First Detection of Tetrodotoxins in the Cotylean Flatworm Prosthiostomum trilineatum

Abstract
:1. Introduction
2. Results and Discussion
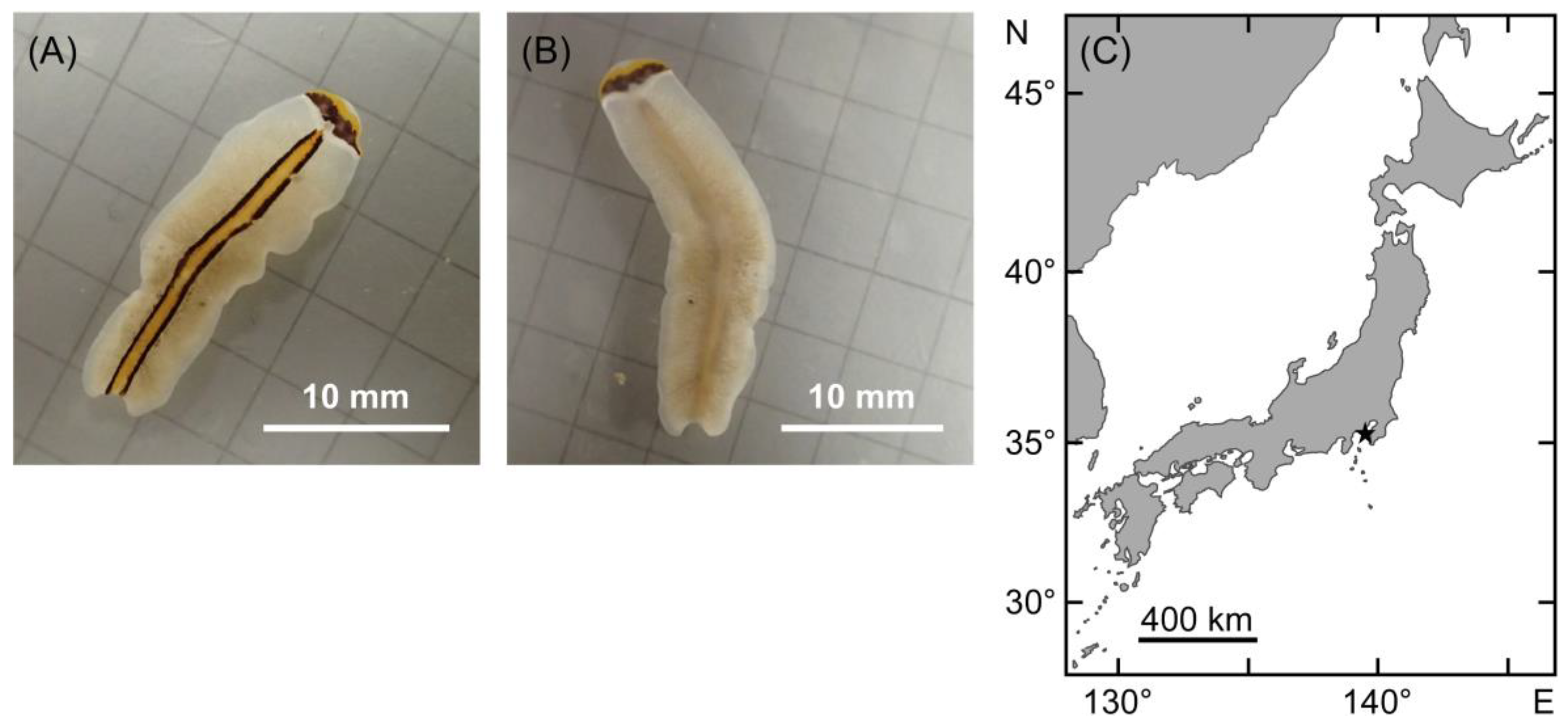
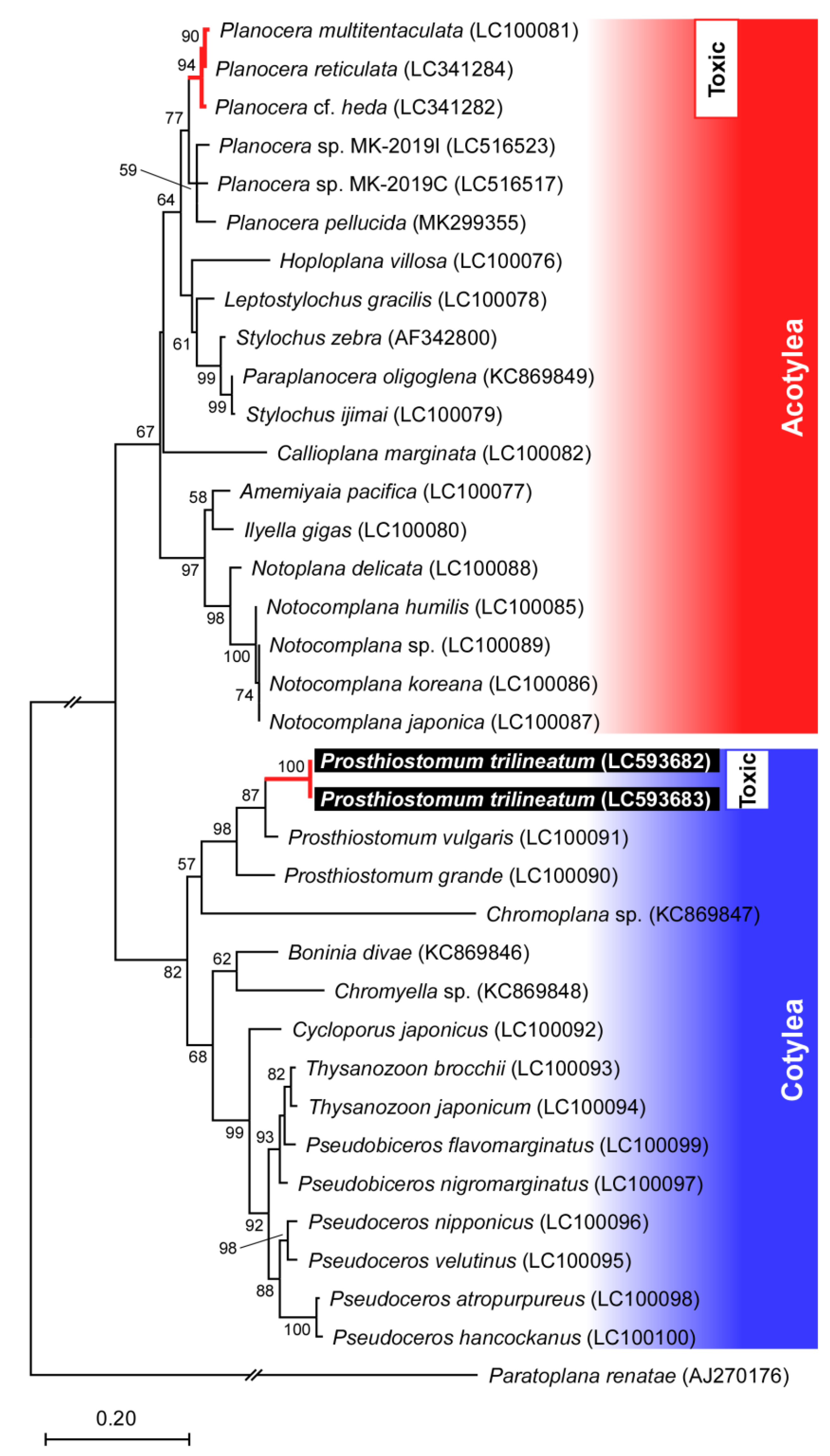
2.1. External Morphology and Molecular Phylogenetic Inference of the Flatworm Pr. trilineatum
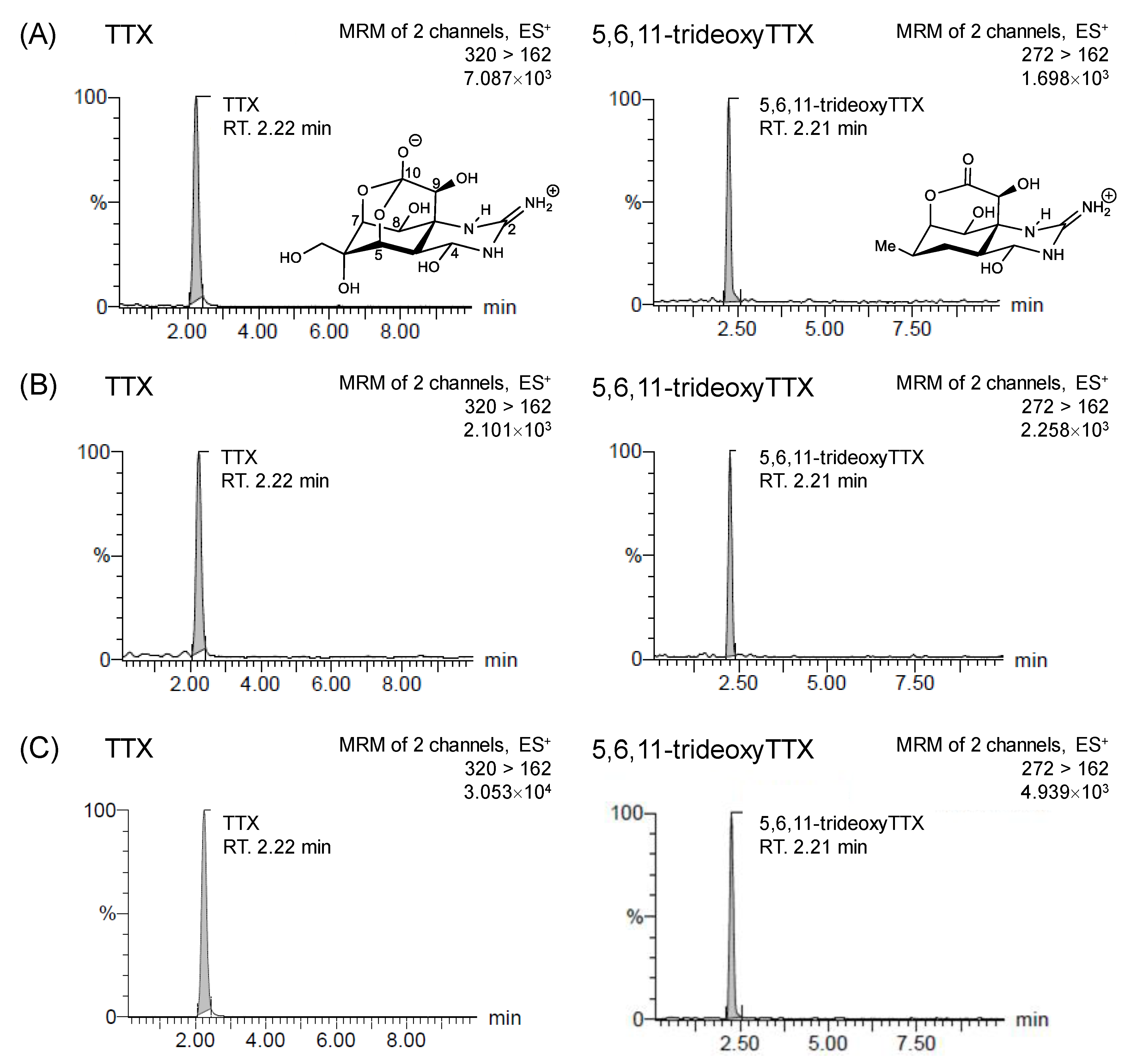
2.2. TTXs Analysis
2.3. Prosthiostomum trilineatum and Possible Relationship with Other Marine Organisms
3. Materials and Methods
3.1. Flatworm Specimens
3.2. Preparation of Sample Solutions of the Flatworm for LC-MS Analysis
3.3. LC-MS/MS Analysis
3.4. Molecular Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsuda, K.; Ikuma, S.; Kawamura, M.; Tachikawa, R.; Sakai, K.; Tamura, C.; Amakasu, O. Tetrodotoxin. VII. On the structures of tetrodotoxin and its derivatives. Chem. Pharm. Bull. 1964, 12, 1357–1374. [Google Scholar] [CrossRef] [Green Version]
- Woodward, R.B. The structure of tetrodotoxin. Pure Appl. Chem. 1964, 86, 49–74. [Google Scholar] [CrossRef]
- Goto, T.; Kishi, Y.; Takahashi, S.; Hirata, Y. Tetrodotoxin. Tetrahedron 1965, 21, 2059–2088. [Google Scholar] [CrossRef]
- Yasumoto, T.; Nagai, H.; Yasumura, D.; Michishita, T.; Endo, A.; Yotsu, M.; Kotaki, Y. Interspecies distribution and possible origin of tetrodotoxin. Ann. N. Y. Acad. Sci. 1986, 479, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. Part D 2006, 1, 145–152. [Google Scholar] [CrossRef]
- Miyazawa, K.; Noguchi, T. Distribution and origin of tetrodotoxin. J. Toxicol. Toxin Rev. 2001, 20, 11–33. [Google Scholar] [CrossRef]
- Narahashi, T. Pharmacology of tetrodotoxin. J. Toxicol. Toxin Rev. 2001, 20, 67–84. [Google Scholar] [CrossRef]
- Colquhon, D.; Henderson, R.; Ritchie, J.M. The binding of labeled tetrodotoxin to non-myelinated nerve fibres. J. Physiol. 1972, 227, 95–126. [Google Scholar] [CrossRef]
- Makarova, M.; Rycek, L.; Hajicek, J.; Baidilov, D.; Hudlicky, T. Tetrodotoxin: History, biology, and synthesis. Angew. Chem. Int. Ed. 2019, 58, 18338–18387. [Google Scholar] [CrossRef]
