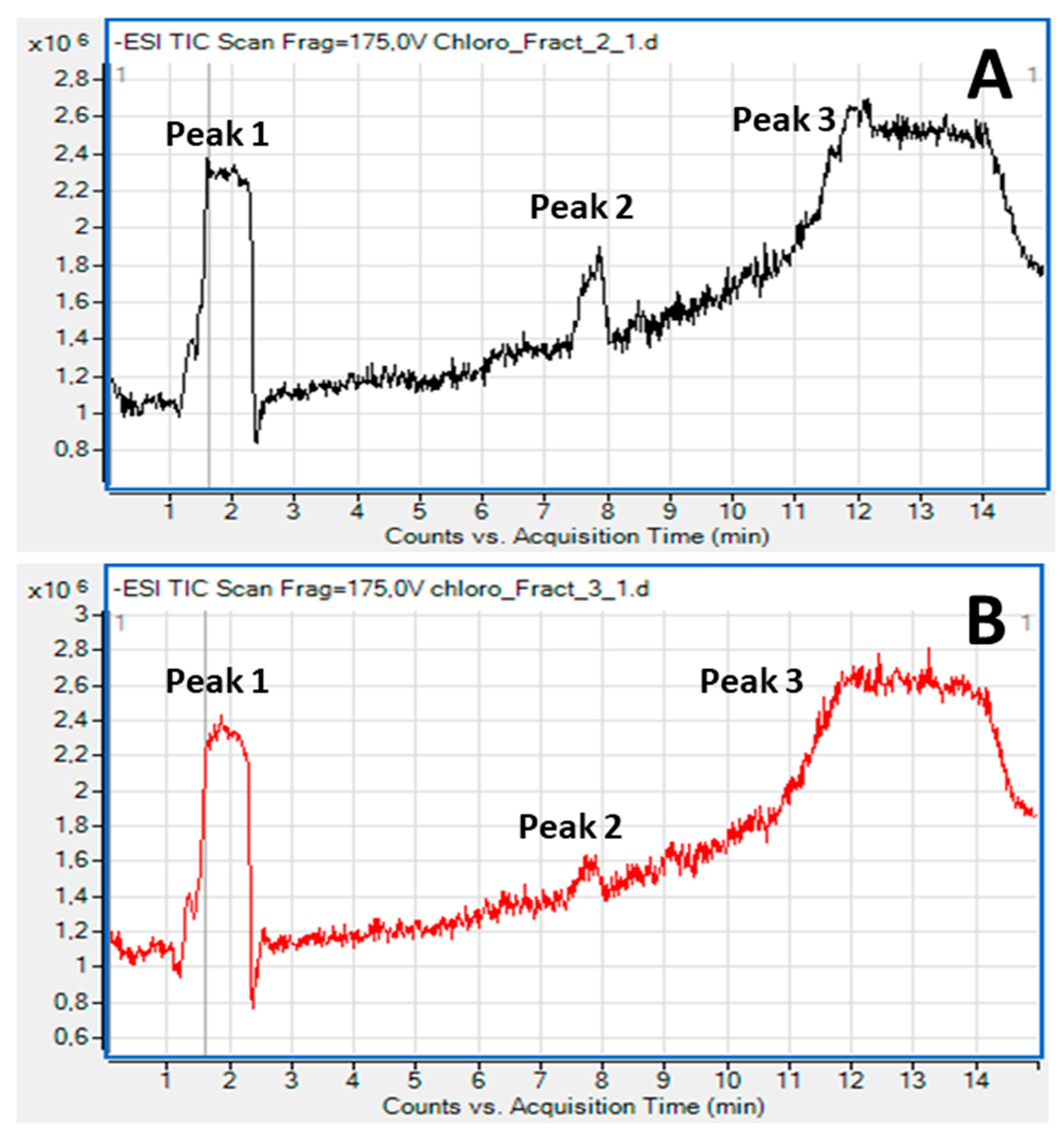
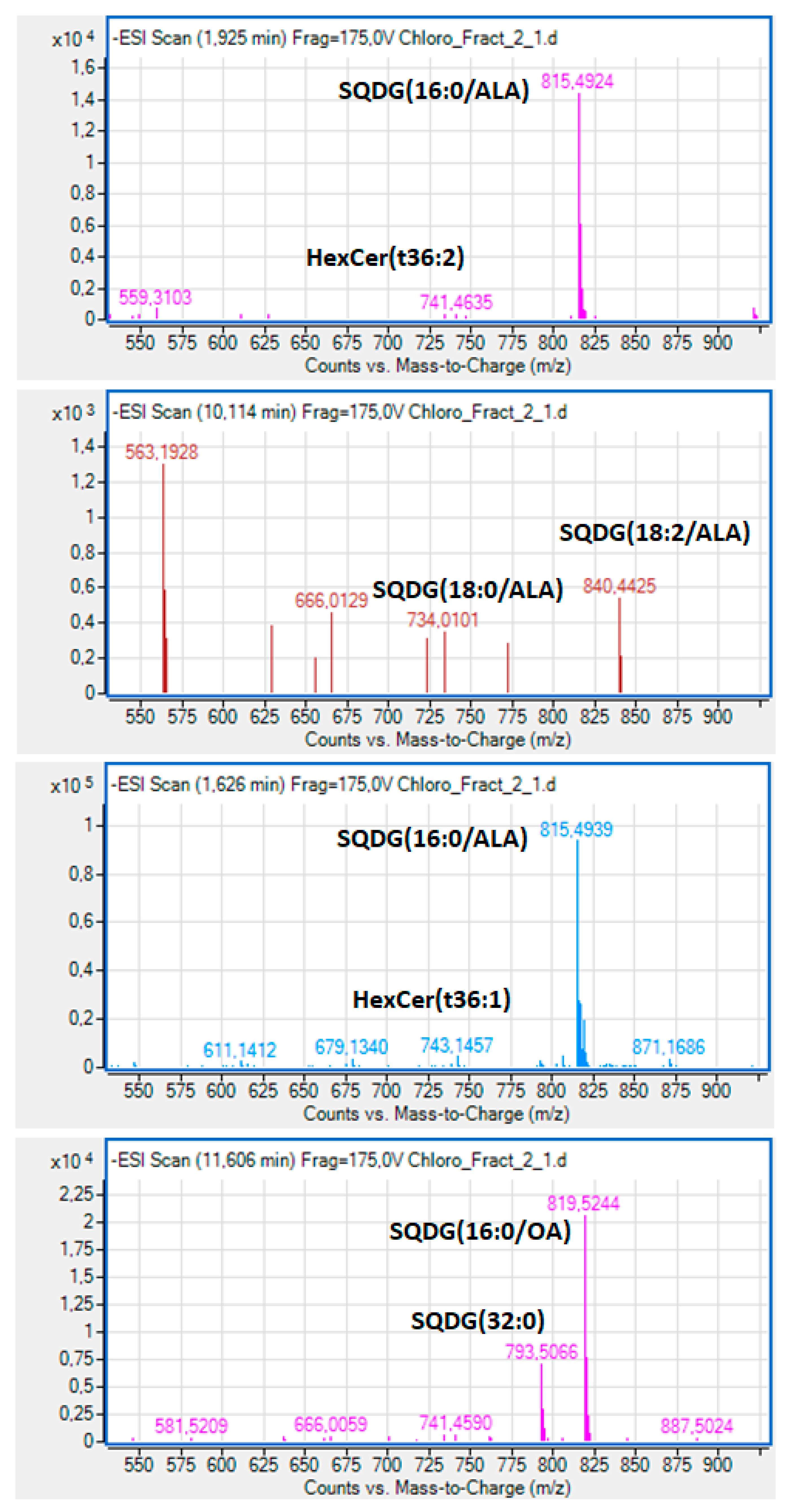
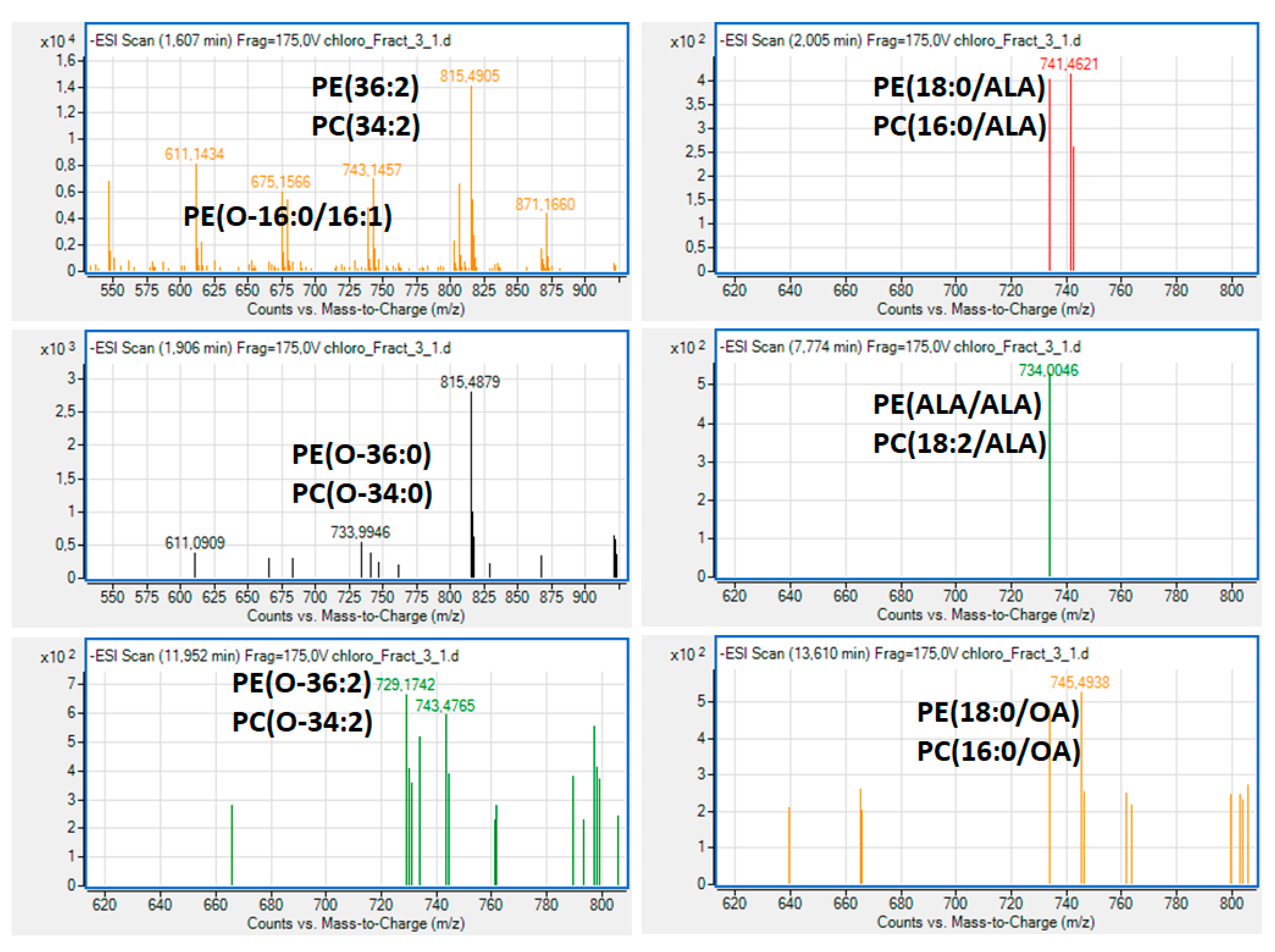
2.3. Bioactive Lipids
3. Materials and Methods
3.1. Microalgal Isolate
3.2. Culturing of Microalga
3.3. Bioactive Lipids Extraction and Fractionation
3.4. Fatty Acid Composition and Structural Elucidation of Microalgal Lipid Fractions by LC-MS Analysis
3.5. Human Platelet-Rich Plasma (hPRP) Aggregation Studies of Microalgal Lipid Fractions
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Becker, E.W. Microalgae: Biotechnology and Microbiology; Cambridge University Press: Cambridge, UK, 1994; ISBN 0-521-35020-4. [Google Scholar]
- Balandrin, M.F.; Klocke, J.A.; Wurtele, E.S.; Bollinger, W.H. Natural plant chemicals: Sources of industrial and medicinal materials. Science 1985, 228, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulou, S.; Karantonis, H.C.; Nomikos, T.; Oikonomou, A.; Fragopoulou, E.; Pantazidou, A. Bioactive polar lipids from Chroococcidiopsis sp. (Cyanobacteria). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 142, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Koukouraki, P.; Tsoupras, A.; Sotiroudis, G.; Demopoulos, C.A.; Sotiroudis, T.G. Antithrombotic properties of Spirulina extracts against platelet-activating factor and thrombin. Food Biosci. 2020, 37, 100686. [Google Scholar] [CrossRef]
- Saha, S.K. Microalgae and cyanobacterial feedstocks for multiple commodities. In Feedstocks: Production Practices, Technologies and Environmental Impacts; Acquaye, A., Ed.; Nova: New York, NY, USA, 2014; pp. 73–107. [Google Scholar]
