Research Progress in Anti-Inflammatory Bioactive Substances Derived from Marine Microorganisms, Sponges, Algae, and Corals


Abstract
:1. Introduction
2. Inflammatory Pathways and Evaluation Model of Anti-Inflammatory Activity
2.1. Inflammatory Pathways
2.2. Evaluation Model of Anti-Inflammatory Activity
3. Anti-Inflammatory Bioactive Substances Derived from Marine Organisms
3.1. Marine Bacteria and Fungi
3.1.1. Anti-Inflammatory Peptides from Marine Bacteria and Fungi
3.1.2. Anti-Inflammatory Polyketides from Marine Bacteria and Fungi
3.1.3. Other Anti-Inflammatory Substances from Marine Bacteria and Fungi
3.2. Marine Sponges
3.2.1. Anti-Inflammatory Peptides from Marine Sponge
3.2.2. Anti-Inflammatory Terpenoids from Marine Sponge
3.2.3. Other Anti-Inflammatory Substances from Marine Sponge
3.3. Marine Algae
3.3.1. Anti-Inflammatory Peptides and Proteins from Marine Algae
3.3.2. Anti-Inflammatory Polysaccharides from Marine Algae
3.3.3. Other Anti-Inflammatory Substances from Marine Algae
3.4. Marine Corals
3.4.1. Anti-Inflammatory Terpenoids from Marine Corals
3.4.2. Other Anti-Inflammatory Substances from Marine Corals
4. Conclusions and Research Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Ricordi, C.; Garcia-Contreras, M.; Farnetti, S. Diet and Inflammation: Possible Effects on Immunity, Chronic Diseases, and Life Span. J. Am. Coll. Nutr. 2015, 34, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Ahluwalia, N.; Albers, R.; Bosco, N.; Bourdet-Sicard, R.; Haller, D.; Holgate, S.T.; Jonsson, L.S.; Latulippe, M.E.; Marcos, A.; et al. A Consideration of Biomarkers to be used for Evaluation of Inflammation in Human Nutritional Studies. Br. J. Nutr. 2013, 109, S1–S34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, M.-H.; Lai, C.-S.; Ho, C.-T. Anti-inflammatory activity of natural dietary flavonoids. Food Funct. 2010, 1, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P. The inflammation theory of disease—The growing realization that chronic inflammation is crucial in many diseases opens new avenues for treatment. EMBO Rep. 2012, 13, 968–970. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.-W.; Karin, M. A cytokine-mediated link between innate immunity, inflammation, and cancer. J. Clin. Investig. 2007, 117, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Luk, H.K.H.; Li, X.; Fung, J.; Lau, S.K.P.; Woo, P.C.Y. Molecular epidemiology, evolution and phylogeny of SARS coronavirus. Infect. Genet. Evol. 2019, 71, 21–30. [Google Scholar] [CrossRef]
- Turnquist, C.; Ryan, B.M.; Horikawa, I.; Harris, B.T.; Harris, C.C. Cytokine Storms in Cancer and COVID-19. Cancer Cell 2020, 38, 598–601. [Google Scholar] [CrossRef]
- Kapoor, S.; Nailwal, N.; Kumar, M.; Barve, K. Recent Patents and Discovery of Anti-inflammatory Agents from Marine Source. Recent Pat. Inflamm. Allergy Drug Discov. 2019, 13, 105–114. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Meng, L.-H.; Wang, B.-G. Progress in Research on Bioactive Secondary Metabolites from Deep-Sea Derived Microorganisms. Mar. Drugs 2020, 18, 614. [Google Scholar] [CrossRef]
- Skropeta, D.; Wei, L. Recent advances in deep-sea natural products. Nat. Prod. Rep. 2014, 31, 999–1025. [Google Scholar] [CrossRef]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H.; Chen, Y.; Chan, W.Y. Marine natural products with anti-inflammatory activity. Appl. Microbiol. Biotechnol. 2016, 100, 1645–1666. [Google Scholar] [CrossRef] [PubMed]
- DiDonato, J.A.; Mercurio, F.; Karin, M. NF-kappa B and the link between inflammation and cancer. Immunol. Rev. 2012, 246, 379–400. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-kappa B in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeckinghaus, A.; Hayden, M.S.; Ghosh, S. Crosstalk in NF-kappa B signaling pathways. Nat. Immunol. 2011, 12, 695–708. [Google Scholar] [CrossRef]
- Ahmad, R.; Rasheed, Z.; Ahsan, H. Biochemical and cellular toxicology of peroxynitrite: Implications in cell death and autoimmune phenomenon. Immunopharmacol. Immunotoxicol. 2009, 31, 388–396. [Google Scholar] [CrossRef]