- Yasumoto, T.; Yasumura, D.; Yotsu, M.; Michishita, T.; Endo, A.; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric. Biol. Chem. 1986, 50, 793–795. [Google Scholar]
- Noguchi, T.; Jeon, J.K.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Occurrence of tetrodotoxin and anhydrotetrodotoxin in Vibrio sp. isolated from the intestines of a xanthid crab, Atergatis floridus. J. Biochem. 1986, 99, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [Green Version]
- Ueyama, N.; Sugimoto, K.; Kudo, Y.; Onodera, K.; Cho, Y.; Konoki, K.; Nishikawa, T.; Yotsu-Yamashita, M. Spiro bicyclic guanidino compounds from pufferfish: Possible biosynthetic intermediates of tetrodotoxin in marine environments. Chem. Eur. J. 2018, 24, 7250–7258. [Google Scholar] [CrossRef] [PubMed]
- Kudo, Y.; Yasumoto, T.; Mebs, D.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Cyclic guanidine compounds from newts support the hypothesis that tetrodotoxin is derived from monoterpene. Angew. Chem. Int. Ed. 2016, 55, 8728–8731. [Google Scholar] [CrossRef] [PubMed]
- McNabb, P.; Selwood, A.I.; Munday, R.; Wood, S.A.; Taylor, D.I.; MacKenzie, L.A.; van Ginkel, R.; Rhodes, L.L.; Cornelisen, C.; Heasman, K.; et al. Detection of tetrodotoxin from the grey side-gilled sea slug—Pleurobranchaea maculata, and associated dog neurotoxicosis on beaches adjacent to the Hauraki Gulf, Auckland, New Zealand. Toxicon 2010, 56, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Salvitti, L.; Wood, S.A.; Taylor, D.I.; McNabb, P.; Cary, S.C. First identification of tetrodotoxin (TTX) in the flatworm Stylochoplana sp.; a source of TTX for the sea slug Pleurobranchaea maculata. Toxicon 2015, 95, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Salvitti, L.R.; Wood, S.A.; Winsor, L.; Cary, S.C. Intracellular immunohistochemical detection of tetrodotoxin in Pleurobranchaea maculata (Gastropoda) and Stylochoplana sp. (Turbellaria). Mar. Drugs 2015, 13, 756–769. [Google Scholar] [CrossRef] [Green Version]