- Saha, S.K.; McHugh, E.; Murray, P.; Walsh, D.J. Microalgae as a source of nutraceuticals. In Phycotoxins: Chemistry and Biochemistry, 2nd ed.; Botana, L.M., Alfonso, A., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2015; pp. 255–292. [Google Scholar]
- Saha, S.K.; Murray, P. Exploitation of microalgae species for nutraceutical purposes: Cultivation aspects. Fermentation 2018, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Tsoupras, A.B.; Iatrou, C.; Frangia, C.; Demopoulos, C.A. The implication of platelet-activating factor in cancer growth and metastasis: Potent beneficial role of paf-inhibitors and antioxidants. Infect. Disord. Drug Targets 2009, 9, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation, not Cholesterol, Is a Cause of Chronic Disease. Nutrients 2018, 10, 604. [Google Scholar] [CrossRef] [Green Version]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Thrombosis and COVID-19: The Potential Role of Nutrition. Front. Nutr. 2020, 7, 583080. [Google Scholar] [CrossRef]
- Zabetakis, I.; Lordan, R.; Norton, C.; Tsoupras, A. COVID-19: The Inflammation Link and the Role of Nutrition in Potential Mitigation. Nutrients 2020, 12, 1466. [Google Scholar] [CrossRef]
- Lordan, R.; Tsoupras, A.; Zabetakis, I. Platelet activation and prothrombotic mediators at the nexus of inflammation and atherosclerosis: Potential role of antiplatelet agents. Blood Rev. 2020, 100694, (online ahead of print). [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation and Cardiovascular Diseases. In The Impact of Nutrition and Statins on Cardiovascular Diseases; Zabetakis, I., Lordan, R., Tsoupras, A., Eds.; Academic Press: Cambridge, MA, USA, 2019; Volume 3, pp. 53–117. [Google Scholar]