- Zhang, J.-M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohe, R.; Flohe, L. Basic Principles and Emerging Concepts in the Redox Control of Transcription Factors. Antioxid. Redox Signal. 2011, 15, 2335–2381. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Liang, X.; Shaikh, A.S.; Zang, J.; Xu, W.; Zhang, Y. JAK/STAT Signal Transduction: Promising Attractive Targets for Immune, Inflammatory and Hematopoietic Diseases. Curr. Drug Targets 2018, 19, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Greenhill, C.J.; Rose-John, S.; Lissilaa, R.; Ferlin, W.; Ernst, M.; Hertzog, P.J.; Mansell, A.; Jenkins, B.J. IL-6 Trans-Signaling Modulates TLR4-Dependent Inflammatory Responses via STAT3. J. Immunol. 2011, 186, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Chen, J.; Wang, L.; Ivashkiv, L.B. Crosstalk among Jak-STAT, toll-like receptor, and ITAM-dependent pathways in macrophage activation. J. Leukocyte Biol. 2007, 82, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Kamezaki, K.; Shimoda, K.; Numata, A.; Matsuda, T.; Nakayama, K.; Harada, M. The role of Tyk2, Stat1 and Stat4 in LPS-induced endotoxin signals. Int. Immunol. 2004, 16, 1173–1179. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Ni, R.; Jia, W.-Q.; Wang, Y.-Y. IL-4R suppresses airway inflammation in bronchial asthma by inhibiting the IL-4/STAT6 pathway. Pulm. Pharmacol. Ther. 2017, 43, 32–38. [Google Scholar] [CrossRef]
- Arbabi, S.; Maier, R.V. Mitogen-activated protein kinases. Crit. Care Med. 2002, 30, S74–S79. [Google Scholar] [CrossRef] [Green Version]
- Ohkura, T.; Yoshimura, T.; Fujisawa, M.; Ohara, T.; Marutani, R.; Usami, K.; Matsukawa, A. Spred2 Regulates High Fat Diet-Induced Adipose Tissue Inflammation, and Metabolic Abnormalities in Mice. Front. Immunol. 2019, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Sui, X.; Kong, N.; Ye, L.; Han, W.; Zhou, J.; Zhang, Q.; He, C.; Pan, H. p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents. Cancer Lett. 2014, 344, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Dent, P.; Yacoub, A.; Fisher, P.B.; Hagan, M.P.; Grant, S. MAPK pathways in radiation responses. Oncogene 2003, 22, 5885–5896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Khymenets, O.; Urpi-Sarda, M.; Tulipani, S.; Garcia-Aloy, M.; Monagas, M.; Mora-Cubillos, X.; Llorach, R.; Andres-Lacueva, C. Cocoa Polyphenols and Inflammatory Markers of Cardiovascular Disease. Nutrients 2014, 6, 844–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seimi, C.; Cocchi, C.A.; Lanfredini, M.; Keen, C.L.; Gershwin, M.E. Chocolate at heart: The anti-inflammatory impact of cocoa flavanols. Mol. Nutr. Food Res. 2008, 52, 1340–1348. [Google Scholar] [CrossRef]
- Andreyev, A.Y.; Fahy, E.; Guan, Z.; Kelly, S.; Li, X.; McDonald, J.G.; Milne, S.; Myers, D.; Park, H.; Ryan, A.; et al. Subcellular organelle lipidomics in TLR-4-activated macrophages. J. Lipid Res. 2010, 51, 2785–2797. [Google Scholar] [CrossRef] [Green Version]
- Gomes, B.S.; Neto, B.P.S.; Lopes, E.M.; Cunha, F.V.M.; Araujo, A.R.; Wanderley, C.W.S.; Wong, D.V.T.; Junior, R.C.P.L.; Ribeiro, R.A.; Sousa, D.P.; et al. Anti-inflammatory effect of the monoterpene myrtenol is dependent on the direct modulation of neutrophil migration and oxidative stress. Chem. Biol. Interact. 2017, 273, 73–81. [Google Scholar] [CrossRef]
- Lee, S.-J.; Lim, K.-T. Phytoglycoprotein inhibits interleukin-1 beta and interleukin-6 via p38 mitogen-activated protein kinase in lipopolysaccharide-stimulated RAW 264.7 cells. Naunyn Schmiedeberg’s Arch. Pharmacol. 2008, 377, 45–54. [Google Scholar] [CrossRef]
- Gresa-Arribas, N.; Vieitez, C.; Dentesano, G.; Serratosa, J.; Saura, J.; Sola, C. Modelling Neuroinflammation In Vitro: A Tool to Test the Potential Neuroprotective Effect of Anti-Inflammatory Agents. PLoS ONE 2012, 7, e45227. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.-Q.; Song, Y.-L.; Zhu, Z.-X.; Su, C.; Zhang, X.; Wang, J.; Shi, S.-P.; Tu, P.-F. Anti-inflammatory dimeric furanocoumarins from the roots of Angelica dahurica. Fitoterapia 2015, 105, 187–193. [Google Scholar] [CrossRef]
- Inada, T.; Hirota, K.; Shingu, K. Intravenous anesthetic propofol suppresses prostaglandin E-2 and cysteinyl leukotriene production and reduces edema formation in arachidonic acid-induced ear inflammation. J. Immunotoxicol. 2015, 12, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, M.d.S.; Gomes-Rochette, N.F.; de Oliveira, M.L.M.; Nunes-Pinheiro, D.C.S.; Tome, A.R.; Maia de Sousa, F.Y.; Pinheiro, F.G.M.; Moura, C.F.H.; Miranda, M.R.A.; Mota, E.F.; et al. Anti-inflammatory and wound healing potential of cashew apple juice (Anacardium occidentale L.) in mice. Exp. Biol. Med. 2015, 240, 1648–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, S.; Leghouchi, E.; Soulimani, R.; Bouayed, J. Reduction of paw edema and liver oxidative stress in carrageenan-induced acute inflammation by Lobaria pulmonaria and Parmelia caperata, lichen species, in mice. Int. J. Vitam. Nutr. Res. 2021, 91, 143–151. [Google Scholar] [CrossRef]
