
Using Bacillus subtilis as a Host Cell to Express an Antimicrobial Peptide from the Marine Chordate Ciona intestinalis

Abstract
:1. Introduction
2. Results
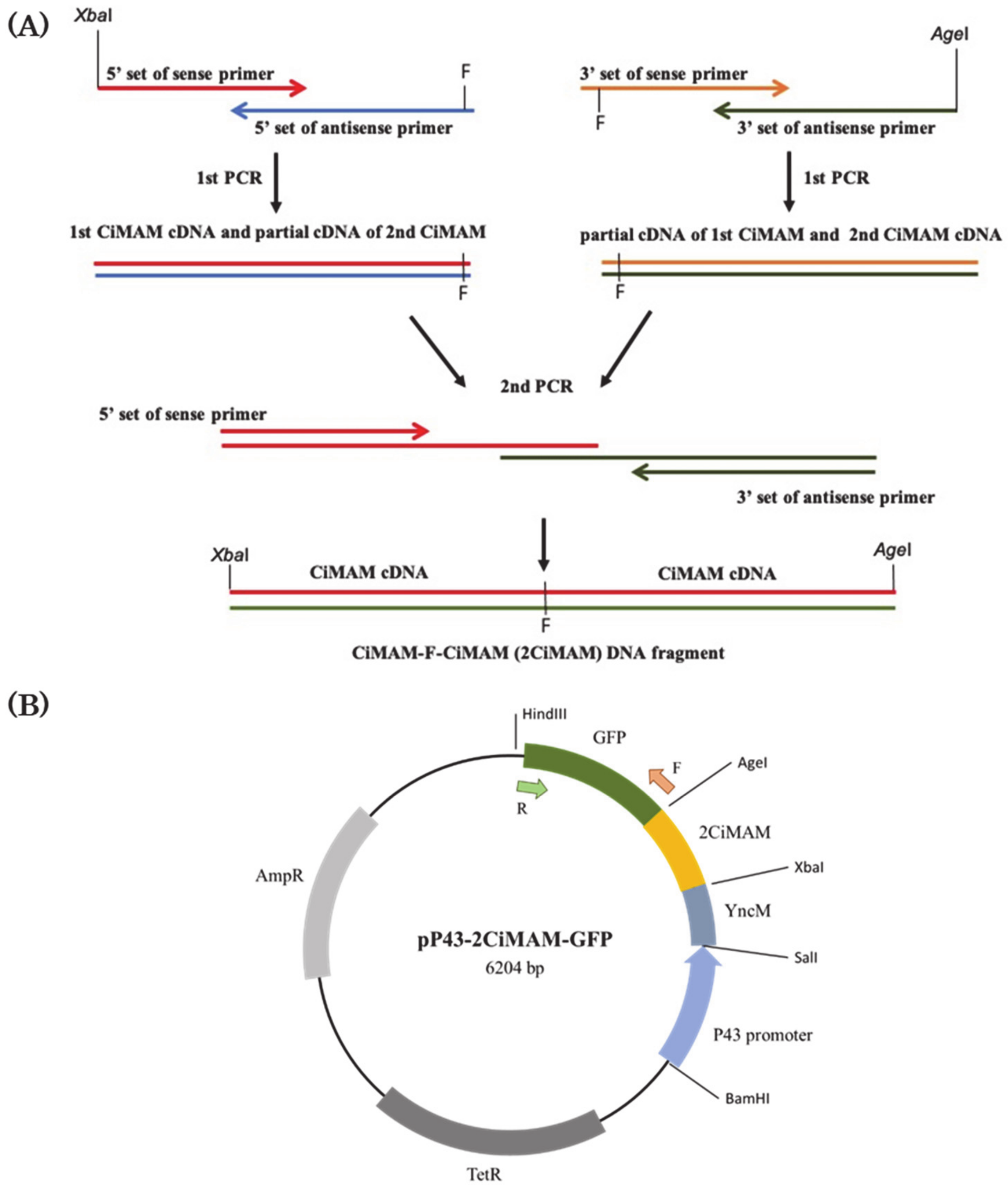
2.1. Construction of Expression Vector
2.2. Gene Transfer and Screening
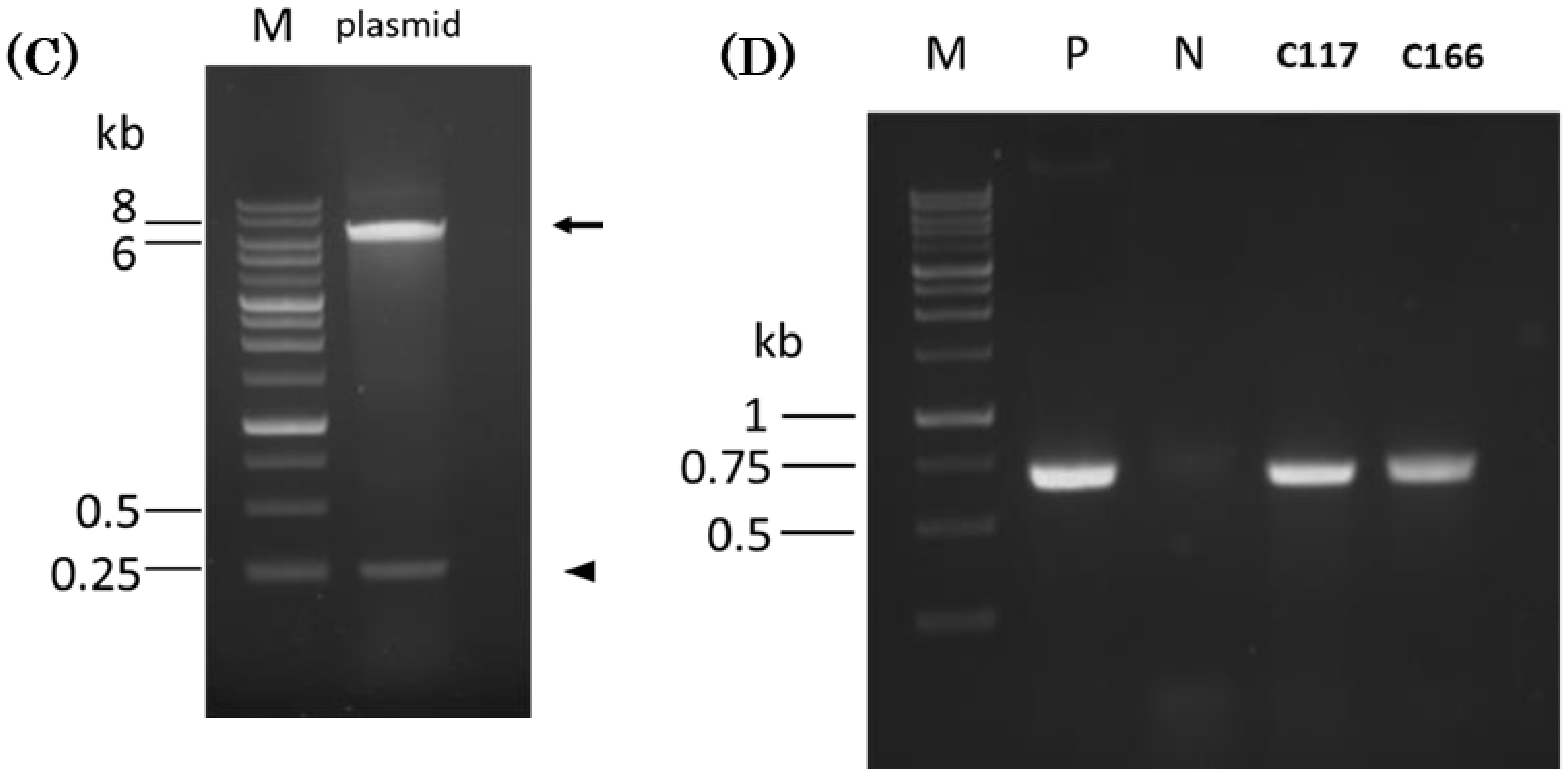