- Lordan, R.; Tsoupras, A.; Zabetakis, I. Phospholipids of animal and marine origin: Structure, function, and anti-inflammatory properties. Molecules 2017, 22, 1964. [Google Scholar] [CrossRef] [Green Version]
- Nasopoulou, C.; Tsoupras, A.B.; Karantonis, H.C.; Demopoulos, C.A.; Zabetakis, I. Fish polar lipids retard atherosclerosis in rabbits by down-regulating paf biosynthesis and up-regulating paf catabolism. Lipids Health Dis. 2011, 10, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.P.; Chen, F.; Liu, X.; Lic, X.Z. Carotenoid composition in the green microalga Chlorococcum. Food Chem. 2002, 76, 319–325. [Google Scholar] [CrossRef]
- Zhang, D.H.; Lee, Y.K. Ketocarotenoid production by a mutant of Chlorococcum sp. in an outdoor tubular photobioreactor. Biotechnol. Lett. 1999, 21, 7–10. [Google Scholar] [CrossRef]
- Saha, S.K.; McHugh, E.; Hayes, J.; Moane, S.; Walsh, D.; Murray, P. Effect of various stress-regulatory factors on growth and lipids biosynthesis in microalga Haematococcus pluvialis. Bioresour. Technol. 2013, 128, 118–124. [Google Scholar] [CrossRef]
- Beevi, U.S.; Sukumaran, R.K. Cultivation of the fresh water microalga Chlorococcum sp. RAP13 in sea water for producing oil suitable for biodiesel. J. Appl. Phycol. 2015, 27, 141–147. [Google Scholar] [CrossRef]
- Antonopoulou, S.; Nomikos, T.; Oikonomou, A.; Kyriacou, A.; Andriotis, M.; Fragopoulou, E.; Pantazidou, A. Characterization of bioactive glycolipids from Scytonema julianum (cyanobacteria). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 140, 219–231. [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Shiels, K.; Saha, S.K.; Nasopoulou, C.; Zabetakis, I. In Vitro Antithrombotic Properties of Salmon (Salmo salar) Phospholipids in a Novel Food-Grade Extract. Mar. Drugs 2019, 17, 62. [Google Scholar] [CrossRef] [Green Version]
