
In Vitro Anti-Orthohantavirus Activity of the High-and Low-Molecular-Weight Fractions of Fucoidan from the Brown Alga Fucus evanescens

 , , , and
, , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
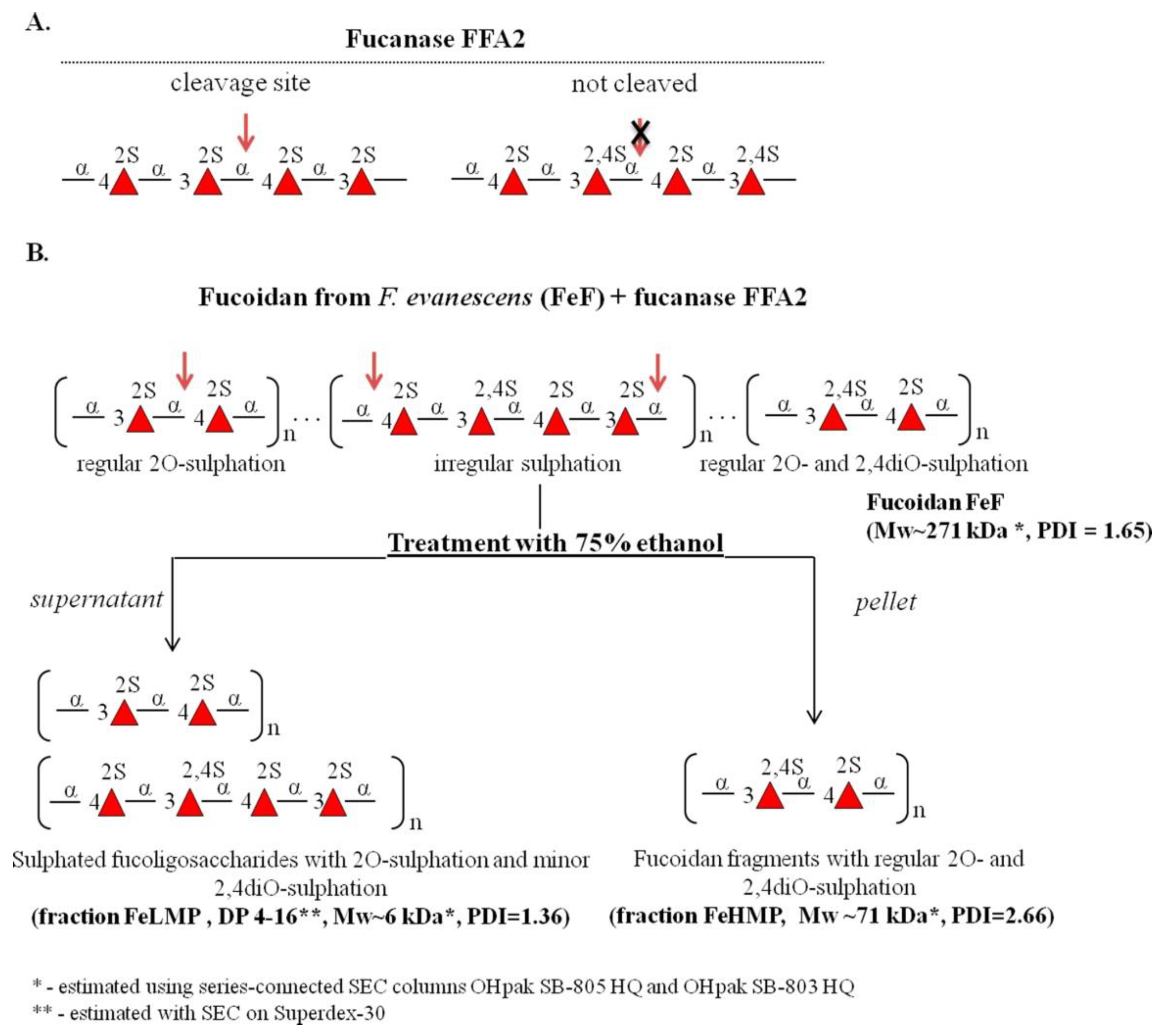
2.1. Preparation of Enzymatically Modified Fucoidan Derivatives
2.2. Antiviral Activity of the Fucoidans against the Orthohantavirus Amur
2.3. Computer Modelling
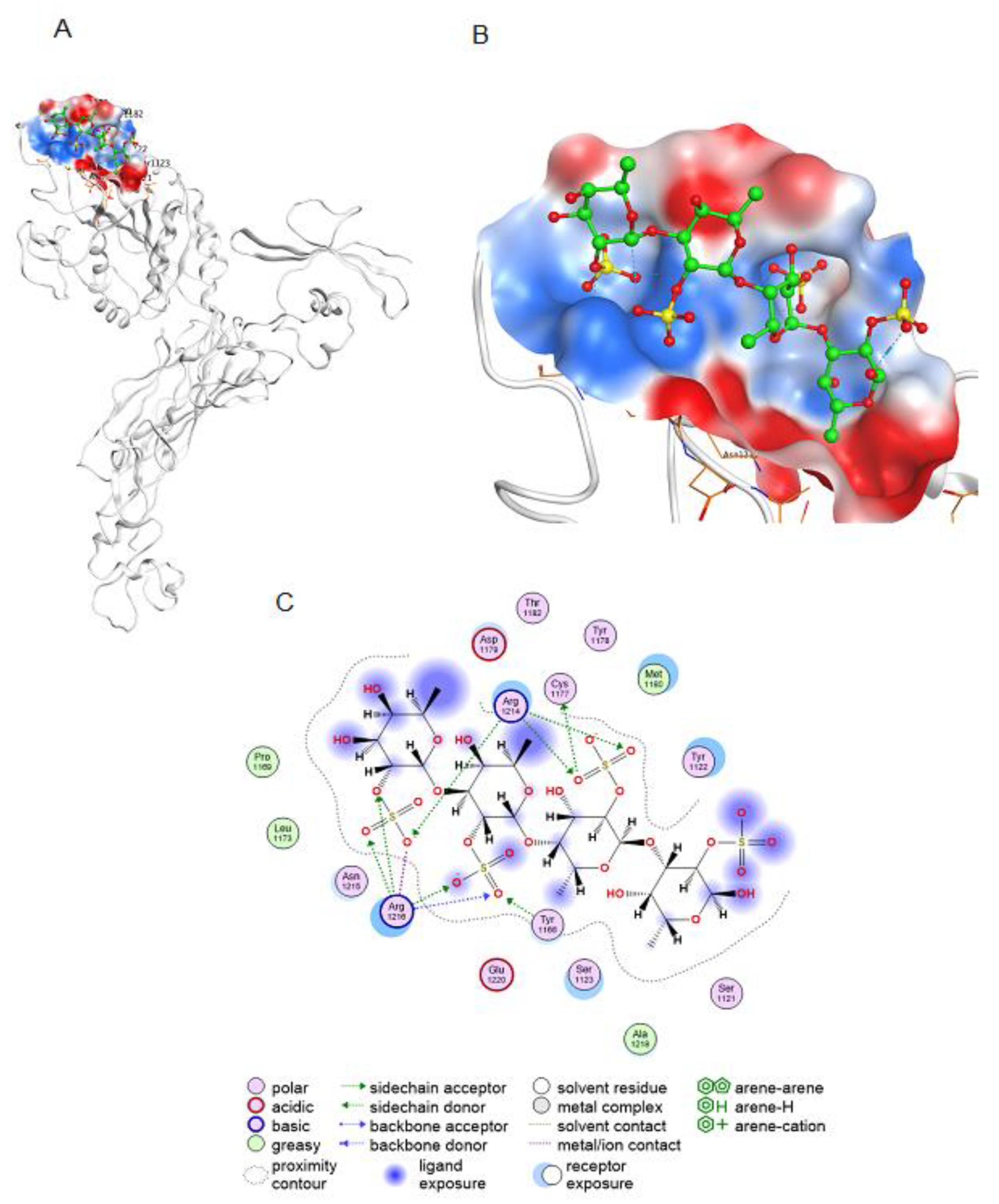
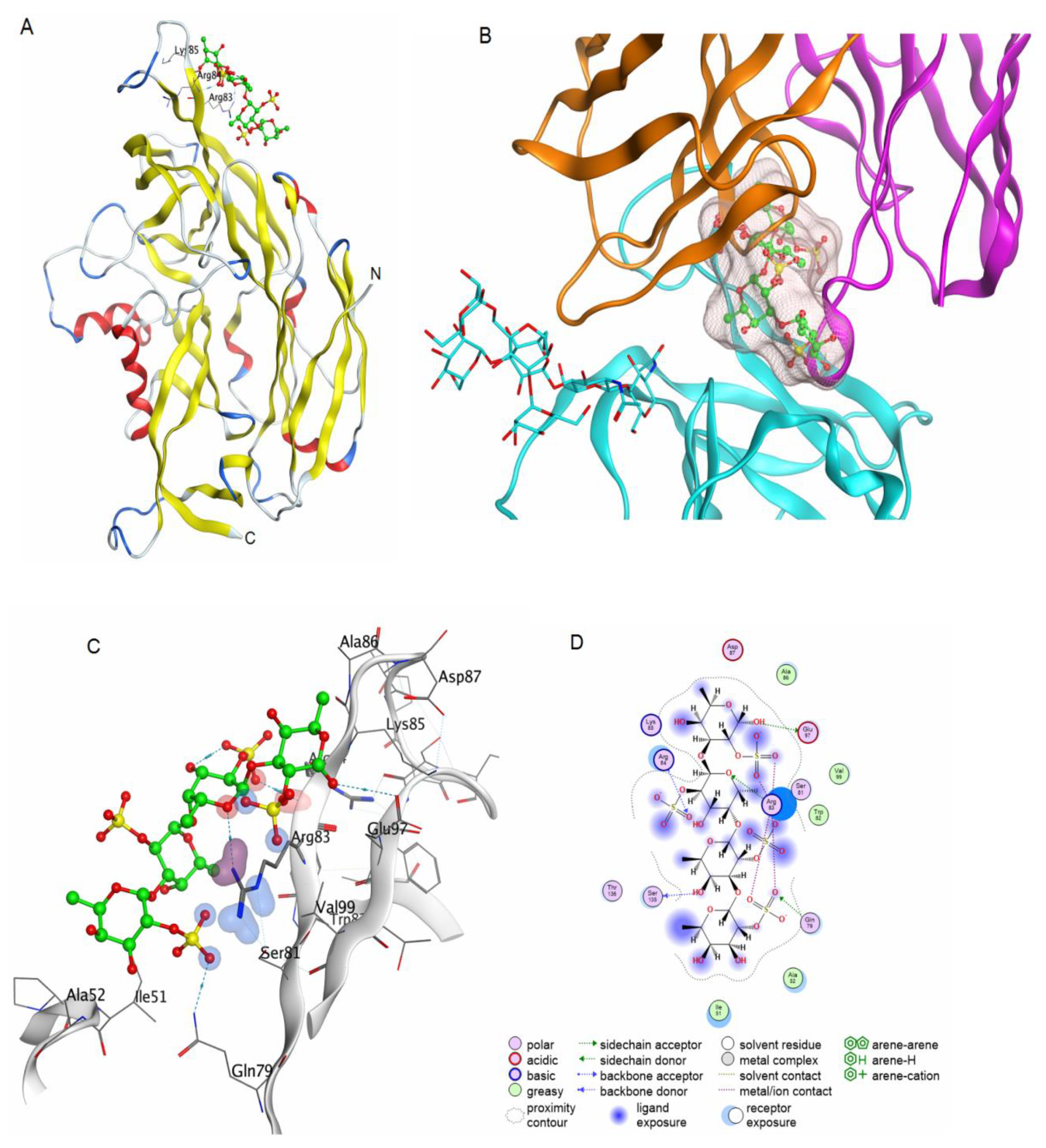
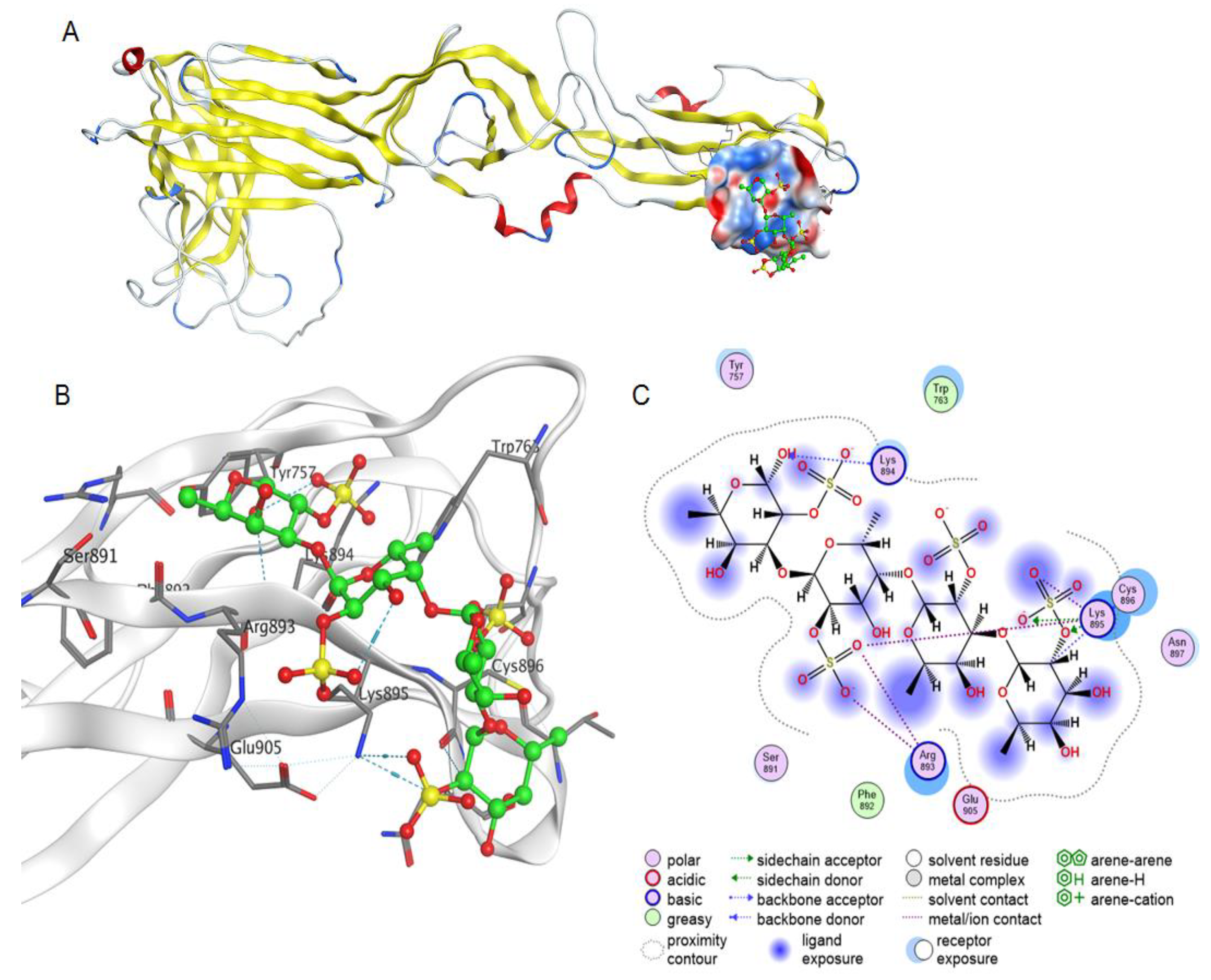
2.3.1. Molecular Docking of 2O-Sulphated Fucooligosaccharide with the Integrin β3 Epitope
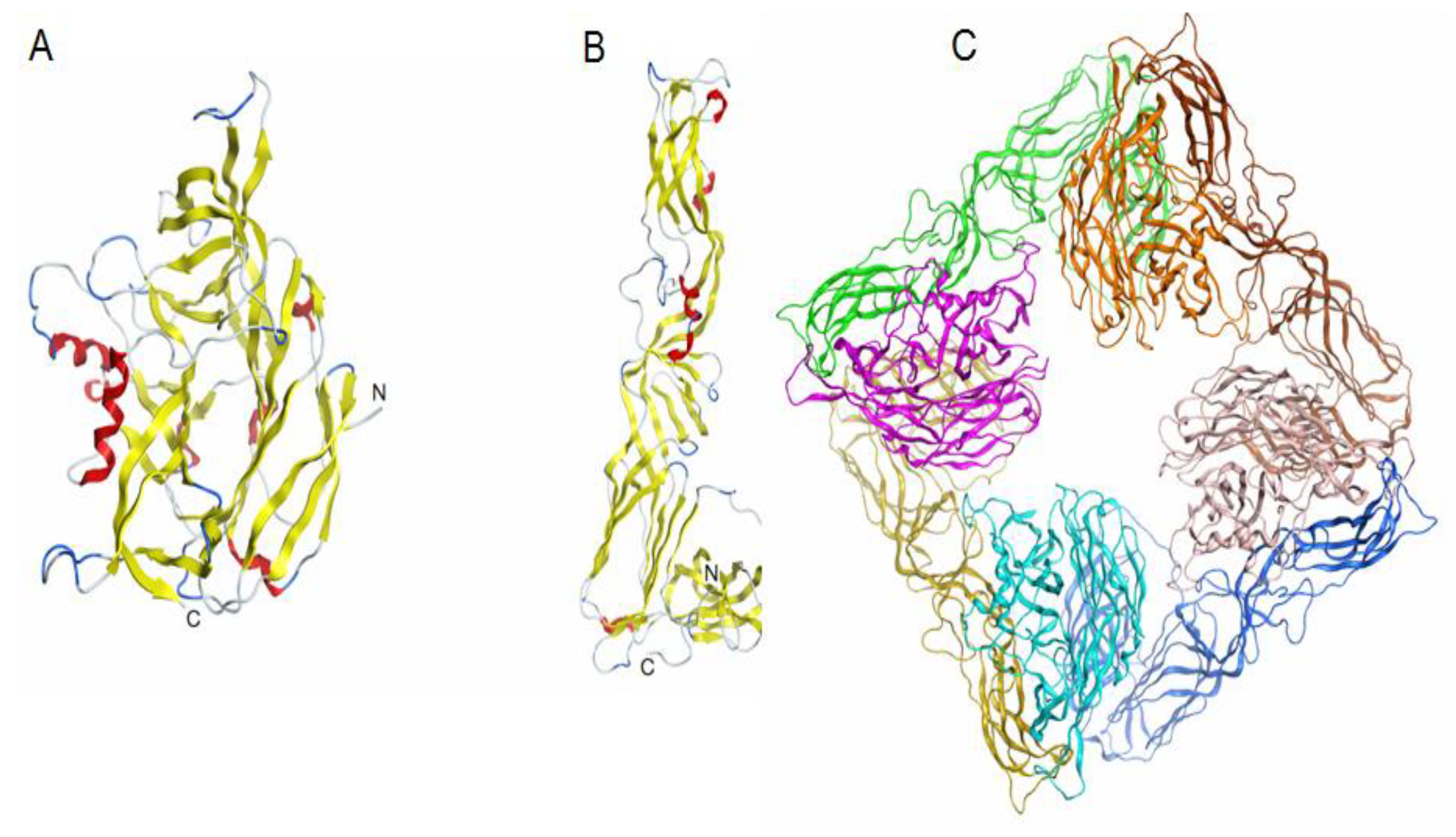
2.3.2. Modelling the 3D Structure of the AMRV Glycoprotein Ectodomains Gn and Gc and the Spike Complex Tetramer (Gn-Gc)4