2.3. Using PCR to Detect the Exogenous Plasmid Transferred into B. subtilis Host Cells
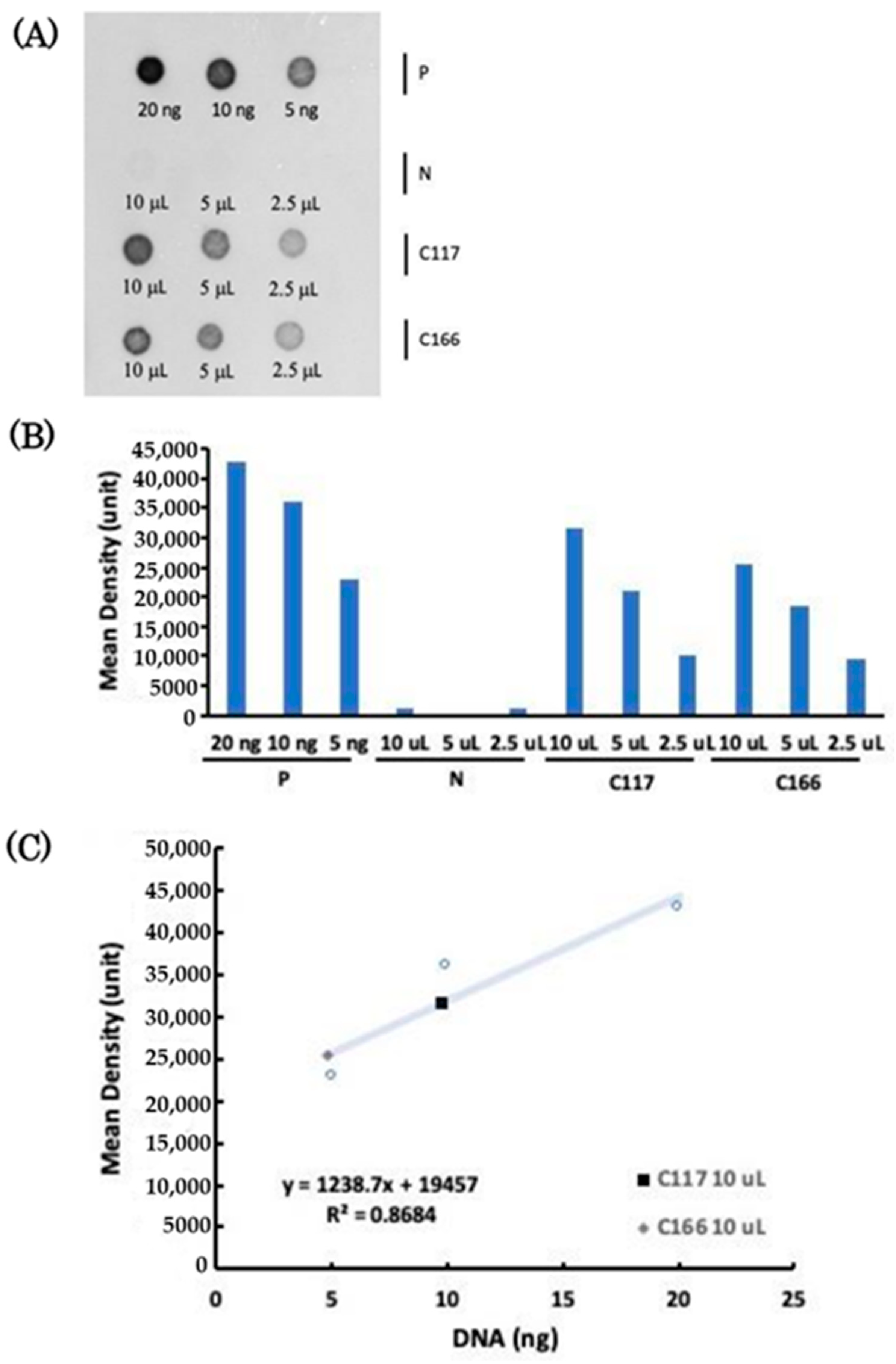
2.4. Copy Number of Plasmid Transferred to the Transgenic B. subtilis Strains
