
Bio-Guided Isolation of Antimalarial Metabolites from the Coculture of Two Red Sea Sponge-Derived Actinokineospora and Rhodococcus spp.

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results and Discussions
2.1. Identification of Red Sea Sponge-Associated Actinobacteria
2.2. Metabolomics Analysis of the Coculture Extract of Actinokineospora spheciospongiae Strain EG49 and Rhodococcus sp. UR59 Using LC–HRMS
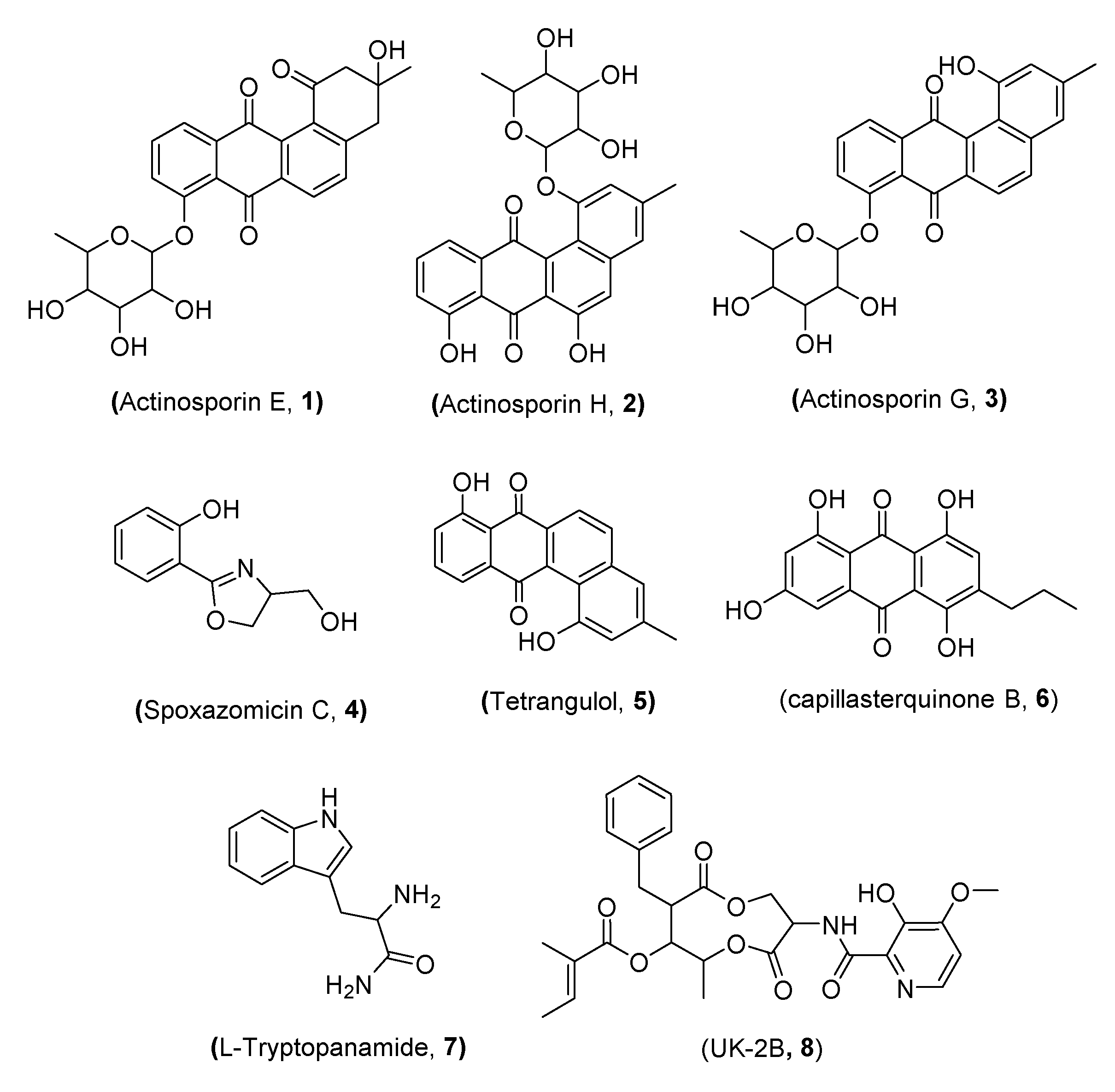
2.3. Identification of the Isolated Compounds (1–8)
2.4. Antimalarial Screening
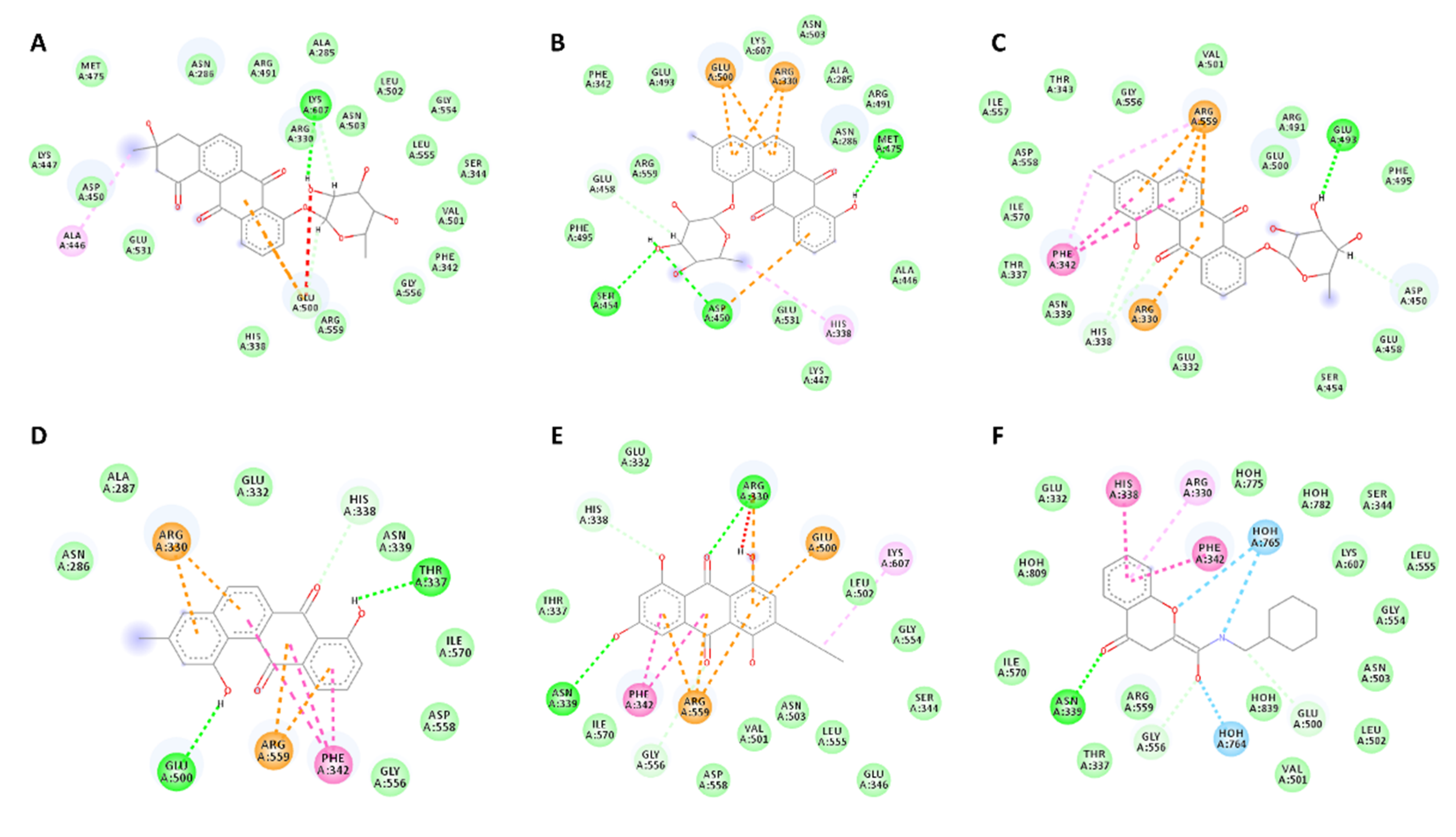
2.5. Docking Analysis
3. Materials and Methods
3.1. General Experimental
3.2. Actinomycetes Isolation
3.3. Molecular Identification and Phylogenetic Analysis
3.4. Co-Cultivation and Extract Preparation
3.5. Metabolic Profiling
3.6. Metabolites Isolation
3.7. Antimalarial Screening
3.8. Molecular Docking
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nai, C.; Meyer, V. From axenic to mixed cultures: Technological advances accelerating a paradigm shift in microbiology. Trends Microbiol. 2018, 26, 538–554. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Choi, Y.H.; van Wezel, G.P. Metabolic profiling as a tool for prioritising antimicrobial compounds. J. Ind. Microbiol. 2006, 43, 299–312. [Google Scholar]
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How many antibiotics are produced by the genus Streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.F.; Somoza, A.D.; Keller, N.P.; Wang, C.C. Advances in Aspergillus secondary metabolite research in the post-genomic era. Nat. Prod. Rep. 2012, 29, 351–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inglis, D.O.; Binkley, J.; Skrzypek, M.S.; Arnaud, M.B.; Cerqueira, G.C.; Shah, P.; Wymore, F.; Wortman, J.R.; Sherlock, G. Comprehensive annotation of secondary metabolite biosynthetic genes and gene clusters of Aspergillus nidulans, A. fumigatus, A. niger and A. oryzae. BMC Microbiol. 2013, 13, 91. [Google Scholar] [CrossRef] [Green Version]
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef]
- Bednarski, Z.; Bednarska, H. First research work by Robert Koch on etiology of anthrax-in cooperation with Józef Knechtel, Polish apothecary. Arch. Hist. Filoz. Med. 2003, 66, 161–168. [Google Scholar]
- Fischer, A. Acid production graphically registered as an indicator of the vital processes in the cultivation of bacteria. J. Exp. Med. 1918, 28, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Degenkolb, T.; Heinze, S.; Schlegel, B.; Strobel, G.; Gräfe, U. Formation of new lipoaminopeptides, acremostatins A, B, and C, by co-cultivation of Acremonium sp. Tbp-5 and Mycogone rosea DSM 12973. Biosci. Biotechnol. Biochem. 2002, 6, 883–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakefield, J.; Hassan, H.M.; Jaspars, M.; Ebel, R.; Rateb, M.E. Dual induction of new microbial secondary metabolites by fungal bacterial co-cultivation. Front. Microbiol. 2017, 8, 1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Sarotti, A.M.; Yang, B.; Turkson, J.; Cao, S. A new N-methoxypyridone from the co-cultivation of Hawaiian endophytic fungi Camporesia sambuci FT1061 and Epicoccum sorghinum FT1062. Molecules 2017, 22, 1166. [Google Scholar] [CrossRef] [Green Version]