- Itoi, S.; Kozaki, A.; Komori, K.; Tsunashima, T.; Noguchi, S.; Kawane, M.; Sugita, H. Toxic Takifugu pardalis eggs found in Takifugu niphobles gut: Implications for TTX accumulation in the pufferfish. Toxicon 2015, 108, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Itoi, S.; Ueda, H.; Yamada, R.; Takei, M.; Sato, T.; Oshikiri, S.; Wajima, Y.; Ogata, R.; Oyama, H.; Shitto, T.; et al. Including planocerid flatworms in the diet effectively toxifies the pufferfish, Takifugu niphobles. Sci. Rep. 2018, 8, 12302. [Google Scholar] [CrossRef]
- Itoi, S.; Sato, T.; Takei, M.; Yamada, R.; Ogata, R.; Oyama, H.; Teranishi, S.; Kishiki, A.; Wada, T.; Noguchi, K.; et al. The planocerid flatworm is a main supplier of toxin to tetrodotoxin-bearing fish juveniles. Chemosphere 2020, 249, 126217. [Google Scholar] [CrossRef]
- Faubel, A. The Polycladida, Turbellaria. Proposal and establishment of a new system. Part I. The Acotylea. Mitt. Hambg. Zool. Mus. Inst. 1983, 80, 17–121. [Google Scholar]
- Faubel, A. The Polycladida, Turbellaria. Proposal and establishment of a new system. Part II The Cotylea. Mitt. Hambg. Zool. Mus. Inst. 1984, 81, 189–259. [Google Scholar]
- Rawlinson, K.A.; Litvaitis, M.K. Cotylea (Platyhelminthes, Polycladida): A cladistic analysis of morphology. Invertebr. Biol. 2008, 127, 121–138. [Google Scholar] [CrossRef]
- Miyazawa, K.; Jeon, J.K.; Maruyama, J.; Noguchi, T.; Ito, K.; Hashimoto, K. Occurrence of tetrodotoxin in the flatworm Planocera multitentaculata (Platyhelminthes). Toxicon 1986, 24, 645–650. [Google Scholar] [CrossRef]
- Tanu, M.B.; Mahmud, Y.; Arakawa, O.; Takatani, T.; Kajihara, H.; Kawatsu, K.; Hamano, Y.; Asakawa, M.; Miyazawa, K.; Noguchi, T. Immunoenzymatic visualization of tetrodotoxin (TTX) in Cephalothrix species (Nemertea: Anopla: Palaeonemertea: Cephalotrichidae) and Planocera reticulata (Platyhelminthes: Turbellaria: Polycladida: Planoceridae). Toxicon 2004, 44, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Itoi, S.; Sugita, H. TTX-bearing planocerid flatworm (Platyhelminthes: Acotylea) in the Ryukyu Islands, Japan. Mar. Drugs 2018, 16, 37. [Google Scholar] [CrossRef] [Green Version]