- Tsoupras, A.; Lordan, R.; Demuru, M.; Shiels, K.; Saha, S.K.; Nasopoulou, C.; Zabetakis, I. Structural Elucidation of Irish Organic Farmed Salmon (Salmo salar) Polar Lipids with Antithrombotic Activities. Mar. Drugs 2018, 16, 176. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Hua, X.; Han, F.; Mu, J.; Di, C.; Bo, F.; Zeng, H. Regulation of lipid metabolism in the green microalga Chlorella protothecoides by heterotrophy–photoinduction cultivation regime. Bioresour. Technol. 2015, 192, 781–791. [Google Scholar] [CrossRef]
- Hu, C.-W.; Chuang, L.-T.; Yu, P.-C.; Chen, C.-N.N. Pigment production by a new thermotolerant microalga Coelastrella sp. F50. Food Chem. 2013, 138, 2071–2078. [Google Scholar] [CrossRef]
- Ra, C.H.; Kang, C.-H.; Kim, N.K.; Lee, C.-G.; Kim, S.-K. Cultivation of four microalgae for biomass and oil production using a two-stage culture strategy with salt stress. Renew. Energy 2015, 80, 117–122. [Google Scholar] [CrossRef]
- Mehariya, S.; Sharma, N.; Iovine, A.; Casella, P.; Marino, T.; Larocca, V.; Molino, A.; Musmarra, D. An Integrated Strategy for Nutraceuticals from Haematococcus pluvialis: From Cultivation to Extraction. Antioxidants 2020, 9, 825. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.O.; Han, J.G.; Kim, Y.; Ha, J.H.; Kim, S.S.; Jeong, M.H.; Jeong, H.S.; Kim, N.Y.; Cho, J.S.; Yoon, W.B.; et al. Lipid production in Porphyridium cruentum grown under different culture conditions. J. Biosci. Bioeng. 2009, 108, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Velea, S.; Ilie, L.; Filipescu, L. Optimization of Porphyridium purpureum culture growth using two variables experimental design: Light and sodium bicarbonate. UPB Sci. Bull. 2011, 73, 81–94. [Google Scholar]