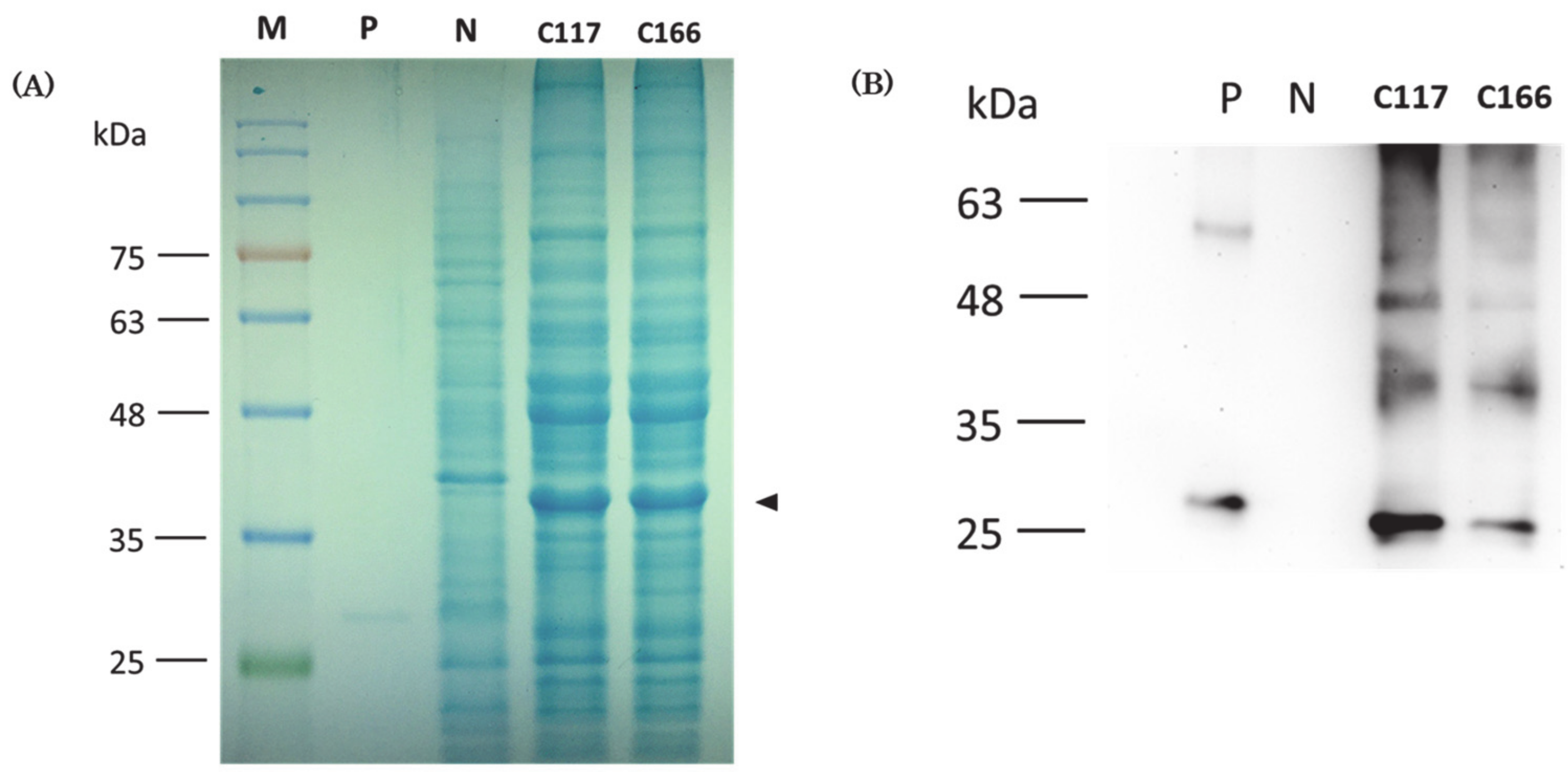
2.5. Western Blot Analysis to Prove That Recombinant Protein Containing CiMAM Was Produced by the Transgenic B. subtilis Strains