- Thissera, B.; Alhadrami, H.A.; Hassan, M.H.; Hassan, H.M.; Bawazeer, M.; Yaseen, M.; Belbahri, L.; Rateb, M.E.; Behery, F.A. Induction of cryptic antifungal pulicatin derivatives from Pantoea agglomerans by microbial coculture. Biomolecules 2020, 10, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobson, M.J. Exploring natural remedies from the past. Parassitologia 1998, 40, 69–81. [Google Scholar]
- Pyae Phyo, A.; Nkhoma, S.; Singhasivanon, P.; Day, N.P.; White, N.J. Emergence of artemisinin-resistant malaria on the western border of Thailand: A longitudinal study. Lancet 2012, 379, 1960–1966. [Google Scholar] [CrossRef] [Green Version]
- Straimer, J.; Gnädig, N.F.; Witkowski, B.; Amaratunga, C.; Duru, V.; Ramadani, A.P.; Dacheux, M.; Khim, N.; Zhang, L.; Lam, S.; et al. K13-propeller mutations confer artemisinin resistance in Plasmodium falciparum clinical isolates. Science 2015, 347, 428–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeka, A.; Lameyre, V.; Afizi, K.; Fredrick, M.; Lukwago, R.; Kamya, M.R.; Talisuna, A.O. Efficacy and safety of fixed-dose artesunate-amodiaquine vs. artemether-lumefantrine for repeated treatment of uncomplicated malaria in Ugandan children. PLoS ONE 2014, 9, e113311. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Pimentel-Elardo, S.M.; Hanora, A.; Radwan, M.; Abou-El-Ela, S.H.; Ahmed, S.; Hentschel, U. Isolation, phylogenetic analysis and anti-infective activity screening of marine sponge-associated actinomycetes. Mar. Drugs 2010, 8, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Kampfer, P.; Glaeser, S.P.; Busse, H.J.; Abdelmohsen, U.R.; Ahmed, S.; Hentschel, U. Actinokineospora spheciospongiae sp. nov., isolated from the marine sponge Spheciospongia vagabunda. Int. J. Syst. Evol. Microbiol. 2015, 65 Pt 3, 879–884. [Google Scholar] [CrossRef]
- Bass, P.D.; Gubler, D.A.; Judd, T.C.; Williams, R.M. Mitomycinoid alkaloids: Mechanism of action, biosynthesis, total syntheses, and synthetic approaches. Chem. Rev. 2013, 113, 6816–6863. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.S.; Yang, D. Enantioselective Synthesis of (+)-Mitomycin K by a Palladium-Catalysed Oxidative Tandem Cyclization. Angew. Chem. 2017, 129, 5980–5983. [Google Scholar] [CrossRef]
- Akiyama, T.; Harada, S.; Kojima, F.; Takahashi, Y.; Imada, C.; Okami, Y.; Muraoka, Y.; Aoyagi, T.; Takeuchi, T. Fluostatins A and B, New Inhibitors of Dipeptidyl Peptidase III, Produced by Streptomyces sp. TA-3391. J. Antibiot. 1998, 51, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Abdelmohsen, U.R.; Cheng, C.; Viegelmann, C.; Zhang, T.; Grkovic, T.; Ahmed, S.; Quinn, R.J.; Hentschel, U.; Edrada-Ebel, R. Dereplication strategies for targeted isolation of new antitrypanosomal actinosporins A and B from a marine sponge associated-Actinokineospora sp. EG49. Mar. Drugs 2014, 12, 1220–1244. [Google Scholar] [CrossRef] [Green Version]