- Antoniou, E.; Margonis, G.A.; Angelou, A.; Pikouli, A.; Argiri, P.; Karavokyros, I.; Papalois, A.; Pikoulis, E. The TNBS-induced colitis animal model: An overview. Ann. Med. Surg. 2016, 11, 9–15. [Google Scholar] [CrossRef]
- Perse, M.; Cerar, A. Dextran Sodium Sulphate Colitis Mouse Model: Traps and Tricks. J. Biomed. Biotechnol. 2012, 2012, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marjoram, L.; Bagnat, M. Infection, Inflammation and Healing in Zebrafish: Intestinal Inflammation. Curr. Pathobiol. Rep. 2015, 3, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Brugman, S. The zebrafish as a model to study intestinal inflammation. Dev. Comp. Immunol. 2016, 64, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Hu, W.-P.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2009, 26, 170–244. [Google Scholar] [CrossRef]
- Ibrar, M.; Ullah, M.W.; Manan, S.; Farooq, U.; Rafiq, M.; Hasan, F. Fungi from the extremes of life: An untapped treasure for bioactive compounds. Appl. Microbiol. Biotechnol. 2020, 104, 2777–2801. [Google Scholar] [CrossRef]
- Li, H.; Huang, H.; Hou, L.; Ju, J.; Li, W. Discovery of Antimycin-Type Depsipeptides from a wbl Gene Mutant Strain of Deepsea-Derived Streptomyces somaliensis SCSIO ZH66 and Their Effects on Pro-inflammatory Cytokine Production. Front. Microbiol. 2017, 8, 678. [Google Scholar] [CrossRef]
- Liu, J.; Gu, B.; Yang, L.; Yang, F.; Lin, H. New Anti-inflammatory Cyclopeptides From a Sponge-Derived Fungus Aspergillus violaceofuscus. Front. Chem. 2018, 6, 226. [Google Scholar] [CrossRef]
- Lee, S.; Kim, D.-C.; Park, J.-S.; Son, J.-Y.; Sohn, J.H.; Liu, L.; Che, Y.; Oh, H. Penicillospirone from a marine isolate of Penicillium sp (SF-5292) with anti-inflammatory activity. Bioorg. Med. Chem. Lett. 2017, 27, 3516–3520. [Google Scholar] [CrossRef]
- Du, X.; Liu, D.; Huang, J.; Zhang, C.; Proksch, P.; Lin, W. Polyketide derivatives from the sponge associated fungus Aspergillus europaeus with antioxidant and NO inhibitory activities. Fitoterapia 2018, 130, 190–197. [Google Scholar] [CrossRef]
- Ding, Y.; An, F.; Zhu, X.; Yu, H.; Hao, L.; Lu, Y. Curdepsidones B–G, Six Depsidones with Anti-Inflammatory Activities from the Marine-Derived Fungus Curvularia sp. IFB-Z10. Mar. Drugs 2019, 17, 266. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, L.; Fu, X.; Li, Z.; Guo, L.; Kou, L.; Liu, M.; Xie, Z. (+)- and (-)-actinoxocine, and actinaphthorans A-B, C-ring expansion and cleavage angucyclinones from a marine-derived Streptomyces sp. Org. Chem. Front. 2019, 6, 3925–3928. [Google Scholar] [CrossRef]
- Chen, S.; Liu, Z.; Chen, Y.; Tan, H.; Li, S.; Liu, H.; Zhang, W.; Zhu, S. Highly Substituted Phenol Derivatives with Nitric Oxide Inhibitory Activities from the Deep-Sea-Derived Fungus Trichobotrys effuse FS524. Mar. Drugs 2020, 18, 134. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Xie, C.-L.; Xia, J.-M.; Luo, Z.-H.; Shao, Z.; Yang, X.-W. New anti-inflammatory guaianes from the Atlantic hydrotherm-derived fungus Graphostroma sp MCCC 3A00421. Sci. Rep. 2018, 8, 530. [Google Scholar] [CrossRef] [Green Version]
- Alvarino, R.; Alonso, E.; Lacret, R.; Oves-Costales, D.; Genilloud, O.; Reyes, F.; Alfonso, A.; Botana, L.M. Caniferolide A, a Macrolide from Streptomyces caniferus, Attenuates Neuroinflammation, Oxidative Stress, Amyloid-Beta, and Tau Pathology in Vitro. Mol. Pharm. 2019, 16, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Kang, J.S.; Choi, B.-K.; Lee, H.-S.; Lee, Y.-J.; Lee, J.; Shin, H.J. Phenazine Derivatives with Anti-Inflammatory Activity from the Deep-Sea Sediment-Derived Yeast-Like Fungus Cystobasidium laryngis IV17-028. Mar. Drugs 2019, 17, 482. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Sun, W.; Deng, M.; Zhou, Q.; Wang, J.; Liu, J.; Chen, C.; Qi, C.; Luo, Z.; Xue, Y.; et al. Asperversiamides, Linearly Fused Prenylated Indole Alkaloids from the Marine-Derived Fungus Aspergillus versicolor. J. Org. Chem. 2018, 83, 8483–8492. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Dong, Z.; Qiu, P.; Wang, Q.; Yan, J.; Lu, Y.; Wasu, P.-A.; Hong, K.; She, Z. Two new bioactive steroids from a mangrove-derived fungus Aspergillus sp. Steroids 2018, 140, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Liu, Y.; Li, J.; Huang, X.; Yan, T.; Cao, W.; Liu, H.; Long, Y.; She, Z. Diaporindenes A-D: Four Unusual 2,3-Dihydro-1H-indene Analogues with Anti-inflammatory Activities from the Mangrove Endophytic Fungus Diaporthe sp SYSU-HQ3. J. Org. Chem. 2018, 83, 11804–11813. [Google Scholar] [CrossRef]