- Ritson-Williams, R.; Yotsu-Yamashita, M.; Paul, V.J. Ecological functions of tetrodotoxin in a deadly polyclad flatworm. Proc. Natl. Acad. Sci. USA 2006, 103, 3176–3179. [Google Scholar] [CrossRef] [Green Version]
- Yeri, M.; Kaburaki, T. Notes on two new species of Japanese polyclads. Annot. Zool. Jpn. 1920, 9, 591–598. [Google Scholar]
- Pitale, R.; Bhave, V.; Apte, D. First record of family Prosthiostomidae and Prosthiostomum trilineatum (Platyhelminthes: Polycladida) from the west coast of India. Mar. Biodivers. Rec. 2014, 7, 1–6. [Google Scholar] [CrossRef]
- Nakamura, M.; Yasumoto, T. Tetrodotoxin derivatives in puffer fish. Toxicon 1985, 23, 271–276. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Schimmele, B.; Yasumoto, T. Isolation and structural assignment of 5-deoxytetrodotoxin from the puffer fish Fugu poecilonotus. Biosci. Biotechnol. Biochem. 1999, 63, 961–963. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Abe, Y.; Kudo, Y.; Ritson-Williams, R.; Paul, V.J.; Konoki, K.; Cho, Y.; Adachi, M.; Imazu, T.; Nishikawa, T.; et al. First identification of 5,11-dideoxytetrodotoxin in marine animals, and characterization of major fragment ions of tetrodotoxin and its analogs by high resolution ESI-MS/MS. Mar. Drugs 2013, 11, 2799–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasumoto, T.; Yotsu, M.; Murata, M.; Naoki, H. New tetrodotoxin analogue from the newt Cynops ensicauda. J. Am. Chem. Soc. 1988, 110, 2344–2345. [Google Scholar] [CrossRef]
- Jang, J.H.; Yotsu-Yamashita, M. 6,11-DideoxyTTX from the puffer fish, Fugu pardalis. Toxicon 2007, 50, 947–951. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Yamagishi, Y.; Yasumoto, T. 5,6,11-Trideoxytetrodotoxin from the puffer fish, Fugu poecilonotus. Tetrahedron Lett. 1995, 36, 9329–9332. [Google Scholar] [CrossRef]
- Khora, S.S.; Yasumoto, T. Isolation of 11-oxotetrodotoxin from the puffer Arothron nigropunctatus. Tetrahedron Lett. 1989, 30, 4393–4394. [Google Scholar] [CrossRef]