- Grkovic, T.; Abdelmohsen, U.R.; Othman, E.M.; Stopper, H.; Edrada-Ebel, R.; Hentschel, U.; Quinn, R.J. Two new antioxidant actinosporin analogues from the calcium alginate beads culture of sponge-associated Actinokineospora sp. strain EG49. Bioorg. Med. Chem. Lett. 2014, 24, 5089–5092. [Google Scholar] [CrossRef] [PubMed]
- Blumauerova, M.; Kralovcova, E.; Matějů, J.; Jizba, J.; Vaněk, Z. Biotransformations of anthracyclinones in Streptomyces coeruleorubidus and Streptomyces galilaeus. Folia Microbiol. 1979, 24, 117. [Google Scholar] [CrossRef]
- Fujioka, K.; Furihata, K.; Shimazu, A.; Hayakawa, Y.; Seto, H. Isolation and characterisation of atramycin A and atramycin B, new isotetracenone type antitumor antibiotics. J. Antibiot. 1991, 44, 1025–1028. [Google Scholar] [CrossRef]
- Mullowney, M.W.; Ó hAinmhire, E.; Tanouye, U.; Burdette, J.E.; Pham, V.C.; Murphy, B.T. A Pimarane Diterpene and Cytotoxic Angucyclines from a Marine-Derived Micromonospora sp. in Vietnam’s East Sea. Mar. Drugs 2015, 13, 5815–5827. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Ding, Z.G.; Long, Y.F.; Zhao, J.Y.; Li, M.G.; Cui, X.L.; Wen, M.L. A new isoflavone derivative from Streptomyces sp. YIM GS3536. Chem. Nat. Compd. 2013, 48, 966–969. [Google Scholar] [CrossRef]
- Maskey, R.P.; Asolkar, R.N.; Speitling, M.; Hoffman, V.; Grün-Wollny, I.; Fleck, W.F.; Laatsch, H. Flavones and new isoflavone derivatives from microorganisms: Isolation and structure elucidation. Z. Naturforsch. B 2003, 58, 686–691. [Google Scholar] [CrossRef]
- Sugumaran, M.; Robinson, W.E. Bioactive dehydrotyrosyl and dehydrodopyl compounds of marine origin. Mar. Drugs 2010, 8, 2906–2935. [Google Scholar] [CrossRef] [Green Version]
- Umezawa, H.; Imoto, M.; Sawa, T.; Isshiki, K.; Matsuda, N.; Uchida, T.; Iinuma, H.; Hamada, M.; Takeuchi, T. Studies on a new epidermal growth factor-receptor kinase inhibitor, erbstatin, produced by MH435-hF3. J. Antibiot. 1986, 39, 170–173. [Google Scholar] [CrossRef] [Green Version]
- Weber, W.; Zähner, H.; Damberg, M.; Russ, P.; Zeeck, A. Metabolic products of microorganisms 201. Ansatrienin A and B, antifungal antibiotics from Streptomyces collinus. Zentralblatt für Bakteriologie Mikrobiologie und Hygiene: I. Abt. Orig. C Allg. Angew. Okol. Mikrobiol. 1981, 2, 122–139. [Google Scholar]
- Chiba, H.; Agematu, H.; Kaneto, R.; Terasawa, T.; Sakai, K.; Dobashi, K.; Yoshioka, T. Rhodopeptins (Mer-N1033), Novel Cyclic Tetrapeptides with Antifungal Activity from Rhodococcus sp. J. Antibiot. 1999, 52, 695–699. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K.; Kawato, H.C.; Inagaki, H.; Nakajima, R.; Kitamura, A.; Someya, K.; Ohta, T. Synthesis and antifungal activity of rhodopeptin analogues. 2. Modification of the west amino acid moiety. Org. Lett. 2000, 2, 977–980. [Google Scholar] [CrossRef]