2.3.3. Molecular Docking of Fucooligosaccharide into the Epitopes for Neutralising Antibodies at the AMRV Gn and Gc Ectodomains
3. Discussion
3.1. Structural Organisation of FeF
3.2. Comparative Analysis of the Anti-AMRV Activity of FeF and Its Derivatives
3.3. The Proposed Mechanism of Action of Fucoidans and Sulphated Fucooligosaccharides on AMRV
4. Materials and Methods
4.1. Virus and Cell Culture
4.2. Studied Compounds
4.3. Preparation of Enzymatic Hydrolysis Products of Fucoidan
4.4. Analytical Methods
4.4.1. Determination of the DP of Oligosaccharides
4.4.2. Molecular Weight Determination of the Fucoidan Samples
4.4.3. PAGE of Sulphated Oligosaccharides
4.4.4. NMR Spectroscopy
4.5. Virological Methods
4.5.1. Focus Formation Assay
4.5.2. Cytotoxicity of the Fucoidans
4.5.3. Anti-AMRV Activity of the Fucoidans
- -
- Pre-treatment of virus with compounds. The virus was mixed with compounds at a 1:1 (v/v) ratio, pre-incubated for 1 h at 37 °C. Then the mixture was applied to the cellular monolayer and incubated for 1 h at 37 °C. After removing the mixture, the cells were washed with PBS and incubated with DMEM containing 0.6% carboxymethyl cellulose (CMC).
- -
- Pre-treatment of cells with compounds. A monolayer of cells was pre-treated with compounds for 1 h at 37 °C before infection. The cells were then rinsed with PBS to remove the compounds and infected with the virus for 1 h at 37 °C. Thereafter, the cells were washed with PBS to remove the unbound virus and incubated with maintenance DMEM with 0.6% CMC.
- -
- The attachment assay. The monolayer of cells was pre-chilled at 4 °C for 1 h and then treated with a mixture of virus and compound (1:1). After further incubation at 4 °C for another 3 h, the compounds and unabsorbed virus were removed by washing with cold PBS and the cells were incubated with DMEM with 0.6% CMC.
- -
- The penetration assay. The monolayer of cells, pre-chilled at 4 °C for 1 h, was infected with the virus and incubated at 4 °C another for 3 h. The unbound virus was removed with cold PBS and the infected cells were treated with the compounds and incubated for 1 h at 37 °C to allow viral penetration. Then, the unpenetrated virus was inactivated with citrate buffer (pH 3.0) and the cells were washed with PBS and incubated with DMEM with 0.6% CMC.
- -
- Treatment of infected cells. The monolayer of cells was infected with the virus at 37 °C for 1 h, then washed and overlaid with DMEM with 0.6% CMC containing different concentrations of the studied compounds.