- Endo, A.; Khora, S.S.; Murata, M.; Naoki, H.; Yasumoto, T. Isolation of 11-nortetrodotoxin-6(R)-ol and other tetrodotoxin derivatives from the puffer Fugu niphobles. Tetrahedron Lett. 1988, 29, 4127–4128. [Google Scholar] [CrossRef]
- Yotsu, M.; Hayashi, Y.; Kohra, S.S.; Sato, S.; Yasumoto, T. Isolation and structural assignment of 11-nortetrodotoxin-6(S)-ol from the puffer Arothron nigropunctatus. Biosci. Biotechnol. Biochem. 1992, 56, 370–371. [Google Scholar] [CrossRef] [Green Version]
- Yamada, R.; Tsunashima, T.; Takei, M.; Sato, T.; Wajima, Y.; Kawase, M.; Oshikiri, S.; Kajitani, Y.; Kosoba, K.; Ueda, H.; et al. Seasonal changes in the tetrodotoxin content of the flatworm Planocera multitentaculata. Mar. Drugs 2017, 15, 56. [Google Scholar] [CrossRef] [Green Version]
- Itoi, S.; Tabuchi, S.; Abe, M.; Ueda, H.; Oyama, H.; Ogata, R.; Okabe, T.; Kishiki, A.; Sugita, H. Difference in tetrodotoxin content between two sympatric planocerid flatworms, Planocera multitentaculata and Planocera reticulata. Toxicon 2020, 173, 57–61. [Google Scholar] [CrossRef]
- Kashitani, M.; Okabe, T.; Oyama, H.; Noguchi, K.; Yamazaki, H.; Suo, R.; Mori, T.; Sugita, H.; Itoi, S. Taxonomic distribution of tetrodotoxin in acotylean flatworms (Polycladida: Platyhelminthes). Mar. Biotechnol. 2020, 22, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Newman, L.J.; Cannon, L.R.G. Marine Flatworms: The World of Polyclads; CSIRO Publishing: Collingwood, Australia, 2003. [Google Scholar]
- Newman, L.J.; Cannon, L.R.G. Fabulous Flatworms: A Guide to Marine Polyclads; CSIRO Publishing: Collingwood, Australia, 2005. [Google Scholar]
- Okabe, T.; Oyama, H.; Kashitani, M.; Ishimaru, Y.; Suo, R.; Sugita, H.; Itoi, S. Toxic flatworm egg plates serve as a possible source of tetrodotoxin for pufferfish. Toxins 2019, 11, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawlinson, K.A. The diversity, development and evolution of polyclad flatworm larvae. EvoDevo 2014, 5, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, P.; Alfonso, A.; Otero, P.; Katikou, P.; Georgantelis, D.; Botana, L.M. Liquid chromatography–mass spectrometry method to detect tetrodotoxin and its analogues in the puffer fish Lagocephalus sceleratus (Gmelin, 1789) from European waters. Food Chem. 2012, 132, 1103–1111. [Google Scholar] [CrossRef]