- Kita, M.; Gise, B.; Kawamura, A.; Kigoshi, H. Stylissatin A, a cyclic peptide that inhibits nitric oxide production from the marine sponge Stylissa massa. Tetrahedron Lett. 2013, 54, 6826–6828. [Google Scholar] [CrossRef]
- Zhang, M.; Sunaba, T.; Sun, Y.; Sasaki, K.; Isoda, H.; Kigoshi, H.; Kita, M. Anti- inflammatory marine cyclic peptide stylissatin A and its derivatives inhibit differentiation of murine preadipocytes. Chem. Commun. 2019, 55, 5471–5474. [Google Scholar] [CrossRef]
- Zhang, M.; Sunaba, T.; Sun, Y.; Shibata, T.; Sasaki, K.; Isoda, H.; Kigoshi, H.; Kita, M. Acyl-CoA dehydrogenase long chain (ACADL) is a target protein of stylissatin A, an anti-inflammatory cyclic heptapeptide. J. Antibiot. 2020, 73, 589–592. [Google Scholar] [CrossRef]
- Li, J.; Yang, F.; Wang, Z.; Wu, W.; Liu, L.; Wang, S.-P.; Zhao, B.-X.; Jiao, W.-H.; Xu, S.-H.; Lin, H.-W. Unusual anti-inflammatory meroterpenoids from the marine sponge Dactylospongia sp. Org. Biomol. Chem. 2018, 16, 6773–6782. [Google Scholar] [CrossRef]
- Gui, Y.H.; Jiao, W.-H.; Zhou, M.; Zhang, Y.; Zeng, D.-Q.; Zhu, H.-R.; Liu, K.-C.; Sun, F.; Chen, H.-F.; Lin, H.-W. Septosones A-C, in Vivo Anti-inflammatory Meroterpenoids with Rearranged Carbon Skeletons from the Marine Sponge Dysidea septosa. Org. Lett. 2019, 21, 767–770. [Google Scholar] [CrossRef]
- Ciaglia, E.; Malfitano, A.M.; Laezza, C.; Fontana, A.; Nuzzo, G.; Cutignano, A.; Abate, M.; Pelin, M.; Sosa, S.; Bifulco, M.; et al. Immuno-Modulatory and Anti-Inflammatory Effects of Dihydrogracilin A, a Terpene Derived from the Marine Sponge Dendrilla membranosa. Int. J. Mol. Sci. 2017, 18, 1643. [Google Scholar] [CrossRef] [Green Version]
- Jiao, W.-H.; Cheng, B.-H.; Chen, G.-D.; Shi, G.-H.; Li, J.; Hu, T.-Y.; Lin, H.-W. Dysiarenone, a Dimeric C-21 Meroterpenoid with Inhibition of COX-2 Expression from the Marine Sponge Dysidea arenaria. Org. Lett. 2018, 20, 3092–3095. [Google Scholar] [CrossRef]
- Di, X.; Rouger, C.; Hardardottir, I.; Freysdottir, J.; Molinski, T.F.; Tasdemir, D.; Omarsdottir, S. 6-Bromoindole Derivatives from the Icelandic Marine Sponge Geodia barretti: Isolation and Anti-Inflammatory Activity. Mar. Drugs 2018, 16, 437. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; Kim, N.-H.; Lee, S.; Kim, Y.N.; Heo, J.D.; Jeong, E.J.; Rho, J.-R. Deacetylphylloketal, a New Phylloketal Derivative from a Marine Sponge, Genus Phyllospongia, with Potent Anti-Inflammatory Activity in In Vitro Co-Culture Model of Intestine. Mar. Drugs 2019, 17, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhoff, J.F.; Labes, A.; Wiese, J. Bio-mining the microbial treasures of the ocean: New natural products. Biotechnol. Adv. 2011, 29, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Abreu, T.M.; Ribeiro, N.A.; Chaves, H.V.; Bezerra Jorge, R.J.; Bezerra, M.M.; Azul Monteiro, H.S.; Vasconcelos, I.M.; Mota, E.F.; Barros Benevides, N.M. Antinociceptive and Anti-inflammatory Activities of the Lectin from Marine Red Alga Solieria filiformis. Planta Med. 2016, 82, 596–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lino de Queiroz, I.N.; Gomes Quindere, A.L.; Gurgel Rodrigues, J.A.; Oliveira Vanderlei, E.D.S.; Ribeiro, N.A.; da Conceicao Rivanor, R.L.; Ribeiro, K.A.; Coura, C.O.; Alves Pereira, K.M.; Chaves, H.V.; et al. Dual effects of a lectin from the green seaweed Caulerpa cupressoides var. lycopodium on inflammatory mediators in classical models of inflammation. Inflamm. Res. 2015, 64, 971–982. [Google Scholar] [CrossRef]