- Nam, S.J.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Isolation and characterization of actinoramides A–C, highly modified peptides from a marine Streptomyces sp. Tetrahedron 2011, 67, 6707–6712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terekhova, L.P.; Galatenko, O.A.; Kulyaeva, V.V.; Malkina, N.D.; Boikova, Y.V.; Katrukha, G.S.; Shashkov, A.S.; Gerbst, A.G.; Nifantiev, N.E. Isolation, NMR spectroscopy, and conformational analysis of the antibiotic ina 2770 (cineromycin B) produced by Streptomyces strain. Russ. Chem. 2007, 56, 815–818. [Google Scholar] [CrossRef]
- Carrano, C.J.; Jordan, M.; Drechsel, H.; Schmid, D.G.; Winkelmann, G. Heterobactins: A new class of siderophores from Rhodococcus erythropolis IGTS8 containing both hydroxamate and catecholate donor groups. Biometals 2001, 14, 119–125. [Google Scholar] [CrossRef]
- Han, X.; Liu, Z.; Zhang, Z.; Zhang, X.; Zhu, T.; Gu, Q.; Li, W.; Che, Q.; Li, D. Geranylpyrrol A and Piericidin F from Streptomyces sp. CHQ-64 Δ rdmF. J. Nat. Prod. 2017, 80, 1684–1687. [Google Scholar] [CrossRef]
- Nakae, K.; Yoshimoto, Y.; Ueda, M.; Sawa, T.; Takahashi, Y.; Naganawa, H.; Takeuchi, T.; Imoto, M. Migrastatin, a novel 14-membered lactone from Streptomyces sp. MK929-43F1. J. Antibiot. 2000, 53, 1228–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.Y.; Wang, X.Q.; Wang, Y.M.; Geng, X.; Xu, X.N.; Su, C.; Yang, Y.L.; Tang, Y.J.; Bai, F.W.; Zhao, X.Q. Genome mining of Streptomyces xinghaiensis NRRL B-24674 T for the discovery of the gene cluster involved in anticomplement activities and detection of novel xiamycin analogs. Appl. Microbiol. Biotechnol. 2018, 102, 9549–9562. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Matsubara, H.; Nakagawa, A.; Furusaki, A.; Matsumoto, T. X-Ray crystallography of protylonolide and absolute configuration of tylosin. J. Antibiot. 1980, 33, 915–917. [Google Scholar] [CrossRef] [Green Version]
- Gräfe, U.; Schlegel, R.; Stengel, C.; Ihn, W.; Radics, L. Isolation and structure of 26-deoxylaidlomycin, a new polyether antibiotic from Streptoverticillium olivoreticuli. J. Basic Microbiol. 1989, 29, 149–155. [Google Scholar] [CrossRef]
- Hirota, A.; Okada, H.; Kanza, T.; Isogai, A.; Hirota, H. Structure elucidation of kaimonolide B, a new plant growth inhibitor macrolide from Streptomyces. Agric. Biol. Chem. 1990, 54, 2489–2490. [Google Scholar] [CrossRef]
- Arakawa, K. Genetic and biochemical analysis of the antibiotic biosynthetic gene clusters on the Streptomyces linear plasmid. Biosci. Biotechnol. Biochem. 2014, 78, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Kusakabe, Y.; Mizuno, T.; Kawabata, S.; Tanji, S.; Seino, A.; Seto, H.; Otake, N. Ferensimycins A and B, two polyether antibiotics. J. Antibiot. 1982, 35, 1119–1129. [Google Scholar] [CrossRef]