- Huo, S.; Basheer, S.; Liu, F.; Elshobary, M.; Zhang, C.; Qian, J.; Xu, L.; Arslan, M.; Cui, F.; Zan, X.; et al. Bacterial intervention on the growth, nutrient removal and lipid production of filamentous oleaginous microalgae Tribonema sp. Algal Res. 2020, 52, 102088. [Google Scholar] [CrossRef]
- Tsoupras, A.; O’Keeffe, E.; Lordan, R.; Redfern, S.; Zabetakis, I. Bioprospecting for Antithrombotic Polar Lipids from Salmon, Herring, and Boarfish By-Products. Foods 2019, 8, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoupras, A.; Lordan, R.; O’Keefe, E.; Shiels, K.; Saha, S.K.; Zabetakis, I. Structural Elucidation of Irish Ale Bioactive Polar Lipids with Antithrombotic Properties. Biomolecules 2020, 10, 1075. [Google Scholar] [CrossRef] [PubMed]
- Vasange, M.; Rolfsen, W.; Bohlin, L. A sulphonoglycolipid from the fern Polypodium decumanum and its effect on the platelet activating-factor receptor in human neutrophils. J. Pharm. Pharmacol. 1997, 49, 562–566. [Google Scholar] [CrossRef]
- Bohlin, L.; Rolfsen, W.; Vasange, M. Use of Sulpho-Quinovosyl-Diacylglycerols for the Treatment of Inflammatory Skin Diseases. EP0952836B1, 28 October 2004. [Google Scholar]
- Berge, J.P.; Debiton, E.; Dumay, J.; Durand, P.; Barthomeuf, C. In vitro anti-inflammatory and anti-proliferative activity of sulfolipids from the red alga Porphyridium cruentum. J. Agric. Food Chem. 2002, 50, 6227–6232. [Google Scholar] [CrossRef]