4.6. Computer Modelling
4.6.1. Modelling the 3D Structure of the 2O-Sulphated Fucooligosaccharide
4.6.2. Homology Modelling of the AMRV Glycoproteins Gn and Gc
4.6.3. Molecular Docking of the AMRV Gn and Gc Glycoproteins and Human Integrin β3 with 2O-Sulphated Fucooligosaccharide
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Milholland, M.T.; Castro-Arellano, I.; Suzan, G.; Garcia-Pena, G.E.; Lee, T.E., Jr.; Rohde, R.E.; Aguirre, A.A.; Mills, J.N. Global Diversity and Distribution of Hantaviruses and Their Hosts. EcoHealth 2018, 15, 163–208. [Google Scholar] [CrossRef]
- Kabwe, E.; Davidyuk, Y.; Shamsutdinov, A.; Garanina, E.; Martynov, E.; Kitaeva, K.; Malisheni, M.; Isaeva, G.; Savitskaya, T.; Urbanowicz, R.A.; et al. Orthohantaviruses, Emerging Zoonotic Pathogens. Pathogens 2020, 9, 775. [Google Scholar] [CrossRef] [PubMed]
- Avsic-Zupanc, T.; Saksida, A.; Korva, M. Hantavirus infections. Clin. Microbiol. Inf. 2019, 21, e6–e16. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Ma, H.; Shu, J.; Zhang, Q.; Han, M.; Liu, Z.; Jin, X.; Zhang, F.; Wu, X. Vaccines and therapeutics against Hantaviruses. Front. Microbiol. 2019, 10, 2989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, M.H.; Patel, T.R. Biodefense Implications of New-World Hantaviruses. Front. Bioeng. Biotechnol. 2020, 8, 925. [Google Scholar] [CrossRef] [PubMed]
- Tkachenko, E.A.; Ishmukhametov, A.A.; Dzagurova, T.K.; Bernshtein, A.D.; Morozov, V.G.; Siniugina, A.A.; Kurashova, S.S.; Balkina, A.S.; Tkachenko, P.E.; Kruger, D.H.; et al. Hemorrhagic Fever with Renal Syndrome, Russia. Emerg. Infect. Dis. 2019, 25, 2325–2328. [Google Scholar] [CrossRef]
- Slonova, R.A.; Kushnareva, T.V.; Iunikhina, O.V.; Maksema, I.G.; Kompanets, G.G.; Kushnarev, E.L.; Borzov, V.P. Epidemiological and epizootological characteristics of foci with a group incidence of hemorrhagic fever with renal syndrome in the Primorsky Territory. Epidemiol. Infect. Dis. 2013, 3, 10–13. [Google Scholar]
- Yashina, L.N.; Hay, J.; Smetannikova, N.A.; Kushnareva, T.V.; Iunikhina, O.V.; Kompanets, G.G. Hemorrhagic Fever with Renal Syndrome in Vladivostok City, Russia. Front. Public Health 2021, 9, 620279. [Google Scholar] [CrossRef]
- Szabo, R. Antiviral therapy and prevention against hantavirus infections. Acta Virol. 2017, 61, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Rusnak, J.M.; Byrne, W.R.; Chung, K.N.; Gibbs, P.H.; Kim, T.T.; Boudreau, E.F.; Cosgriff, T.; Pittman, P.; Kim, K.Y.; Erlichman, M.S.; et al. Experience with intravenous ribavirin in the treatment of hemorrhagic fever with renal syndrome in Korea. Antivir. Res. 2009, 81, 68–76. [Google Scholar] [CrossRef]
- Malinin, O.V.; Platonov, A.E. Insufficient efficacy and safety of intravenous ribavirin in treatment of haemorrhagic fever with renal syndrome caused by Puumala virus. Infect. Dis. 2017, 49, 514–520. [Google Scholar] [CrossRef]
- Usov, A.I.; Bilan, M.I. Fucoidans-sulfated polysaccharides of brown algae. Success Chem. 2009, 78, 846–862. [Google Scholar] [CrossRef]
- Rabanal, M.; Ponce, N.M.A.; Navarro, D.A.; Gómez, R.M.; Stortz, C.A. The system of fucoidans from the brown seaweed Dictyota dichotoma: Chemical analysis and antiviral activity. Carbohydr. Polym. 2014, 101, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Cunha, L.; Grenha, A. Sulfated Seaweed Polysaccharides as Multifunctional Materials in Drug Delivery Applications. Mar. Drugs 2016, 14, 42. [Google Scholar] [CrossRef]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological Activities of Fucoidan and the Factors Mediating Its Therapeutic Effects: A Review of Recent Studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef] [Green Version]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Lu, F.; Wei, X.J.; Zhao, R.X. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671. [Google Scholar] [CrossRef] [Green Version]
- Imbs, T.I.; Krasovskaya, N.P.; Ermakova, S.P.; Makarieva, T.N.; Shevchenko, N.M.; Zvyagintseva, T.N. Comparative study of chemical composition and antitumor activity of aqueous-ethanol extracts of brown algae Laminaria cichorioides, Costaria costata, and Fucus evanescens. Russ. J. Mar. Biol. 2009, 35, 164–170. [Google Scholar] [CrossRef]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silchenko, A.S.; Ustyuzhanina, N.E.; Kusaykin, M.I.; Krylov, V.B.; Shashkov, A.S.; Dmitrenok, A.S.; Usoltseva, R.V.; Zueva, A.O.; Nifantiev, N.E.; Zvyagintseva, T.N. Expression and biochemical characterization and substrate specificity of the fucoidanase from Formosa algae. Glycobiology 2017, 27, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Rasin, A.B.; Silchenko, A.S.; Kusaykin, M.I.; Malyarenko, O.S.; Zueva, A.O.; Kalinovsky, A.I.; Airong, J.; Surits, V.V.; Ermakova, S.P. Enzymatic transformation and anti-tumor activity of Sargassum horneri fucoidan. Carbohydr. Polym. 2020, 246, 116635. [Google Scholar] [CrossRef] [PubMed]