- Dutot, M.; Grassin-Delyle, S.; Salvator, H.; Brollo, M.; Rat, P.; Fagon, R.; Naline, E.; Devillier, P. A marine-sourced fucoidan solution inhibits Toll-like-receptor-3-induced cytokine release by human bronchial epithelial cells. Int. J. Biol. Macromol. 2019, 130, 429–436. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of Bioactivities of Fucoidan from the Brown Seaweed Fucus vesiculosus L. of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Fernando, I.P.S.; Lee, W.W.; Sanjeewa, K.K.A.; Kim, H.-S.; Lee, D.-S.; Jeon, Y.-J. Isolation and purification of fucoidan fraction in Turbinaria ornata from the Maldives; Inflammation inhibitory potential under LPS stimulated conditions in in-vitro and in-vivo models. Int. J. Biol. Macromol. 2019, 131, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Ananthi, S.; Gayathri, V.; Malarvizhi, R.; Bhardwaj, M.; Vasanthi, H.R. Anti-arthritic potential of marine macroalgae Turbinaria ornata in Complete Freund’s Adjuvant induced rats. Exp. Toxicol. Pathol. 2017, 69, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Ye, B.-R.; Kim, E.-A.; Kim, J.; Kim, M.-S.; Lee, W.W.; Ahn, G.-N.; Kang, N.; Jung, W.-K.; Heo, S.-J. Bis (3-bromo-4,5-dihydroxybenzyl) ether, a novel bromophenol from the marine red alga Polysiphonia morrowii that suppresses LPS-induced inflammatory response by inhibiting ROS-mediated ERK signaling pathway in RAW 264.7 macrophages. Biomed. Pharmacother. 2018, 103, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Zbakh, H.; Talero, E.; Avila, J.; Alcaide, A.; de los Reyes, C.; Zubia, E.; Motilva, V. The Algal Meroterpene 11-Hydroxy-1 ‘-O-Methylamentadione Ameloriates Dextran Sulfate Sodium-Induced Colitis in Mice. Mar. Drugs 2016, 14, 149. [Google Scholar] [CrossRef] [Green Version]
- De los Reyes, C.; Ortega, M.J.; Zbakh, H.; Motilva, V.; Zubia, E. Cystoseira usneoides: A Brown Alga Rich in Antioxidant and Anti-inflammatory Meroditerpenoids. J. Nat. Prod. 2016, 79, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Ko, J.-Y.; Kim, E.-A.; Hwang, E.-K.; Park, C.S.; Lee, J.-S.; Kim, C.-Y.; Lee, H.-S.; Kang, H.-K.; Cha, S.-H.; et al. Identification and large isolation of an anti-inflammatory compound from an edible brown seaweed, Undariopsis peterseniana, and evaluation on its anti-inflammatory effect in in vitro and in vivo zebrafish. J. Appl. Phycol. 2017, 29, 1587–1596. [Google Scholar] [CrossRef]
- Dimou, M.; Ioannou, E.; Daskalaki, M.G.; Tziveleka, L.A.; Kampranis, S.C.; Roussis, V. Disulfides with Anti-inflammatory Activity from the Brown Alga Dictyopteris membranacea. J. Nat. Prod. 2016, 79, 584–589. [Google Scholar] [CrossRef]
- Ali, I.; Manzoor, Z.; Koo, J.-E.; Moon, S.-R.; Byeon, S.-H.; Yoo, E.-S.; Kang, H.-K.; Hyun, J.-W.; Lee, N.-H.; Koh, Y.-S. Monoolein, isolated from Ishige sinicola, inhibits lipopolysaccharide-induced inflammatory response by attenuating mitogen-activated protein kinase and NF-kappa B pathways. Food Sci. Biotechnol. 2017, 26, 507–511. [Google Scholar] [CrossRef]
- Xu, S.-Y.; Huang, X.; Cheong, K.-L. Recent Advances in Marine Algae Polysaccharides: Isolation, Structure, and Activities. Mar. Drugs 2017, 15, 388. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Nath, A.K.; Tang, Y.; Choi, Y.-J.; Debnath, T.; Choi, E.-J.; Kim, E.-K. Investigation of the Anti-Prostate Cancer Properties of Marine-Derived Compounds. Mar. Drugs 2018, 16, 160. [Google Scholar] [CrossRef] [Green Version]
- Vo Thanh, S.; Ton That Huu, D.; Le Ba, V.; Le Canh Viet, C.; Phung Thi Thuy, O.; Ha, H.; Kim, Y.H.; Hoang Le Tuan, A.; Yang, S.Y. Coral and Coral-Associated Microorganisms: A Prolific Source of Potential Bioactive Natural Products. Mar. Drugs 2019, 17, 468. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.-C.; Lin, W.-S.; Peng, B.-R.; Chang, Y.-C.; Fang, L.-S.; Li, G.-Q.; Hwang, T.-L.; Wen, Z.-H.; Sung, P.-J. New Furanocembranoids from Briareum violaceum. Mar. Drugs 2019, 17, 214. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-Y.; Lin, S.-C.; Feng, C.-W.; Chen, P.-C.; Su, Y.-D.; Li, C.-M.; Yang, S.-N.; Jean, Y.-H.; Sung, P.-J.; Duh, C.-Y.; et al. Anti-Inflammatory and Analgesic Effects of the Marine-Derived Compound Excavatolide B Isolated from the Culture-Type Formosan Gorgonian Briareum excavatum. Mar. Drugs 2015, 13, 2559–2579. [Google Scholar] [CrossRef] [Green Version]