- Cho, J.Y.; Williams, P.G.; Kwon, H.C.; Jensen, P.R.; Fenical, W. Lucentamycins A–D, cytotoxic peptides from the marine-derived actinomycete Nocardiopsis lucentensis. J. Nat. Prod. 2007, 70, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Qian-cutrone, J.; Ueki, T.; Huang, S.; MookhtiaR, K.A.; Ezekiel, R.; Kalinowski, S.S.; Brown, K.S.; Golik, J.; Lowe, S.; Pirnik, D.M.; et al. Glucolipsin A and B, two new glucokinase activators produced by Streptomyces purpurogeniscleroticus and Nocardia vaccinii. J. Antibiot. 1999, 52, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Howlett, D.R.; George, A.R.; Owen, D.E.; Ward, R.V.; Markwell, R.E. Common structural features determine the effectiveness of carvedilol, daunomycin and rolitetracycline as inhibitors of Alzheimer β-amyloid fibril formation. Biochem. J. 1999, 343, 419–423. [Google Scholar] [CrossRef]
- Dashti, Y.; Grkovic, T.; Abdelmohsen, U.R.; Hentschel, U.; Quinn, R.J. Actinomycete metabolome induction/suppression with N-Acetylglucosamine. J. Nat. Prod. 2017, 80, 828–836. [Google Scholar] [CrossRef]
- Inahashi, Y.; Iwatsuki, M.; Ishiyama, A.; Namatame, M.; Nishihara-Tsukashima, A.; Matsumoto, A.; Hirose, T.; Sunazuka, T.; Yamada, H.; Otoguro, K.; et al. Spoxazomicins A–C, novel antitrypanosomal alkaloids produced by an endophytic actinomycete, Streptosporangium oxazolinicum K07-0460 T. J. Antibiot. 2011, 64, 303–307. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.C.C.; Cao, S.; Raveh, A.; MacArthur, R.; Dranchak, P.; Chlipala, G.; Okoneski, M.T.; Guha, R.; Eastman, R.T.; Yuan, J.; et al. Actinoramide A identified as a potent antimalarial from titration-based screening of marine natural product extracts. J. Nat. Prod. 2015, 78, 2411–2422. [Google Scholar] [CrossRef] [Green Version]
- Elleuch, L.; Shaaban, M.; Smaoui, S.; Mellouli, L.; Karray-Rebai, I.; Fguira, L.F.B.; Shaaban, K.A.; Laatsch, H. Bioactive secondary metabolites from a new terrestrial Streptomyces sp. TN262. Appl. Biochem. Biotechnol. 2010, 162, 579–593. [Google Scholar] [CrossRef] [Green Version]
- Giralt, E.; Lo Re, D. The therapeutic potential of migrastatin-core analogs for the treatment of metastatic cancer. Molecules 2017, 22, 198. [Google Scholar] [CrossRef] [Green Version]
- Ujikawa, K.; Vilegas, W.; Vilegas, J.H.; Llabrés, G. Antibiotic 26-deoxylaidlomycin isolated from Streptomyces sp. Ar386 from Brazilian soil. Rev. Latinoam. Microbiol. 1996, 38, 185–191. [Google Scholar] [PubMed]
- Le, T.V.; Hanh, T.T.H.; Huong, P.T.T.; Dang, N.H.; Van Thanh, N.; Cuong, N.X.; Nam, N.H.; Thung, D.C.; Van Kiem, P.; Van Minh, C. Anthraquinone and butenolide constituents from the crinoid Capillaster multiradiatus. Chem. Pharm. Bull. 2018, 66, 1023–1026. [Google Scholar]
- Kuntsmann, M.P.; Mitscher, L.A. The Structural Characterization of Tetrangomycin and Tetrangulol. J. Org. Chem. 1966, 31, 2920–2925. [Google Scholar] [CrossRef] [PubMed]
- Ukei, M.; Abe, K.; Hanafi, M.; Shibata, K.; Tanaka, T.; Tanaiguchi, M. UK-2 A, B, C and D Novel antifungal antibiotics from Streptomyces sp. 517-02. J. Antibiot. 1996, 49, 639–643. [Google Scholar] [CrossRef]