- Colin, S.; Deniaud, E.; Jam, M.; Descamps, V.; Chevolot, Y.; Kervarec, N.; Yvin, J.-C.; Barbeyron, T.; Michel, G.; Kloareg, B. Cloning and biochemical characterization of the fucanase FcnA: Definition of a novel glycoside hydrolase family specific for sulfated fucans. Glycobiology 2006, 16, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Chang, Y.; Zhang, Y.; Mei, X.; Xue, C. Discovery and Characterization of an Endo-1,3-Fucanase From Marine Bacterium Wenyingzhuangia fucanilytica: A Novel Glycoside Hydrolase Family. Front. Microbiol. 2020, 11, 1674. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Rasin, A.B.; Kusaykin, M.I.; Malyarenko, O.S.; Shevchenko, N.M.; Zueva, A.O.; Kalinovsky, A.I.; Zvyagintseva, T.N.; Ermakova, S.P. Modification of native fucoidan from Fucus evanescens by recombinant fucoidanase from marine bacteria Formosa algae. Carbohydr. Polym. 2018, 193, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Rasin, A.B.; Zueva, A.O.; Kusaykin, M.I.; Zvyagintseva, T.N.; Kalinovsky, A.I.; Kurilenko, V.V.; Ermakova, S.P. Fucoidan Sulfatases from Marine Bacterium Wenyingzhuangia fucanilytica CZ1127T. Biomolecules 2018, 8, 98. [Google Scholar] [CrossRef] [Green Version]
- Krylova, N.V.; Ermakova, S.P.; Lavrov, V.F.; Leneva, I.A.; Kompanets, G.G.; Iunikhina, O.V.; Nosik, M.N.; Ebralidze, L.K.; Falynskova, I.N.; Silchenko, A.S.; et al. The Comparative Analysis of Antiviral Activity of Native and Modified Fucoidans from Brown Algae Fucus evanescens In Vitro and In Vivo. Mar. Drugs 2020, 18, 224. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.E.; Kariwa, H.; Mizutani, T.; Yoshimatsu, K.; Arikawa, J.; Takashima, I. In vitro antiviral activity of lactoferrin and ribavirin upon hantavirus. Arch. Virol. 2000, 145, 1571–1582. [Google Scholar] [CrossRef]
- Safronetz, D.; Haddock, E.; Feldmann, F.; Ebihara, H.; Feldmann, H. In Vitro and In Vivo Activity of Ribavirin against Andes Virus Infection. PLoS ONE 2011, 6, e23560. [Google Scholar] [CrossRef]
- Ogg, M.; Jonsson, C.B.; Camp, J.V.; Hooper, J.W. Ribavirin Protects Syrian Hamsters against Lethal Hantavirus Pulmonary Syndrome—After Intranasal Exposure to Andes Virus. Viruses 2013, 5, 2704–2720. [Google Scholar] [CrossRef] [Green Version]
- Gavrilovskaya, I.N.; Shepley, M.; Shaw, R.; Ginsberg, M.H.; Mackow, E.R. beta3 Integrins mediate the cellular entry of hantaviruses that cause respiratory failure. Proc. Natl. Acad. Sci. USA 1998, 95, 7074–70799. [Google Scholar] [CrossRef] [Green Version]
- Larson, R.S.; Brown, D.C.; Ye, C.; Hjelle, B. Peptide antagonists that inhibit Sin Nombre virus and hantaan virus entry through the beta3-integrin receptor. J. Virol. 2005, 79, 7319–7326. [Google Scholar] [CrossRef] [Green Version]
- Hall, P.R.; Malone, L.; Sillerud, L.O.; Ye, C.; Hjelle, B.L.; Larson, R.S. Characterization and NMR solution structure of a novel cyclic pentapeptide inhibitor of pathogenic hantaviruses. Chem. Biol. Drug Des. 2007, 69, 180–190. [Google Scholar] [CrossRef]
- Li, S.; Rissanen, I.; Zeltina, A.; Hepojoki, J.; Raghwani, J.; Harlos, K.; Pybus, O.G.; Huiskonen, J.T.; Bowden, T.A. A Molecular-Level Account of the Antigenic Hantaviral Surface. Cell Rep. 2016, 15, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Rissanen, I.; Krumm, S.A.; Stass, R.; Whitaker, A.; Voss, J.E.; Bruce, E.A.; Rothenberger, S.; Kunz, S.; Burton, D.R.; Huiskonen, J.T.; et al. Structural Basis for a Neutralizing Antibody Response Elicited by a Recombinant Hantaan Virus Gn Immunogen. mBio 2021, 6, e0253120. [Google Scholar] [CrossRef]
- Yan, G.; Zhang, Y.; Ma, Y.; Yi, J.; Liu, B.; Xu, Z.; Zhang, Y.; Zhang, C.; Zhang, F.; Xu, Z.; et al. Identification of a novel B-cell epitope of Hantaan virus glycoprotein recognized by neutralizing 3D8 monoclonal antibody. J. Gen. Virol. 2012, 93 Pt 12, 2595–2600. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Hayashi, K.; Hashimoto, M.; Nakano, T.; Hayashi, T. Novel Antiviral Fucoidan from Sporophyll of Undaria pinnatifida (Mekabu). Chem. Pharm. Bull. 2004, 52, 1091–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K.; Nakano, T.; Hashimoto, M.; Kanekiyo, K.; Hayashi, T. Defensive effects of a fucoidan from brown alga Undaria pinnatifida against herpes simplex virus infection. Int. Immunopharmacol. 2008, 8, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Hidari, K.I.; Takahashi, N.; Arihara, M.; Nagaoka, M.; Morita, K.; Suzuki, T. Structure and anti-dengue virus activity of sulfated polysaccharide from a marine alga. Biochem. Biophys. Res. Commun. 2008, 376, 91–95. [Google Scholar] [CrossRef]