- Kosker, A.R.; Ozogul, F.; Ayas, D.; Durmus, M.; Ucar, Y.; Regenstein, J.M.; Ozogul, Y. Tetrodotoxin levels of three pufferfish species (Lagocephalus sp.) caught in the North-Eastern Mediterranean sea. Chemosphere 2019, 219, 95–99. [Google Scholar] [CrossRef]
- Katikou, P.; Georgantelis, D.; Sinouris, N.; Petsi, A.; Fotaras, T. First report on toxicity assessment of the Lessepsian migrant pufferfish Lagocephalus sceleratus (Gmelin, 1789) from European waters (Aegean Sea, Greece). Toxicon 2009, 54, 50–55. [Google Scholar] [CrossRef]
- McNabb, P.S.; Taylor, D.I.; Ogilvie, S.C.; Wilkinson, L.; Anderson, A.; Hamon, D.; Wood, S.A.; Peake, B.M. First detection of tetrodotoxin in the bivalve Paphies australis by liquid chromatography coupled to triple quadrupole mass spectrometry with and without precolumn reaction. J. AOAC Int. 2014, 97, 325–333. [Google Scholar] [CrossRef]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Eurosurveillance 2015, 20, 2–8. [Google Scholar] [CrossRef]
- Turner, A.D.; Dhanji-Rapkova, M.; Coates, L.; Bickersta, L.; Milligam, S.; O’Neill, A.; Faulkner, D.; McEneny, H.; Baker-Austin, C.; Lees, D.N.; et al. Detection of tetrodotoxin shellfish poisoning (TSP) toxins and causative factors in bivalve molluscs from the UK. Mar. Drugs 2017, 15, 277. [Google Scholar] [CrossRef] [Green Version]
- Gerssen, A.; Bovee, T.; Klijnstra, M.; Poelman, M.; Portier, L.; Hoogenboom, R. First report on the occurrence of tetrodotoxins in bivalve mollusks in the Netherlands. Toxins 2018, 10, 450. [Google Scholar] [CrossRef] [Green Version]