- Tseng, W.-R.; Ahmed, A.E.; Huang, C.-Y.; Tsai, Y.-Y.; Tai, C.-J.; Orfali, R.S.; Hwang, T.-L.; Wang, Y.-H.; Dai, C.-F.; Sheu, J.-H. Bioactive Capnosanes and Cembranes from the Soft Coral Klyxum flaccidum. Mar. Drugs 2019, 17, 461. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.F.; Chen, Y.-W.; Huang, C.-Y.; Tseng, Y.-J.; Lin, C.-C.; Dai, C.-F.; Wu, Y.-C.; Sheu, J.-H. Isolation and Structure Elucidation of Cembranoids from a Dongsha Atoll Soft Coral Sarcophyton stellatum. Mar. Drugs 2018, 16, 210. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Liu, X.; Jiang, M.; Luo, G.; Wu, Z.; Chen, B.; Li, J.; Liu, L.; Chen, S. Anti-Inflammatory Cembrane-Type Diterpenoids and Prostaglandins from Soft Coral Lobophytum sarcophytoides. Mar. Drugs 2019, 17, 481. [Google Scholar] [CrossRef] [Green Version]
- Torres-Mendoza, D.; Gonzalez, Y.; Felix Gomez-Reyes, J.; Guzman, H.M.; Luis Lopez-Perez, J.; Gerwick, W.H.; Fernandez, P.L.; Gutierrez, M. Uprolides N, O and P from the Panamanian Octocoral Eunicea succinea. Molecules 2016, 21, 819. [Google Scholar] [CrossRef] [Green Version]
- Lai, K.-H.; You, W.-J.; Lin, C.-C.; El-Shazly, M.; Liao, Z.-J.; Su, J.-H. Anti-Inflammatory Cembranoids from the Soft Coral Lobophytum crassum. Mar. Drugs 2017, 15, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, T.-H.; Sung, C.-S.; Lan, Y.-H.; Wang, Y.-C.; Lu, M.-C.; Wen, Z.-H.; Wu, Y.-C.; Sung, P.-J. New Anti-Inflammatory Cembranes from the Cultured Soft Coral Nephthea columnaris. Mar. Drugs 2015, 13, 3443–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whuang, T.-Y.; Tsai, H.-C.; Su, Y.-D.; Hwang, T.-L.; Sung, P.-J. Sterols from the Octocoral Nephthea columnaris. Mar. Drugs 2017, 15, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-Y.; Tseng, W.-R.; Ahmed, A.E.; Chiang, P.-L.; Tai, C.-J.; Hwang, T.-L.; Dai, C.-F.; Sheu, J.-H. Anti-Inflammatory Polyoxygenated Steroids from the Soft Coral Lobophytum michaelae. Mar. Drugs 2018, 16, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.-W.; Uvarani, C.; Huang, C.-Y.; Hwang, T.-L.; Dai, C.-F.; Sheu, J.-H. New anti-inflammatory tocopherol- derived metabolites from the Taiwanese soft coral Cladiella hirsuta. Bioorg. Med. Chem. Lett. 2015, 25, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Sung, P.-J.; Uvarani, C.; Su, J.-H.; Lu, M.-C.; Hwang, T.-L.; Dai, C.-F.; Wu, S.-L.; Sheu, J.-H. Glaucumolides A and B, Biscembranoids with New Structural Type from a Cultured Soft Coral Sarcophyton glaucum. Sci. Rep. 2015, 5, 15624. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Substances | Species | Model | Activities | Reference |
|---|---|---|---|---|
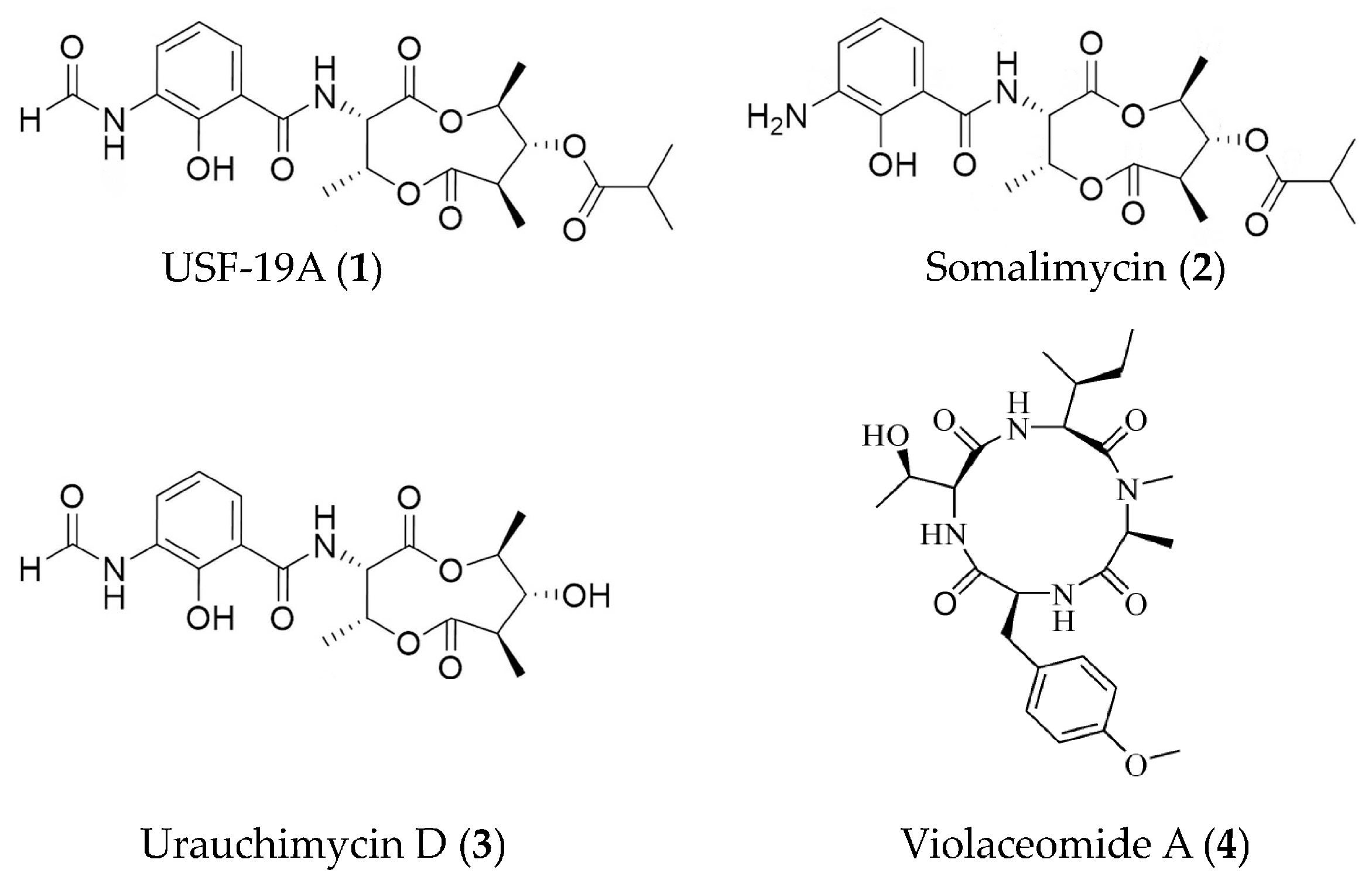
| USF-19A (1), somalimycin (2), and urauchimycin D (3) | Streptomyces somaliensis SCSIO ZH66 | ovalbumin-stimulated mouse splenocytes | against IL-5 with IC50 values of 0.57 μM, > 10 μM and > 10 μM | [44] |
| Violaceomide A (4) | Aspergillus violaceofuscus | LPS-stimulated THP-1 cells | against mRNA expression of IL-10 with inhibitory rate of 84.3% at 10 μM | [45] |
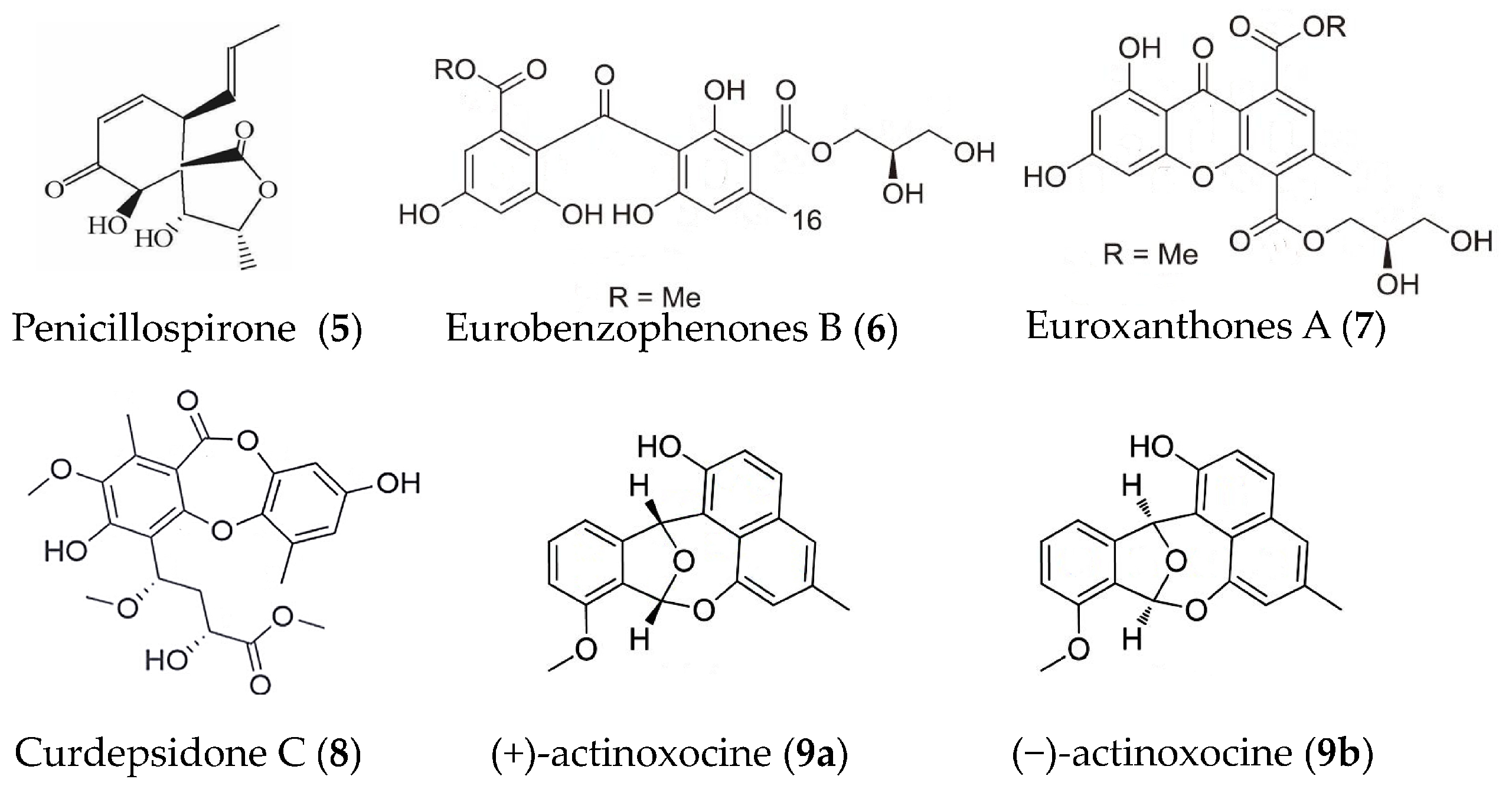
| Penicillospirone (5) | Penicillium sp. SF-5292 | LPS-induced RAW264.7 macrophages and BV2 microglia | against the production of NO, PGE2, TNFα, IL-1β, IL-6, and IL-12 | [46] |
| Eurobenzophenones B (6) euroxanthones A (7) | Aspergillus europaeus | LPS induced BV2 microglia | against NO at 10 μM | [47] |
| Curdepsidone C (8) | Curvularia sp. IFB-Z10 | Propionibacterium acnes-induced THP-1cells | against IL-1β release with an IC50 value of 7.47 ± 0.35 μM | [48] |
| (+)- and (−)-actinoxocine (9a, 9b) | Streptomyces sp. | LPS- and Pam3CSK4-induced RAW 264.7 mouse macrophages | against TNFα protein release | [49] |
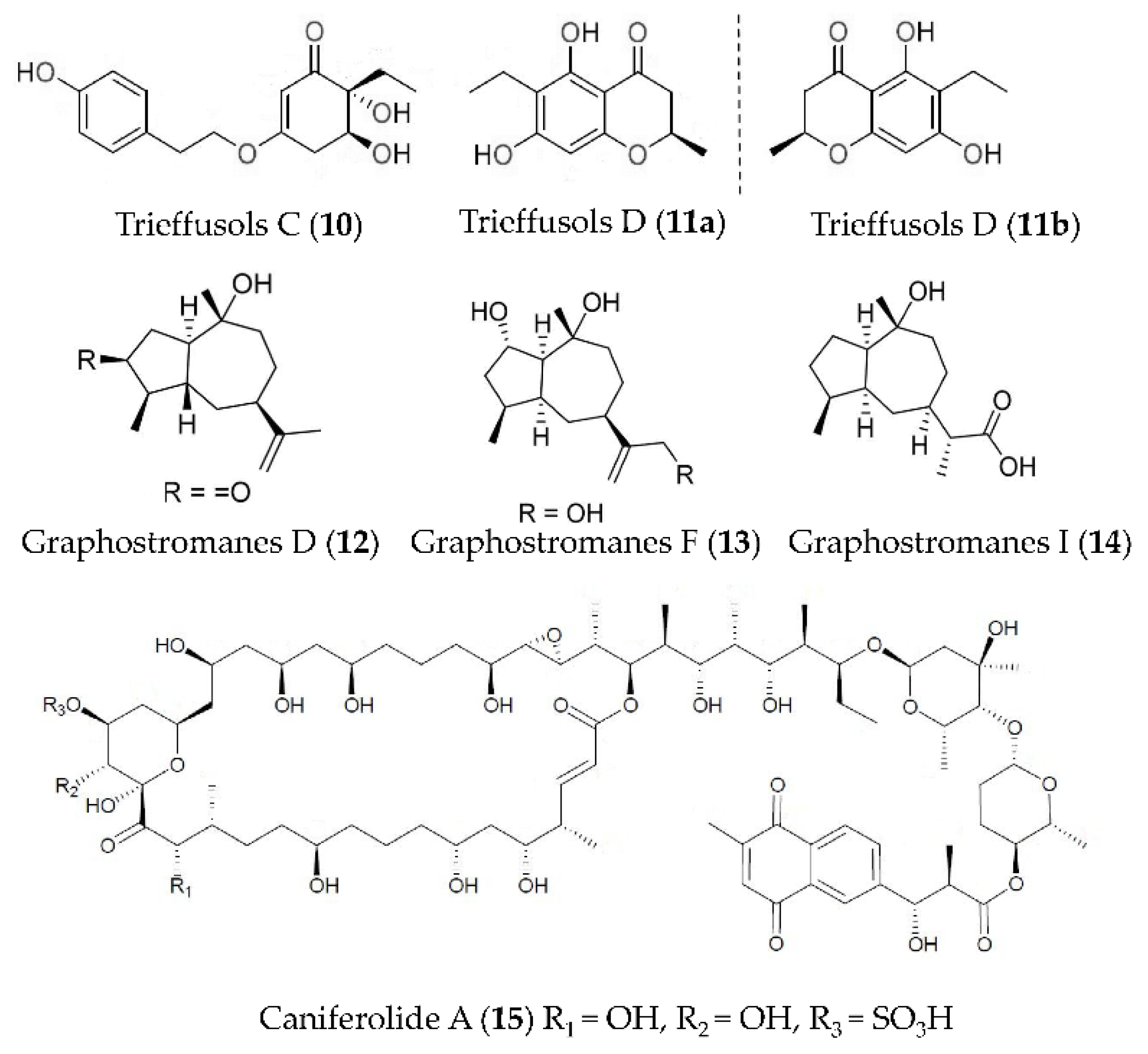
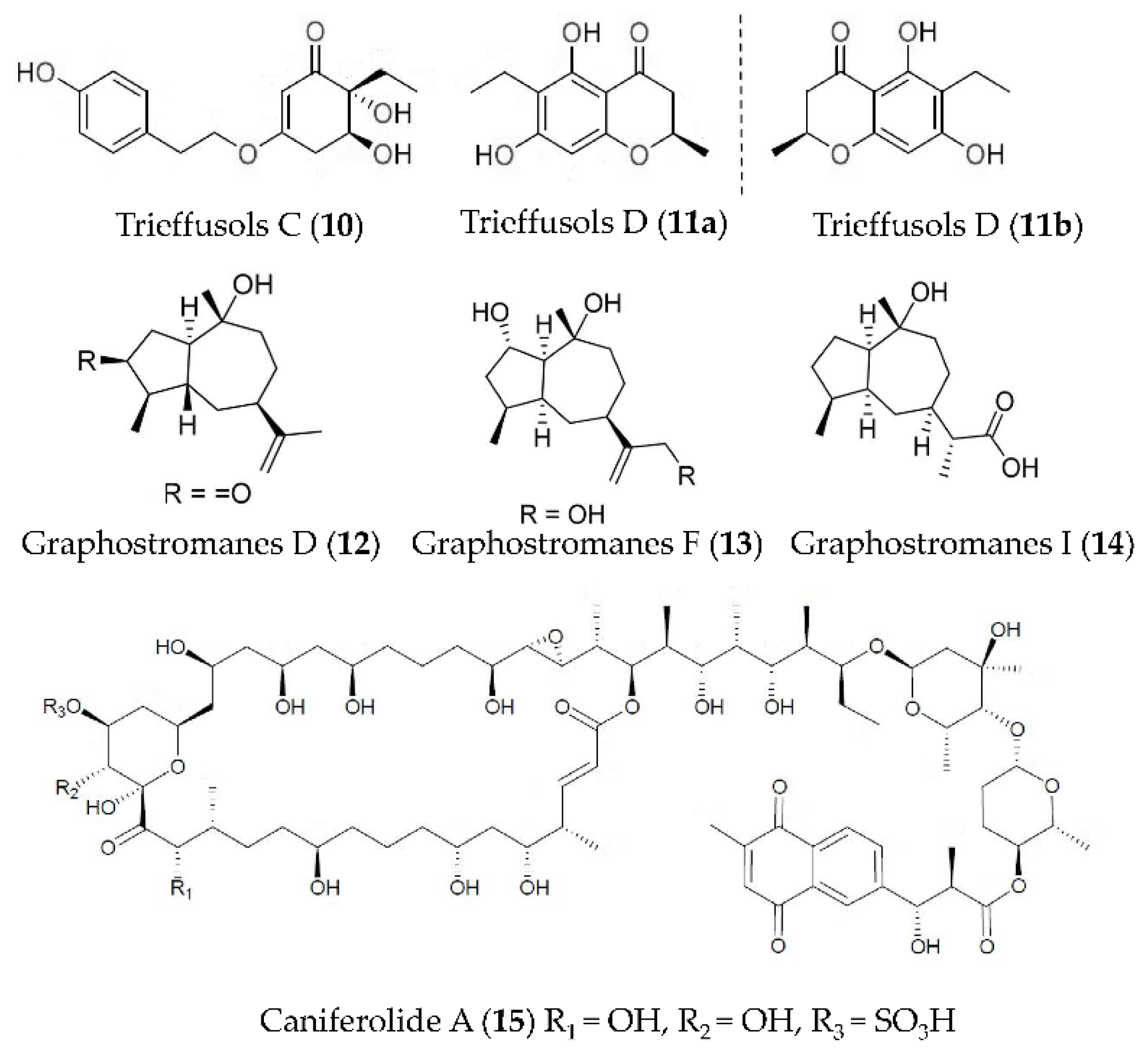
| Trieffusols C and D (10, 11) | Trichobotrys effuse FS524 | LPS-induced RAW264.7 macrophages | against NO with IC50 values ranging from 51.9 to 55.9 μM | [50] |
| Graphostromanes D, F and I (12–14) | Graphostroma sp. MCCC 3A00421 | LPS-induced RAW264.7 macrophages | against NO with IC50 values of 14.2, 72.9 and 88.2 μM | [51] |
| Caniferolide A (15) | Streptomyces caniferus | LPS induced BV2 microglial cells | against NFκBp65 translocation to the nucleus, the production of IL-1β, IL-6 and TNFα, the release of NO, and the activities of iNOS, JNK and p38 | [52] |
| 6-[1-(2-aminobenzoyloxy) ethyl]-1-Phenazinecarboxylic acid (16), Saphenol (17), (R)-saphenic acid (18), Phenazine-1-carboxylic acid (19), 6-(1-hydroxyehtyl) phenazine-1-carboxylic acid (20), 6-acetyl-phenazine-1-carboxylic acid (21) | Cystobasidium larynges IV17-028 | LPS-induced RAW264.7 macrophages | against NO production at 30 μg/mL | [53] |