- Thuy, T.T.T.; Ly, B.M.; Van, T.T.T.; Quang, N.V.; Tu, H.C.; Zheng, Y.; Seguin-Devaux, C.; Bilan, M.I.; Usov, A.I. Anti-HIV activity of fucoidans from three brown seaweed specie. Carbohydr. Polym. 2015, 115, 122–128. [Google Scholar] [CrossRef]
- Wang, W.; Wu, J.; Zhang, X.; Hao, C.; Zhao, X.; Jiao, G.; Shan, X.; Tai, W.; Yu, G. Inhibition of influenza A virus infection by fucoidan targeting viral neuraminidase and cellular EGFR pathway. Sci. Rep. 2017, 7, 40760. [Google Scholar] [CrossRef]
- Maksema, I.G.; Makarenkova, I.D. Antiviral activity of naturally occurring fucoidans in experimental infection caused by Hantavirus. Pac. Med. J. 2008, 2, 86–89. [Google Scholar]
- Makarenkova, I.D.; Kompanets, G.G.; Besednova, N.N.; Slonova, R.A.; Zviagintseva, T.N.; Shevchenko, N.M. Screening of biopolymers from sea hydrocoles affecting the adsorption of Hantaan virus. Virus Res. 2007, 52, 29–32. [Google Scholar]
- Makarenkova, I.D.; Kompanets, G.G.; Besednova, N.N.; Slonova, R.A.; Zviagintseva, T.N.; Shevchenko, N.M. Inhibiting effects of fucoidans on Hantaan virus adsorption on a model of peritoneal macrophages in vitro. Virus Res. 2008, 53, 12–15. [Google Scholar]
- Pavliga, S.N.; Kompanets, G.G.; Tsygankov, V.Y. The Experimental Research (In Vitro) of Carrageenans and Fucoidans to Decrease Activity of Hantavirus. Food Environ. Virol. 2016, 8, 120–124. [Google Scholar] [CrossRef]
- Zayed, A.; Ulber, R. Fucoidan production: Approval key challenges and opportunities. Carbohydr. Polym. 2019, 211, 289–297. [Google Scholar] [CrossRef]
- Preeprame, S.; Lee, J.B.; Hayashi, K.; Sankawa, U. A novel antivirally active fucan sulfate derived from an edible brown alga, Sargassum horneri. Chem. Pharm. Bull. 2001, 49, 484–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, K.J.; Merry, C.L.R.; Lyon, M.; Thompson, J.E.; Roberts, I.S.; Gallagher, J.T. A New Model for the Domain Structure of Heparan Sulfate Based on the Novel Specificity of K5 Lyase. JBC 2004, 279, 27239–27245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kuppevelt, T.H.; Oosterhof, A.; Versteeg, E.M.M.; Podhumljak, E.; van de Westerlo, E.M.A.; Daamen, W.F. Sequencing of glycosaminoglycans with potential to interrogate sequence-specific interactions. Sci. Rep. 2017, 7, 14785. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, T.; Chattopadhyay, K.; Marschall, M.; Karmakar, P.; Mandal, P.; Ray, B. Focus on antivirally active sulfated polysaccharides: From structure–activity analysis to clinical evaluation. Glycobiology 2009, 19, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Tan, X.; Zhang, Y.; Li, F.; Ping, L.; Liu, H. Molecular Targets and Related Biologic Activities of Fucoidan: A Review. Mar. Drugs 2020, 18, 376. [Google Scholar] [CrossRef]
- Cifuentes-Muñoz, N.; Salazar-Quiroz, N.; Tischler, N.D. Hantavirus Gn and Gc Envelope Glycoproteins: Key Structural Units for Virus Cell Entry and Virus Assembly. Viruses 2014, 6, 1801–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leger, P.; Lozach, P.Y. Bunyaviruses: From transmission by arthropods to virus entry into the mammalian host first-target cells. Future Virol. 2015, 10, 859–881. [Google Scholar] [CrossRef] [Green Version]
- Mittler, E.; Dieterle, M.E.; Kleinfelter, L.M.; Slough, M.M.; Chandran, K.; Jangra, R.K. Hantavirus entry: Perspectives and recent advances. Adv. Virus Res. 2019, 104, 185–224. [Google Scholar] [CrossRef] [PubMed]
- Ermonval, M.; Baychelier, F.; Tordo, N. What Do We Know about How Hantaviruses Interact with Their Different Hosts? Viruses 2016, 8, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavrilovskaya, I.N.; Brown, E.J.; Ginsberg, M.H.; Mackow, E.R. Cellular entry of hantaviruses which cause hemorrhagic fever with renal syndrome is mediated by beta3 integrins. J. Virol. 1999, 73, 3951–3959. [Google Scholar] [CrossRef] [Green Version]
- Popugaeva, E.; Witkowski, P.T.; Schlegel, M.; Ulrich, R.G.; Auste, B.; Rang, A.; Krüger, D.H.; Klempa, B. Dobrava-Belgrade hantavirus from Germany shows receptor usage and innate immunity induction consistent with the pathogenicity of the virus in humans. PLoS ONE 2012, 7, e35587. [Google Scholar] [CrossRef] [Green Version]