- Dell’Aversano, C.; Tartaglione, L.; Polito, G.; Dean, K.; Giacobbe, M.; Casabianca, S.; Capellacci, S.; Penna, A.; Turner, A.D. First detection of tetrodotoxin and high levels of paralytic shellfish poisoning toxins in shellfish from Sicily (Italy) by three different analytical methods. Chemosphere 2019, 215, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Biessy, L.; Boundy, M.J.; Smith, K.F.; Harwood, D.T.; Hawes, I.; Wood, S.A. Tetrodotoxin in marine bivalves and edible gastropods: A mini-review. Chemosphere 2019, 236, 124404. [Google Scholar] [CrossRef] [PubMed]
- Adachi, M.; Sakakibara, R.; Satake, Y.; Isobe, M.; Nishikawa, T. Synthesis of 5,6,11-trideoxytetrodotoxin. Chem. Lett. 2014, 43, 1719–1721. [Google Scholar] [CrossRef]
- Tsunashima, T.; Hagiya, M.; Yamada, R.; Koito, T.; Tsuyuki, N.; Izawa, S.; Kosoba, K.; Itoi, S.; Sugita, H. A molecular framework for the taxonomy and systematics of Japanese marine turbellarian flatworms (Platyhelminthes, Polycladida). Aquat. Biol. 2017, 26, 159–167. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Compounds | Content (μg/individual) | |
|---|---|---|
| Specimen 1 | Specimen 2 | |
| TTX | 223 | 8.4 |
| 5,6,11-trideoxyTTX | 170 | 6.2 |
| Compounds | Transition | Cone (V) | CE (V) |
|---|---|---|---|
| TTX | 320 > 302 | 42 | 24 |
| 320 > 162 | 42 | 38 | |
| 5,6,11-trideoxyTTX | 272 > 254 | 46 | 19 |
| 272 > 162 | 46 | 28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suo, R.; Kashitani, M.; Oyama, H.; Adachi, M.; Nakahigashi, R.; Sakakibara, R.; Nishikawa, T.; Sugita, H.; Itoi, S. First Detection of Tetrodotoxins in the Cotylean Flatworm Prosthiostomum trilineatum. Mar. Drugs 2021, 19, 40. https://0-doi-org.brum.beds.ac.uk/10.3390/md19010040
Suo R, Kashitani M, Oyama H, Adachi M, Nakahigashi R, Sakakibara R, Nishikawa T, Sugita H, Itoi S. First Detection of Tetrodotoxins in the Cotylean Flatworm Prosthiostomum trilineatum. Marine Drugs. 2021; 19(1):40. https://0-doi-org.brum.beds.ac.uk/10.3390/md19010040
Chicago/Turabian StyleSuo, Rei, Maho Kashitani, Hikaru Oyama, Masaatsu Adachi, Ryota Nakahigashi, Ryo Sakakibara, Toshio Nishikawa, Haruo Sugita, and Shiro Itoi. 2021. "First Detection of Tetrodotoxins in the Cotylean Flatworm Prosthiostomum trilineatum" Marine Drugs 19, no. 1: 40. https://0-doi-org.brum.beds.ac.uk/10.3390/md19010040