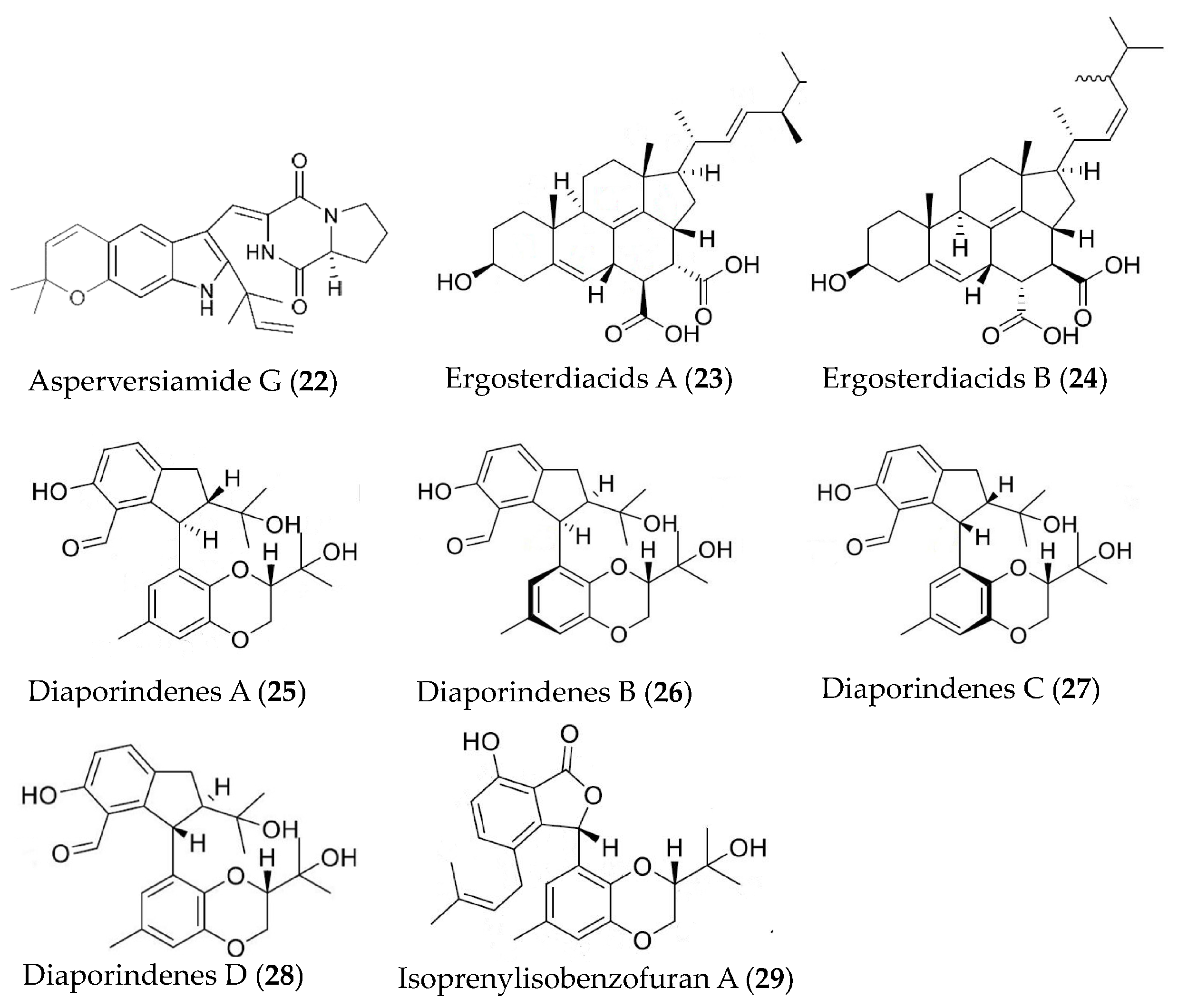
| Asperversiamide G (22) | Aspergillus versicolor | LPS-induced RAW264.7 macrophages | against iNOS with an IC50 value of 5.39 μM | [54] |
| Ergosterdiacids A and B (23, 24) | Aspergillus sp. | LPS-induced RAW264.7 macrophages | against NO with IC50 values of 4.5 and 3.6 μM | [55] |
| Diaporindenes A-D (25–28), isoprenylisobenzofuran A (29) | Diaporthe sp. SYSU-HQ3 | LPS-induced RAW264.7 macrophages | against NO with IC50 values from 4.2 to 9.0 μM | [56] |
| Bioactive Substances | Species | Model | Activities | Reference |
|---|---|---|---|---|
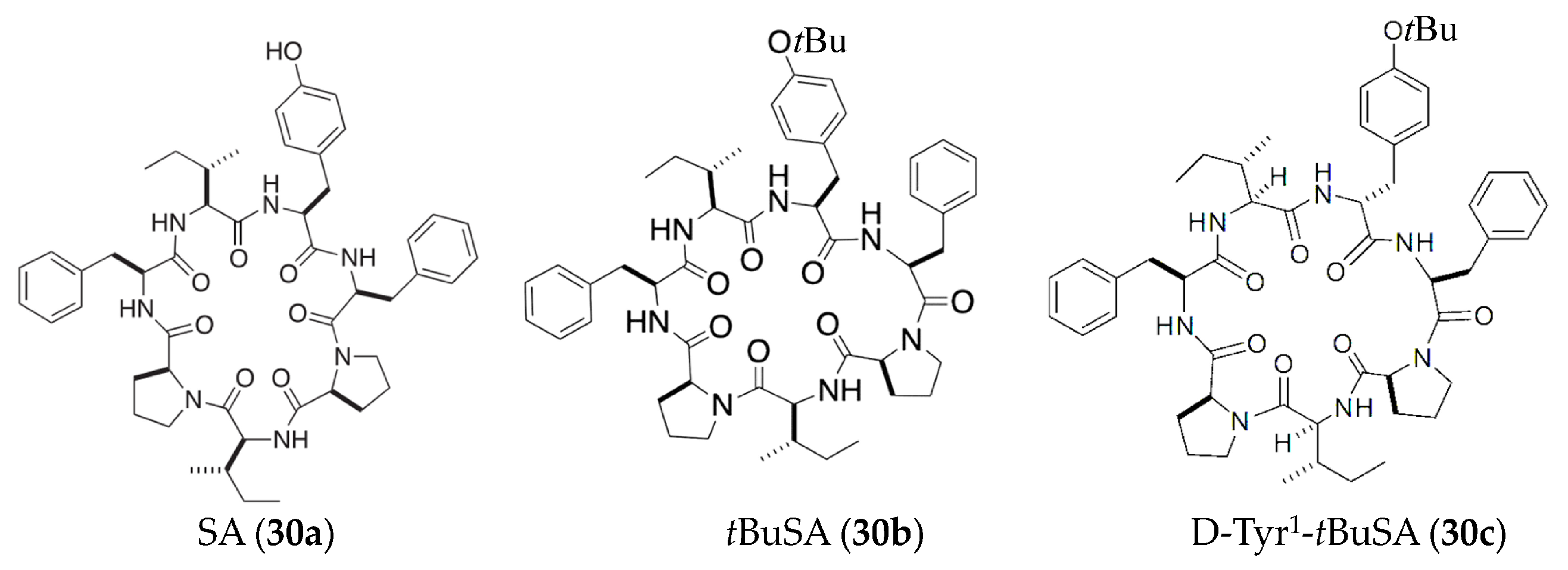
| SA and tBuSA (30a, 30b) | Stylissa massa | LPS-induced RAW264.7 macrophages | against NO with EC50 values of 87 μM | [57,58] |
| D-Tyr1-tBuSA (30c) | Stylissa massa | LPS-induced RAW264.7 macrophages | against production of IL-6 and TNFα (EC50 = 1.4 and 5.9 μM, respectively) and the expression of iNOS (EC50 = 20 μM) | [59] |
| Dactylospongins A and B (31, 32) | Dactylospongia sp. | LPS-stimulated THP-1 cells | against production of IL-6, IL-1β, IL-8, and PGE2 with IC50 values of 5.1–9.2 μM | [60] |
| Septosones A (33) | Dysidea septosa | CuSO4-induced zebrafish; human HEK-293T cells | against migration of macrophages surrounding the neuromast; against TNFα-induced NF-κB activation with IC50 value of 6.8 μM | [61] |
| 9,11-dihydrogracilin A (DHG, 34) | Dendrilla membranosa | Phytohemagglutinin-activated Human peripheral blood mononuclear cells | against production of IL-6 and IL-10 at 3 μM | [62] |
| Dysiarenone (35) | Dysidea arenaria | LPS-induced RAW264.7 macrophages | against COX-2 expression and PGE2 production with IC50 value of 6.4 μM | [63] |
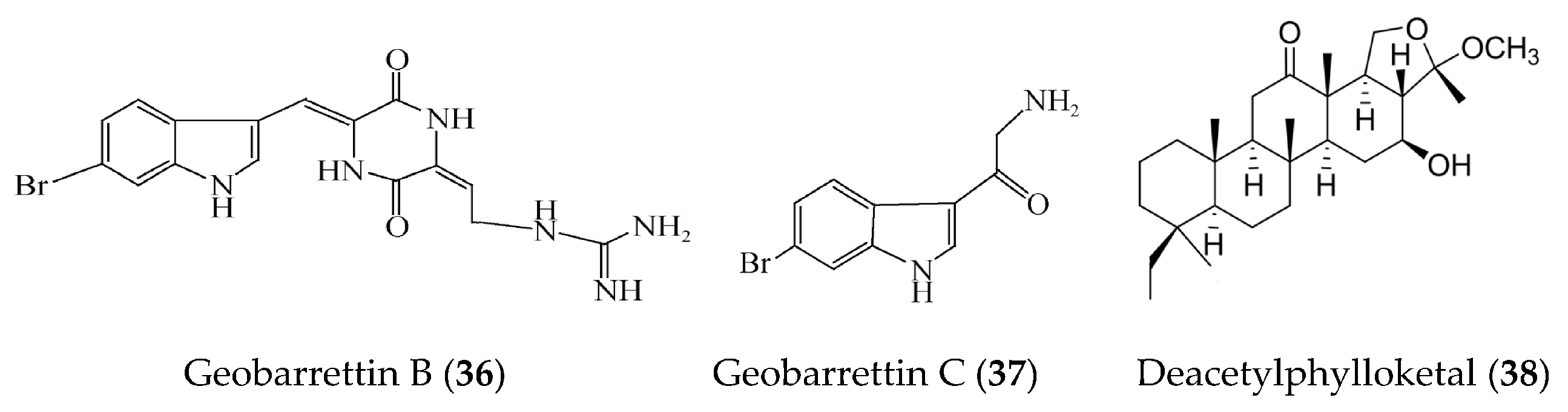
| Geobarrettin B and C (36, 37) | Geodia barretti | LPS-activated DCs | against secretion of IL-10 with inhibitory rate of 29% and 13% at 10 μg/ml | [64] |
| Deacetylphylloketal (38) | Phyllospongia sp. | LPS-induced co-culture system that consisted of human epithelial Caco-2 cells and THP-1 macrophage cells | against production and/or gene expression of NO, PGE2, IL-6, IL-1β, and TNFα, iNOS, and COX-2 | [65] |
| Bioactive Substances | Species | Model | Activities | Reference |
|---|---|---|---|---|
| Lectin | Solieria filiformis | carrageenan-induced peritonitis and paw edema induced by carrageenan, dextran, and serotonin | against neutrophil migration in peritonitis model and decreased paw edema | [67] |
| Lectin | Caulerpa cupressoides | zymosan-induced arthritis of the rat temporomandibular joint | against leukocyte influx and the expression of IL-1β and TNFα at concentrations of 0.1, 1 or 10 mg/kg | [68] |
| Fucoidan | Ascophyllum nodosum | Poly(I:C)-induced human bronchial epithelial cells | against the production of cytokines (IL-1α, IL-1β, TNFα, and IL-6) and PGE2 at the concentration of 0.1% (m/v) | [69] |
| Fucoidan | Fucus vesiculosus L. | LPS-induced human mononuclear U937 cells | against COX-1, COX-2 and hyaluronidase activity with IC50 values of 27, 4.3 and 2.9 μg/mL, and concentration-dependently inhibit the MAPK p38 | [70] |
| Purified fucoidan fraction | Turbinaria ornata | LPS-induced RAW264.7 macrophages and zebrafish embryo | against NO production with IC50 value of 30.83 μg/mL and dose-dependently against iNOS, COX-2, and pro-inflammatory cytokines including PGE2 levels; against production of NO and ROS | [71] |
| Fucoidan like sulphated polysaccharide | Turbinaria ornata | Freud’s adjuvant induced mouse arthritis | against inflammation and bone damage at a low dose of 5 mg/kg | [72] |
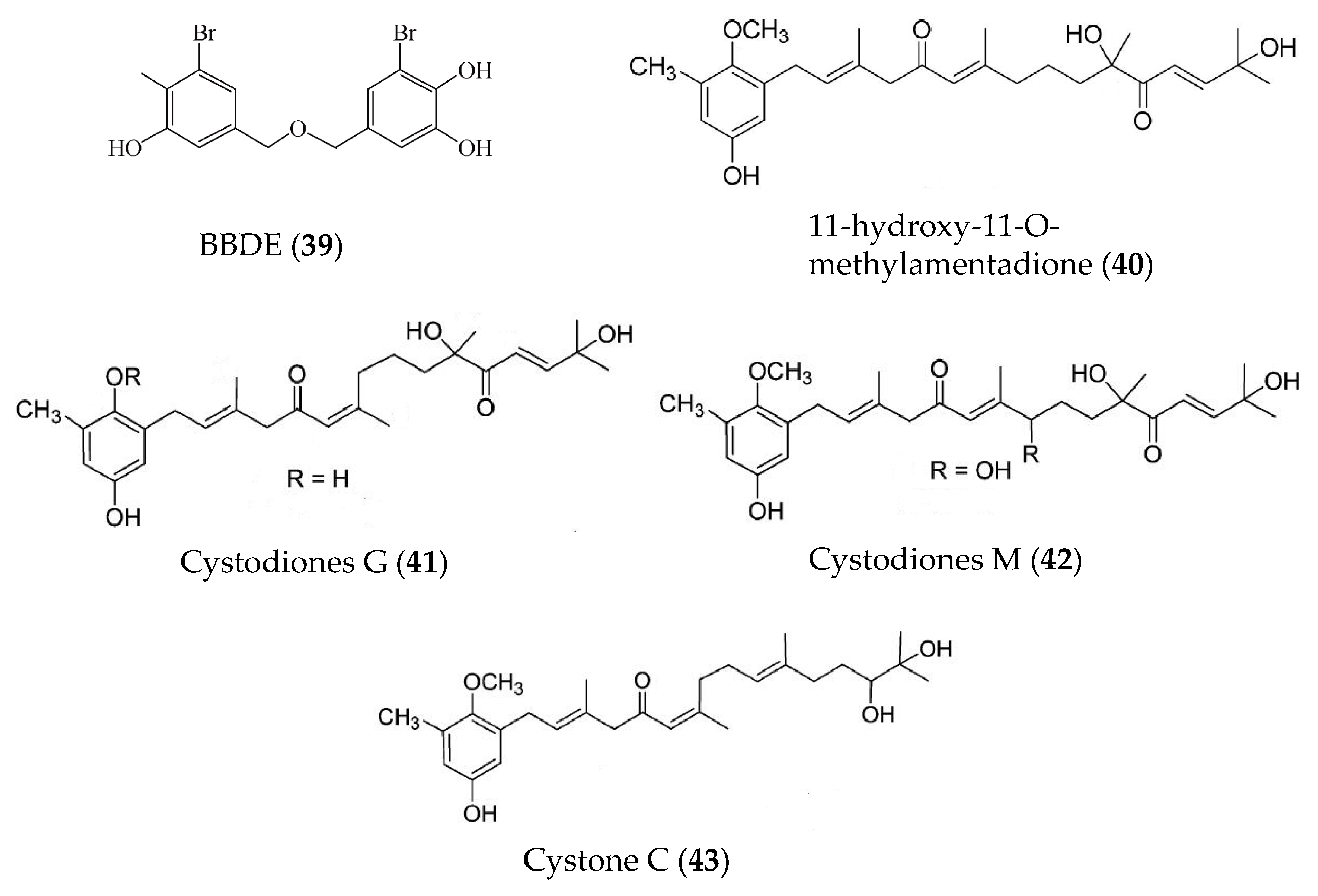
| BBDE (39) | Polysiphonia morrowii | LPS-induced RAW264.7 macrophages | against NO, PGE2, iNOS, COX2, and pro-inflammatory cytokines (TNFα, IL-1β, and IL-6) at 2 μM | [73] |
| 11-hydroxy-1′-O-methylamentadione (40) | Cystoseira usneoides | DSS-induced mouse colitis | Increasing mucus production and against myeloperoxidase activity, production of TNFα, IL-1β and IL-10, and expression of COX-2 and iNOS | [74] |
| Cystodiones G and M (41, 42), cystone C (43) | Cystoseira usneoides | LPS-stimulated THP-1 human macrophages | against the production of TNFα at concentrations of 10, 8 and 5 μM | [75] |