- Guo, Z.K.; Wang, T.; Guo, Y.; Song, Y.C.; Tan, R.X.; Ge, H.M. Cytotoxic angucyclines from Amycolatopsis sp. HCa1, a rare actinobacteria derived from Oxya chinensis. Planta Med. 2011, 77, 2057. [Google Scholar] [CrossRef] [Green Version]
- Hanafi, M.; Shibata, K.; Ueki, M.; Taniguchi, M. UK-2A, B, C and D, Novel Antifungal Antibiotics from Streptomyces sp. 517-02. J. Antibiot. 1996, 49, 1226–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konopka, J.B. N-acetylglucosamine functions in cell signalling. Scientifica 2012, 2012, 489208. [Google Scholar] [CrossRef] [Green Version]
- Naseem, S.; Parrino, S.M.; Buenten, D.M.; Konopka, J.B. Novel roles for GlcNAc in cell signalling. Commun. Integr. Biol. 2012, 5, 156–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigali, S.; Nothaft, H.; Noens, E.E.; Schlicht, M.; Colson, S.; Müller, M.; Joris, B.; Koerten, H.K.; Hopwood, D.A.; Titgemeyer, F.; et al. The sugar phosphotransferase system of Streptomyces coelicolor is regulated by the GntR-family regulator DasR and links N-acetylglucosamine metabolism to the control of development. Mol. Microbiol. 2006, 61, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Boonlarppradab, C.; Suriyachadkun, C.; Rachtawee, P.; Choowong, W. Saccharosporones A, B and C, cytotoxic antimalarial angucyclinones from Saccharopolyspora sp. BCC 21906. J. Antibiot. 2013, 266, 305–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, G.; Derbyshire, E.R.; Caldera, E.; Currie, C.R.; Clardy, J. Antibiotic and antimalarial quinones from fungus-growing ant-associated Pseudonocardia sp. J. Nat. Prod. 2012, 75, 1806–1809. [Google Scholar] [CrossRef] [PubMed]
- Supong, K.; Thawai, C.; Suwanborirux, K.; Choowong, W.; Supothina, S.; Pittayakhajonwut, P. Antimalarial and antitubercular C-glycosylated benz [α] anthraquinones from the marine-derived Streptomyces sp. BCC45596. Phytochem. Lett. 2012, 5, 651–656. [Google Scholar] [CrossRef]
- Baragaña, B.; Forte, B.; Choi, R.; Hewitt, S.N.; Bueren-Calabuig, J.A.; Pisco, J.P.; Norcross, N.R. Lysyl-tRNA synthetase as a drug target in malaria and cryptosporidiosis. Proc. Natl. Acad. Sci. USA 2019, 116, 7015–7020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashelford, K.E.; Chuzhanova, N.A.; Fry, J.C.; Jones, A.J.; Weightman, A.J. At least 1 in 20 16S rRNA sequence records currently held in public repositories is estimated to contain substantial anomalies. Appl. Environ. Microbiol. 2005, 71, 7724–7736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Peplies, J.; Glockner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life v2: Online annotation and display of phylogenetic trees made easy. Nucleic Acids Res. 2011, 39, W475–W478. [Google Scholar] [CrossRef]