- Zen, K.; Liu, Y.; Cairo, D.; Parkos, C.A. CD11b/CD18-dependent interactions of neutrophils with intestinal epithelium are mediated by fucosylated proteoglycans. J. Immunol. 2002, 169, 5270–5278. [Google Scholar] [CrossRef] [Green Version]
- Pan, T.-J.; Li, L.-X.; Zhang, J.-W.; Yang, Z.-S.; Shi, D.-M.; Yang, Y.-K.; Wu, W.-Z. Antimetastatic Effect of Fucoidan-Sargassum against Liver Cancer Cell Invadopodia Formation via Targeting Integrin αVβ3 and Mediating αVβ3/Src/E2F1 Signaling. J. Cancer 2019, 10, 4777–4792. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Gonzalez, R.; Cruz-Suarez, L.E.; Ricque-Marie, D.; Mendoza-Gamboa, E.; Rodriguez-Padilla, C.; Trejo-Avila, L.M. In vitro characterization of the antiviral activity of fucoidan from Cladosiphon okamuranus against Newcastle Disease Virus. Virol. J. 2012, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Lokugamage, K.; Kariwa, H.; Hayasaka, D.; Cui, B.Z.; Iwasaki, T.; Lokugamage, N.; Ivanov, L.I.; Volkov, V.I.; Demenev, V.A.; Slonova, R.; et al. Genetic Characterization of Hantaviruses Transmitted by the Korean Field Mouse (Apodemus peninsulae), Far East Russia. Emerg. Infect. Dis. 2002, 8, 768–776. [Google Scholar] [CrossRef]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Popivnich, I.B.; Isakov, V.V.; Scobun, A.S.; Sundukova, E.V.; Elyakova, L.A. A new procedure for the separation of water-soluble polysaccharides from brown seaweeds. Carbohydr. Res. 1999, 322, 32–39. [Google Scholar] [CrossRef]
- Shevchenko, N.M.; Anastyuk, S.D.; Gerasimenko, N.I.; Dmitrenok, P.S.; Isakov, V.V.; Zvyagintseva, T.N. Polysaccharide and lipid composition of the brown seaweed Laminaria gurjanovae. Rus. J. Bioorg. Chem. 2007, 33, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Zueva, A.O.; Silchenko, A.S.; Rasin, A.B.; Kusaykin, M.I.; Usoltseva, R.V.; Kalinovsky, A.I.; Kurilenko, V.V.; Zvyagintseva, T.N.; Thinh, P.D.; Ermakova, S.P. Expression and biochemical characterization of two recombinant fucoidanases from the marine bacterium Wenyingzhuangia fucanilytica CZ1127T. Int. J. Biol. Macromol. 2020, 164, 3025–3037. [Google Scholar] [CrossRef] [PubMed]
- Dodgson, K.S. Determination of inorganic sulphate in studies on the enzymic and non-enzymic hydrolysis of carbohydrate and other sulphate esters. Biochem. J. 1961, 78, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Malkin, G.A.; Dzagurova, T.K.; Korotina, N.A.; Balovneva, M.V.; Konyushko, O.I.; Sockova, S.E.; Tkachenko, E.A. Replication features of hantaviruses—Causative agents Hemorrhagic Fever With Renal Syndrome in cell cultures of different origins. Epidemiol. Infect. Dis. 2014, 19, 18–24. [Google Scholar]
- Solà-Riera, C.; Gupta, S.; Ljunggren, H.-G.; Klingström, J. Orthohantaviruses belonging to three phylogroups all inhibit apoptosis in infected target cells. Sci. Rep. 2019, 9, 834. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Serris, A.; Stass, R.; Bignon, E.A.; Muena, N.A.; Manuguerra, J.C.; Jangra, R.K.; Li, S.; Chandran, K.; Tischler, N.D.; Huiskonen, J.T.; et al. The Hantavirus Surface Glycoprotein Lattice and Its Fusion Control Mechanism. Cell 2020, 183, 442–456. [Google Scholar] [CrossRef]
- Xiong, J.P.; Stehle, T.; Goodman, S.L.; Arnaout, M.A. A novel adaptation of the integrin PSI domain revealed from its crystal structure. J. Biol. Chem. 2004, 279, 40252–40254. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krylova, N.V.; Silchenko, A.S.; Pott, A.B.; Ermakova, S.P.; Iunikhina, O.V.; Rasin, A.B.; Kompanets, G.G.; Likhatskaya, G.N.; Shchelkanov, M.Y. In Vitro Anti-Orthohantavirus Activity of the High-and Low-Molecular-Weight Fractions of Fucoidan from the Brown Alga Fucus evanescens. Mar. Drugs 2021, 19, 577. https://0-doi-org.brum.beds.ac.uk/10.3390/md19100577
Krylova NV, Silchenko AS, Pott AB, Ermakova SP, Iunikhina OV, Rasin AB, Kompanets GG, Likhatskaya GN, Shchelkanov MY. In Vitro Anti-Orthohantavirus Activity of the High-and Low-Molecular-Weight Fractions of Fucoidan from the Brown Alga Fucus evanescens. Marine Drugs. 2021; 19(10):577. https://0-doi-org.brum.beds.ac.uk/10.3390/md19100577
Chicago/Turabian StyleKrylova, Natalia V., Artem S. Silchenko, Anastasia B. Pott, Svetlana P. Ermakova, Olga V. Iunikhina, Anton B. Rasin, Galina G. Kompanets, Galina N. Likhatskaya, and Mikhail Y. Shchelkanov. 2021. "In Vitro Anti-Orthohantavirus Activity of the High-and Low-Molecular-Weight Fractions of Fucoidan from the Brown Alga Fucus evanescens" Marine Drugs 19, no. 10: 577. https://0-doi-org.brum.beds.ac.uk/10.3390/md19100577