- Oku, H.; Wongtangtintharn, S.; Iwasaki, H.; Shimatani, M.I.M.; Toda, T. Tumor specific cytotoxicity of glucosylceramide. Cancer Chemother. Pharmacol. 2007, 60, 767–775. [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Harrington, J.; Pienaar, R.; Devaney, K.; Heaney, S.; Koidis, A.; Zabetakis, I. The Effects of Oxidation on the Antithrombotic Properties of Tea Lipids Against PAF, Thrombin, Collagen, and ADP. Foods 2020, 9, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rippka, R.; Stanier, R.Y.; Deruelles, J.; Herdman, M.; Waterbury, J.B. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Lees, M.; Sloan-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissue. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Tsoupras, A.; Zabetakis, I.; Lordan, R. Platelet aggregometry assay for evaluating the effects of platelet agonists and antiplatelet compounds on platelet function in vitro. MethodsX 2018, 6, 63–70. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
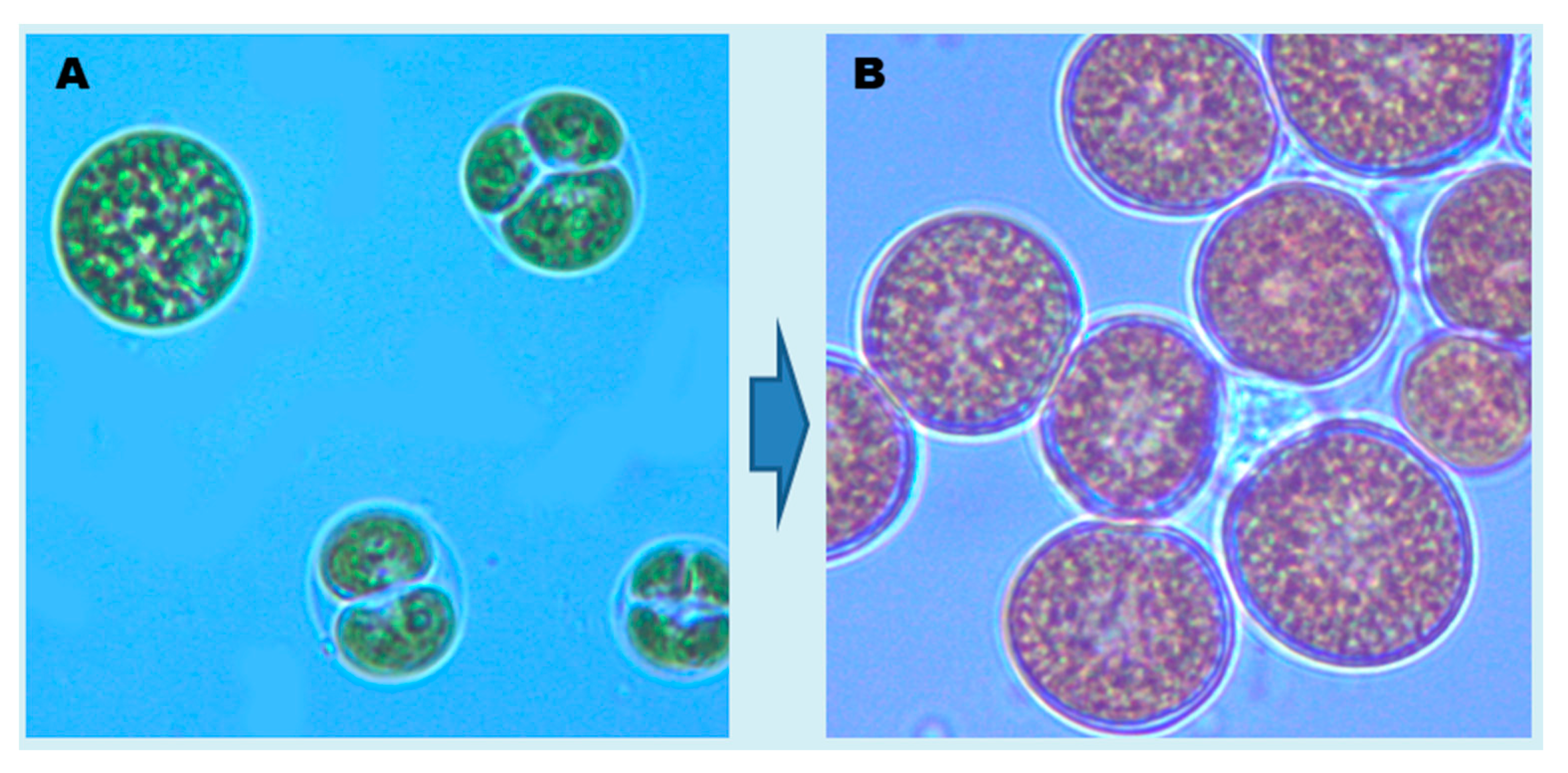{kind=link}
| Microalgal Species | Growth Conditions | Lipid Yield (% Dry Weight Biomass) | References |
|---|---|---|---|
| Chlorella protothecoides | Freshwater, heterotrophy-photoinduction | 50.5 | [23] |
| Chlorococcum sp. RAP13 | Marine, heterotrophic | 38.9 ± 1.9 * | [19] |
| Chlorococcum sp. RAP13 | Marine, photoautotrophic | 20.8 ± 2.6 * | [19] |
| Chlorococcum sp. SABC 012504 | Marine, photoautotrophic | 22 ± 2.52 * | This study |
| Coelastrella sp. F50 | Marine, photoautotrophic | 22 ± 1.7 * | [24] |
| Dunaliella salina | Marine, photoautotrophic, two-stage low-salt stress | 43 | [25] |
| Dunaliella tertiolecta | Marine, photoautotrophic, two-stage low-salt stress | 40 | [25] |