- Ngwa, C.J.; Kiesow, M.J.; Papst, O.; Orchard, L.M.; Filarsky, M.; Rosinski, A.N.; Voss, T.S.; Llinás, M.; Pradel, G. Transcriptional profiling defines histone acetylation as a regulator of gene expression during human-to-mosquito transmission of the malaria parasite Plasmodium falciparum. Front. Cell. Infect. Microbiol. 2017, 7, 320. [Google Scholar] [CrossRef] [Green Version]
- Basova, S.; Wilke, N.; Koch, J.C.; Prokop, A.; Berkessel, A.; Pradel, G.; Ngwa, C.J. Organoarsenic Compounds with in vitro Activity against the Malaria Parasite Plasmodium falciparum. Biomedicines 2020, 8, 260. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Alhadrami, H.A.; El-Gendy, A.O.; Shamikh, Y.I.; Belbahri, L.; Hassan, H.M.; Rateb, M.E. Microbial natural products as potential inhibitors of SARS-CoV-2 main protease (Mpro). Microorganisms 2020, 8, 970. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Alhadrami, H.A.; El-Hawary, S.S.; Mohammed, R.; Hassan, H.M.; Rateb, M.E.; Bakeer, W. Discovery of two brominated oxindole alkaloids as Staphylococcal DNA gyrase and pyruvate kinase inhibitors via inverse virtual screening. Microorganisms 2020, 8, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Rt (min) | m/z [M − H]− | m/z [M + H]+ | Molecular Formula | Tentative Identification | Strain EG49 | Strain UR59 | Coculture | Bioactivity | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| 2.47 | 303.1341 | C16H18N2O4 | Mitomycin-K | - | + | + | antitumor | [20] | |
| 2.91 | 327.0866 | C18H14O6 | Fluostatin-B | - | - | + | antinociceptive | [22] | |
| 2.94 | 657.1821 | C32H33O15 | Actinosporin A | + | - | + | anti-trypanosomal | [23] | |
| 2.96 | 613.1926 | C31H33O13 | Actinosporin C | + | - | + | antioxidant | [24] | |
| 2.99 | 399.1075 | C21H18O8 | Daunomycinone | - | - | + | - | [48] | |
| 3.04 | 599.2125 | C31H34O12 | Actinosporin F | - | - | + | - | [49] | |
| 3.08 | 469.1492 | C25H24O9 | Actinosporin E | - | - | + | - | [49] | |
| 3.11 | 467.1336 | C25H22O9 | Actinosporin H | - | - | + | - | [49] | |
| 3.15 | 513.1399 | C26H25O11 | Atramycin A | - | - | + | antitumor | [26] | |
| 3.25 | 451.1389 | C25H22O8 | Actinosporin G | - | - | + | - | [49] | |
| 3.30 | 309.0757 | C18H12O5 | Lagumycin B | - | - | + | anticancer | [27] | |
| 3.76 | 178.0499 | C9H9NO3 | Erbstatin | - | - | + | anticancer | [30] | |
| 3.81 | 635.3315 | C36H48N2O8 | Ansatrienin A | - | - | + | antifungal | [32] | |
| 3.93 | 192.0655 | C10H11NO3 | Spoxazomicin C | - | - | + | anti-trypanosomal | [50] | |
| 4.12 | 661.3568 | C32H48N6O9 | Actinoramide B | - | - | + | antimalarial | [51] | |
| 4.17 | 293.1749 | C17H26O4 | Cineromycin-B | - | - | + | antibacterial | [52] | |
| 4.54 | 438.1974 | C19H27N5O7 | Heterobactin B | - | - | + | siderophore | [37] | |
| 6.41 | 451.1391 | C25H24O8 | Atramycin B | - | - | + | antitumor | [26] | |
| 6.34 | 444.2744 | C26H39NO5 | Piericidin-F | - | + | + | anticancer | [38] | |
| 6.91 | 488.2649 | C27H39NO7 | Migrastatin | - | + | + | anticancer | [53] | |
| 7.24 | 547.3455 | C24H46N6O8 | Proferrioxamine-A1 | - | - | + | siderophore | [40] | |
| 7.40 | 393.2640 | C23H38O5 | Protylonolide | - | - | + | antibiotic | [41] | |