| Apo-9′-fucoxanthinone (44) | Undariopsis peterseniana | LPS-stimulated RAW 264.7 cells; LPS-stimulated zebrafish embryos | against NO, PGE2, iNOS and COX-2, and pro-inflammatory cytokines (TNFα, IL-6, and IL-1β); against inflammatory stress and expression of COX-2 and iNOS | [76] |
| Disulfide (45) | Dictyopteris membranacea | LPS-induced RAW264.7 macrophages | against NO with IC50 value of 3.8 µM | [77] |
| Monoolein (46) | Ishige sinicola | LPS-stimulated primary murine bone marrow-derived dendritic cells | against IL-12 p40, IL-6, and TNFα production with IC50 values of 1.69, 6.87, and 5.19 μM; against the activation of MAPK and NF-κB pathways by inhibiting the phosphorylation of p38, ERK1/2, JNK1/2, and IκBα | [78] |
| Bioactive Substances | Species | Model | Activities | Reference |
|---|---|---|---|---|
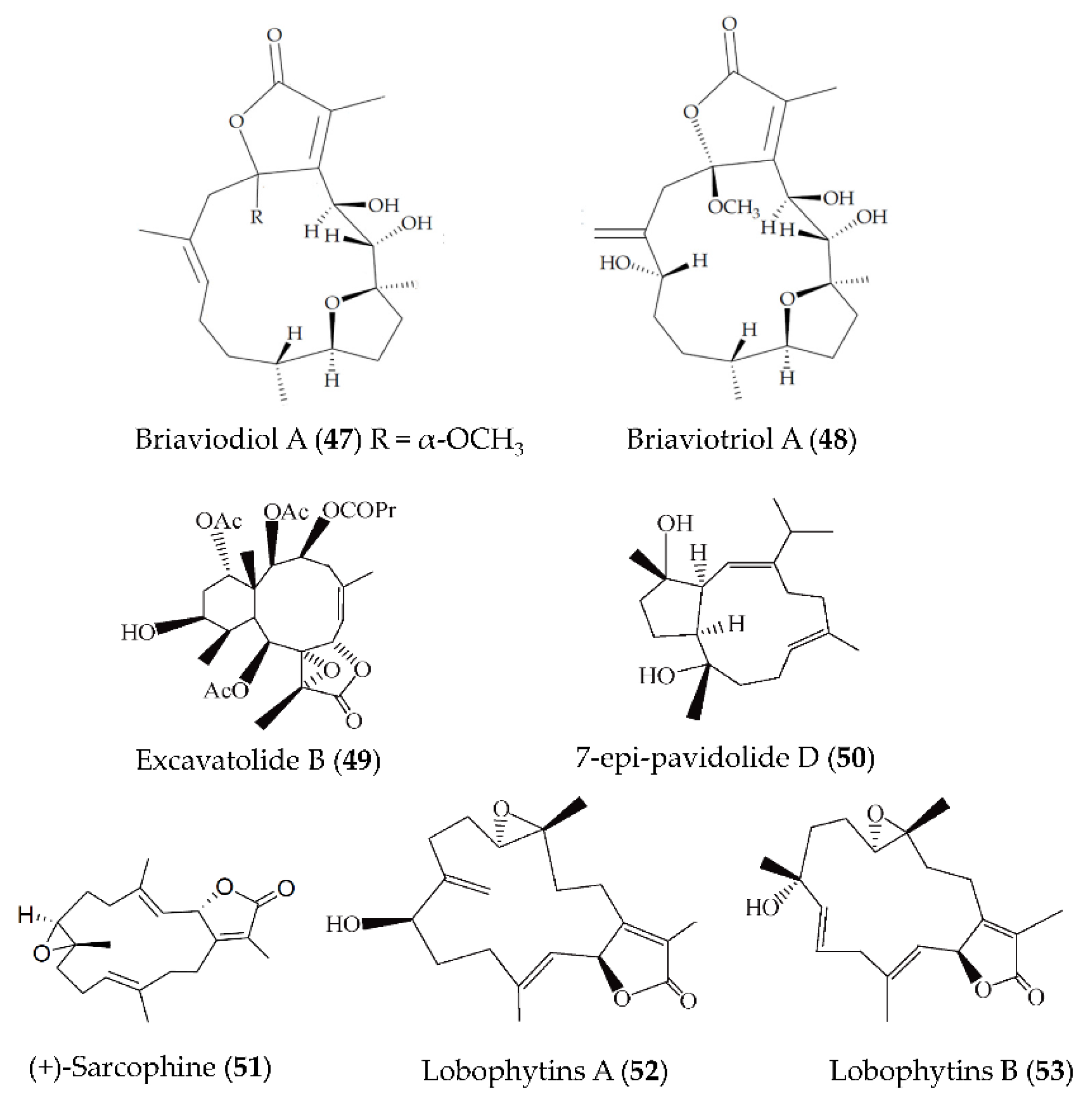
| Briaviodiol A (47) briaviotriol A (48) | Briareum violaceum | LPS-induced RAW264.7 macrophages | against iNOS release with inhibitory rate of 67.7% and 61.9% at 10 μM | [82] |
| Excavatolide B (49) | Briareum excavatum | LPS-induced RAW264.7 macrophages; carrageenan-induced mouse paw edema | against iNOS protein expression at concentrations ranging from 1 to 50 μM and against iNOS protein expression at 50 μM; against edema and redness of hind paws at 15 and 60 mg/kg | [83] |
| 7-epi-pavidolide D (50) | Klyxum flaccidum | fMLF/CB-induced human neutrophils | against 24.46% of superoxide anion generation and 29.96% of elastase release with IC50 > 10 μM | [84] |
| (+)-Sarcophine (51) | Sarcophyton stellatum | LPS-induced RAW264.7 macrophages | against iNOS protein expression at 50 and 100 µM, and COX-2 expression at 25–100 µM | [85] |
| Lobophytins A and B (52, 53) | Lobophytum sarcophytoides | LPS-induced RAW264.7 macrophages | against NO with IC50 values of 26.7 and 17.6 µM | [86] |
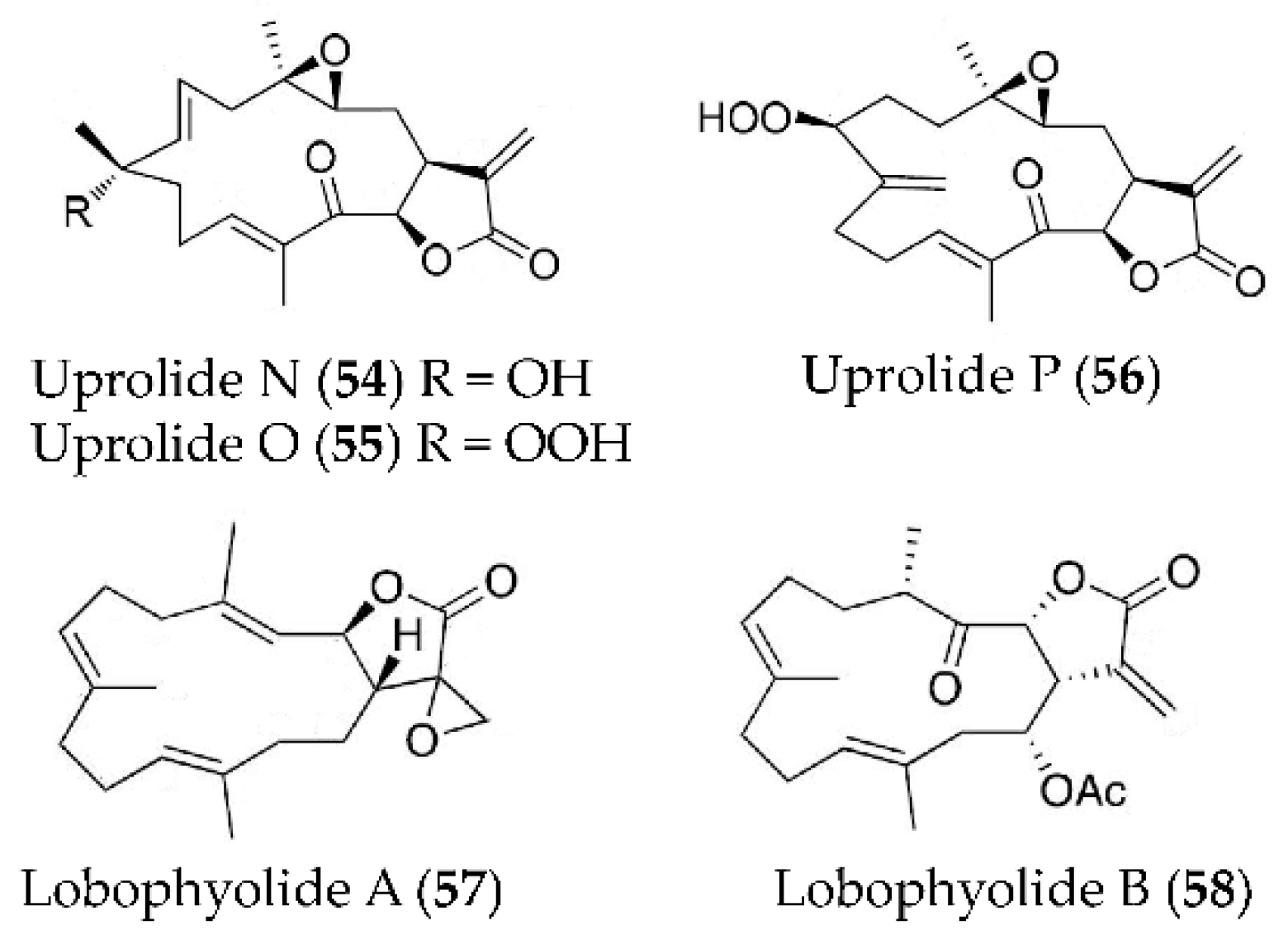
| Uprolide N, O and P (54–56) | Eunicea succinea | LPS-induced peritoneal macrophages | against TNFα production with IC50 values of 1.39, 2.73 and 2.27 µM, and against IL-6 production with IC50 values of 3.26, 4.22 and 2.60 µM | [87] |
| Lobophyolide A and B (57, 58) | Lobophytum crassum | LPS-activated DCs | against IL-12 release with inhibitory rate of 93.4% and 93.6% at 50 µg/mL; against NO production with inhibitory rate of 93.5% and 95.9% at 50 µg/mL | [88] |
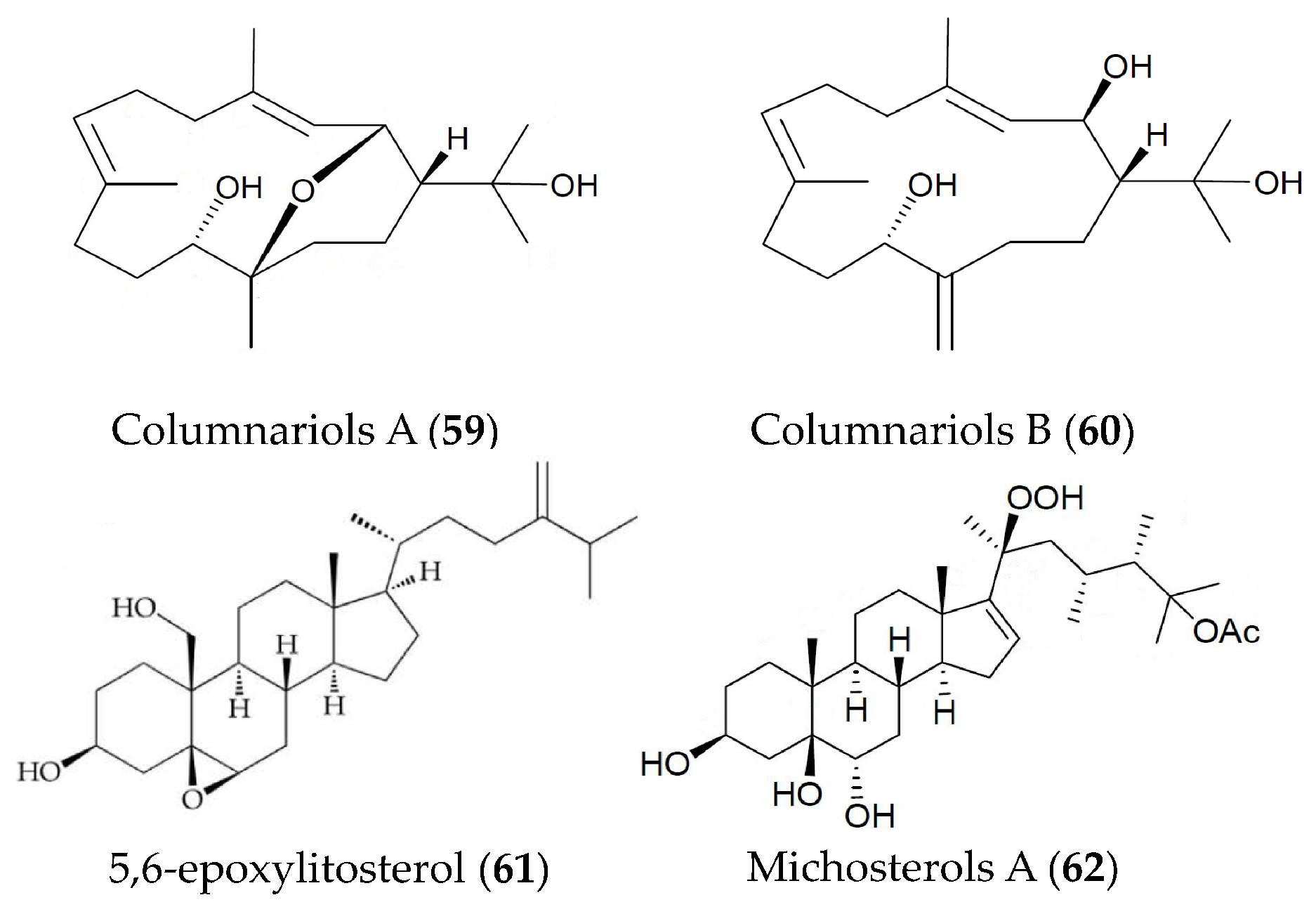
| Columnariols A and B (59, 60) | Nephthea columnaris | LPS-induced RAW264.7 macrophages | against iNOS and COX-2 protein expressions at 50 µM | [89] |
| 5,6-epoxylitosterol (61) | Nephthea columnaris | fMet-Leu-Phe/Cytochalastin B induced human neutrophils | against superoxide anions generation and elastase release with IC50 values of 4.60 and 3.90 µM | [90] |
| Michosterols A (62) | Lobophytum michaelae | fMLF/CB-induced human neutrophils | against superoxide anions generation and elastase release with IC50 values of 7.1 and 4.5 µM | [91] |
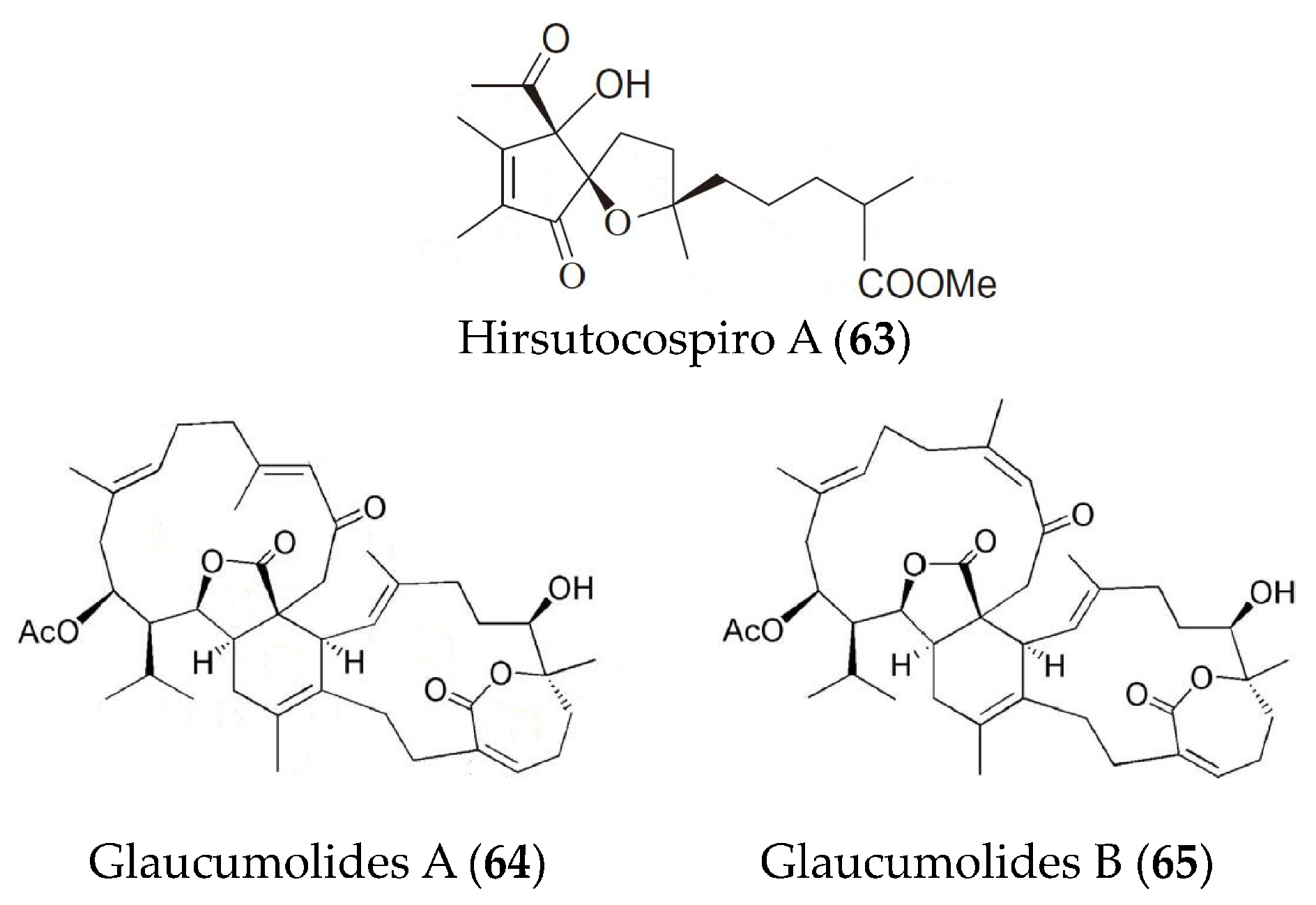
| Hirsutocospiro A (63) | Cladiella hirsuta. | fMLF/CB-induced human neutrophils | against superoxide generation and elastase release with IC50 values of 4.1 and 3.7 µM | [92] |
| Glaucumolides A and B (64, 65) | Sarcophyton glaucum | fMLP/CB-stimulated human neutrophils; LPS-induced RAW264.7 macrophages | against superoxide anion generation and elastase release with IC50 values of 2.79 and 3.97 µM; against iNOS and COX-2 expression at concentrations of 10 and 20 µM | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.-Q.; Ma, Q.-Y.; Gao, X.-Z.; Wang, X.; Zhang, B.-L. Research Progress in Anti-Inflammatory Bioactive Substances Derived from Marine Microorganisms, Sponges, Algae, and Corals. Mar. Drugs 2021, 19, 572. https://0-doi-org.brum.beds.ac.uk/10.3390/md19100572
Li C-Q, Ma Q-Y, Gao X-Z, Wang X, Zhang B-L. Research Progress in Anti-Inflammatory Bioactive Substances Derived from Marine Microorganisms, Sponges, Algae, and Corals. Marine Drugs. 2021; 19(10):572. https://0-doi-org.brum.beds.ac.uk/10.3390/md19100572
Chicago/Turabian StyleLi, Chao-Qun, Qin-Yuan Ma, Xiu-Zhen Gao, Xuan Wang, and Bei-Li Zhang. 2021. "Research Progress in Anti-Inflammatory Bioactive Substances Derived from Marine Microorganisms, Sponges, Algae, and Corals" Marine Drugs 19, no. 10: 572. https://0-doi-org.brum.beds.ac.uk/10.3390/md19100572