| Haematococcus pluvialis | Freshwater, photoautotrophic | 32–37 | [26] |
| Haematococcus pluvialis SCCAP K-0084 | Freshwater, photoautotrophic | 25–46 ± 1.25–2.3 ** | [18] |
| Isochrysis galbana | Marine, photoautotrophic, two-stage low-salt stress | 47 | [25] |
| Nannochloropsis oculata | Marine, photoautotrophic, two-stage low-salt stress | 29 | [25] |
| Porphyridium cruentum | Marine, photoautotrophic | 19.3 | [27] |
| Porphyridium purpureum | Marine, photoautotrophic | 9–14 | [28] |
| Tribonema sp. | Freshwater, photoautotrophic, bacterial photoautotrophic co-cultivation | 34.67–49.17 | [29] |
| Flow Through (FT) (R %) | Fraction-1 (R %) | Fraction-2 (R %) | Fraction-3 (R %) | |||||
|---|---|---|---|---|---|---|---|---|
| Fatty acids | Unsap | Sap | Unsap | Sap | Unsap | Sap | Unsap | Sap |
| Caprylic (C8:0) | 1.75 | 0.00 | 0.56 | 0.00 | 1.06 | 0.00 | 0.00 | 0.00 |
| Pelargonic (C9:0) | 16.21 | 0.00 | 1.12 | 0.00 | 1.48 | 0.00 | 0.00 | 0.00 |
| Lauric (C12:0) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.05 |
| Myristic (C14:0) | 0.00 | 0.10 | 0.57 | 0.11 | 0.53 | 0.20 | 0.00 | 0.40 |
| Pentadecylic (C15:0) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.15 |
| Palmitic (C16:0 (PA)) | 19.03 | 28.80 | 18.25 | 32.57 | 21.75 | 26.28 | 21.51 | 52.70 |
| Palmitoleic (C16:1) | 0.00 | 0.18 | 0.00 | 1.40 | 0.00 | 2.54 | 0.00 | 0.34 |
| Margaric (C17:0) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.38 | 0.00 | 0.58 |
| Stearic (C18:0) | 63.02 | 41.83 | 68.36 | 25.59 | 75.18 | 21.89 | 78.49 | 23.26 |
| Oleic (C18:1 (OA)) | 0.00 | 22.74 | 9.91 | 21.55 | 0.00 | 20.86 | 0.00 | 13.47 |