| 7.58 | 683.4347 | C37H62O11 | 26-Deoxylaidlomycin | - | - | + | antibacterial | [54] | |
| 7.76 | 482.3687 | C25H47N5O4 | Rhodopeptin C1 | - | + | + | Antifungal | [33] | |
| 8.15 | 639.4084 | C35H58O10 | Kaimonolide B | - | + | + | plant growth inhibitor | [43] | |
| 9.09 | 496.3846 | C26H49N5O4 | Rhodopeptin C2 | - | + | + | antifungal | [34] | |
| 9.14 | 524.4156 | C28H53N5O4 | Rhodopeptin B5 | - | + | + | antifungal | [34] | |
| 9.80 | 315.0865 | C17H14O6 | Capillasterquinone B | - | - | + | NO production inhibitor | [55] | |
| 9.81 | 305.0810 | C19H12O4 | Tetrangulol | - | - | + | antibiotic | [56] | |
| 9.98 | 457.3141 | C25H44O7 | 8,15-Dideoxylankanolide | - | + | + | - | [44] | |
| 10.22 | 527.2022 | C27H31N2O9 | UK-2B | + | - | + | antifungal | [57] | |
| 11.01 | 629.4242 | C34H60O10 | Ferensimycin-A | - | - | + | antibiotic | [45] | |
| 11.23 | 523.3601 | C26H46N6O5 | Lucentamycin C | - | - | + | anticancer | [46] | |
| 11.28 | 917.6546 | C50H92O14 | Glucolipsin-A | - | - | + | glucokinase activator | [47] |
| Compound | IC50 Values (µg/mL) 1 |
|---|---|
| Coculture extract | 0.13 |
| 1 | 12.6 |
| 2 | 13.6 |
| 3 | 11.2 |
| 4 | >50 |
| 5 | 9.7 |
| 6 | 9.2 |
| 7 | >50 |
| 8 | >50 |
| Chloroquine | 0.022 |
| Compound | Binding Energy (kcal/mol) | H-Bonding | Hydrophobic Interactions |
|---|---|---|---|
| 1 | −8.9 | ARG-330 | ALA-446, GLU-500, LYS-607 |
| 2 | −8.3 | ASP-450, SER-454, MET-475 | ARG-330, HIS-338, ASP-450, GLU-458, GLU-500 |
| 3 | −10.3 | GLU-493 | ARG-330, PHE-342, ASP-450, ARG-559 |
| 5 | −9.1 | GLU-500, THR-337 | ARG-330, HIS-338, PHE-342, ARG-559 |
| 6 | −9.0 | ARG-330, ASN-339 | ARG-330, HIS-338, PHE-342, GLU-500, ARG-559, LYS-607 |
| Co-crystalised ligand | −9.5 | ASN-339, GLY-556 | ARG-330, HIS-338, PHE-342 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhadrami, H.A.; Thissera, B.; Hassan, M.H.A.; Behery, F.A.; Ngwa, C.J.; Hassan, H.M.; Pradel, G.; Abdelmohsen, U.R.; Rateb, M.E. Bio-Guided Isolation of Antimalarial Metabolites from the Coculture of Two Red Sea Sponge-Derived Actinokineospora and Rhodococcus spp. Mar. Drugs 2021, 19, 109. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020109
Alhadrami HA, Thissera B, Hassan MHA, Behery FA, Ngwa CJ, Hassan HM, Pradel G, Abdelmohsen UR, Rateb ME. Bio-Guided Isolation of Antimalarial Metabolites from the Coculture of Two Red Sea Sponge-Derived Actinokineospora and Rhodococcus spp. Marine Drugs. 2021; 19(2):109. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020109
Chicago/Turabian StyleAlhadrami, Hani A., Bathini Thissera, Marwa H. A. Hassan, Fathy A. Behery, Che Julius Ngwa, Hossam M. Hassan, Gabriele Pradel, Usama Ramadan Abdelmohsen, and Mostafa E. Rateb. 2021. "Bio-Guided Isolation of Antimalarial Metabolites from the Coculture of Two Red Sea Sponge-Derived Actinokineospora and Rhodococcus spp." Marine Drugs 19, no. 2: 109. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020109