| Linoleic (C18:2 (LA)) | 0.00 | 1.64 | 0.00 | 11.76 | 0.00 | 18.30 | 0.00 | 2.72 |
| Linolenic (C18:3 (ALA/GLA)) | 0.00 | 4.23 | 1.24 | 4.83 | 0.00 | 5.05 | 0.00 | 6.13 |
| Stearidonic (C18:4) | 0.00 | 0.15 | 0.00 | 0.74 | 0.00 | 1.11 | 0.00 | 0.19 |
| Gadoleic (C20:1) | 0.00 | 0.34 | 0.00 | 1.45 | 0.00 | 3.35 | 0.00 | 0.00 |
| Dihomolinolenic (C20:3) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.04 | 0.00 | 0.00 |
| Total number of SFAs | 4 | 3 | 5 | 3 | 5 | 4 | 2 | 6 |
| Total number of UFAs | 0 | 6 | 2 | 6 | 0 | 7 | 0 | 5 |
| Total peak area | 1,421,451.59 | 149,087,839.98 | 6,085,702.65 | 284,509,430.70 | 3,285,702.02 | 265,139,703.41 | 1,211,926.85 | 171,329,929.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiels, K.; Tsoupras, A.; Lordan, R.; Nasopoulou, C.; Zabetakis, I.; Murray, P.; Saha, S.K. Bioactive Lipids of Marine Microalga Chlorococcum sp. SABC 012504 with Anti-Inflammatory and Anti-Thrombotic Activities. Mar. Drugs 2021, 19, 28. https://0-doi-org.brum.beds.ac.uk/10.3390/md19010028
Shiels K, Tsoupras A, Lordan R, Nasopoulou C, Zabetakis I, Murray P, Saha SK. Bioactive Lipids of Marine Microalga Chlorococcum sp. SABC 012504 with Anti-Inflammatory and Anti-Thrombotic Activities. Marine Drugs. 2021; 19(1):28. https://0-doi-org.brum.beds.ac.uk/10.3390/md19010028
Chicago/Turabian StyleShiels, Katie, Alexandros Tsoupras, Ronan Lordan, Constantina Nasopoulou, Ioannis Zabetakis, Patrick Murray, and Sushanta Kumar Saha. 2021. "Bioactive Lipids of Marine Microalga Chlorococcum sp. SABC 012504 with Anti-Inflammatory and Anti-Thrombotic Activities" Marine Drugs 19, no. 1: 28. https://0-doi-org.brum.beds.ac.uk/10.3390/md19010028