2.6. Using Amino Acid Sequencing to Further Confirm That Recombinant Proteins Were Produced by Transgenic Strains
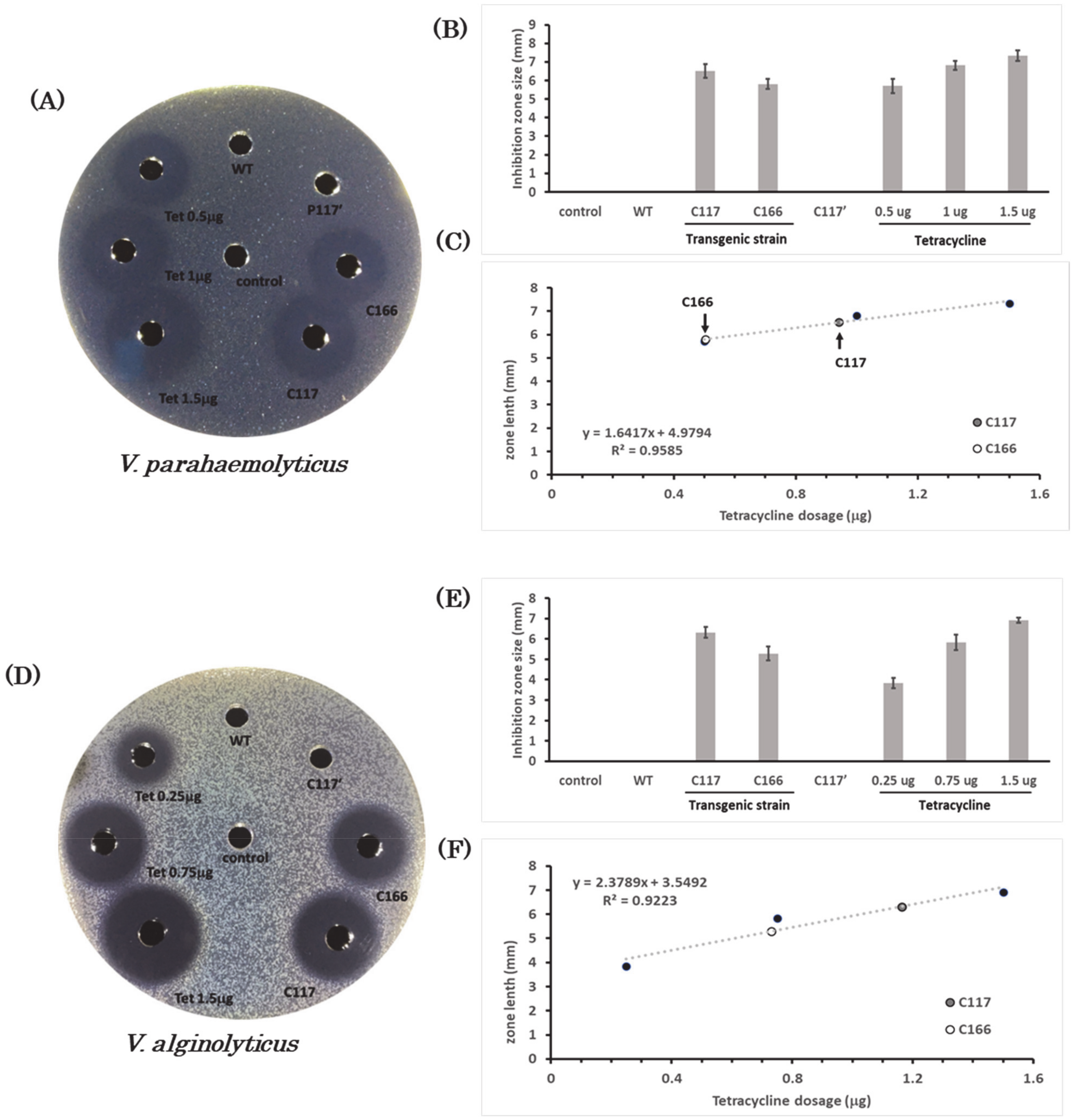
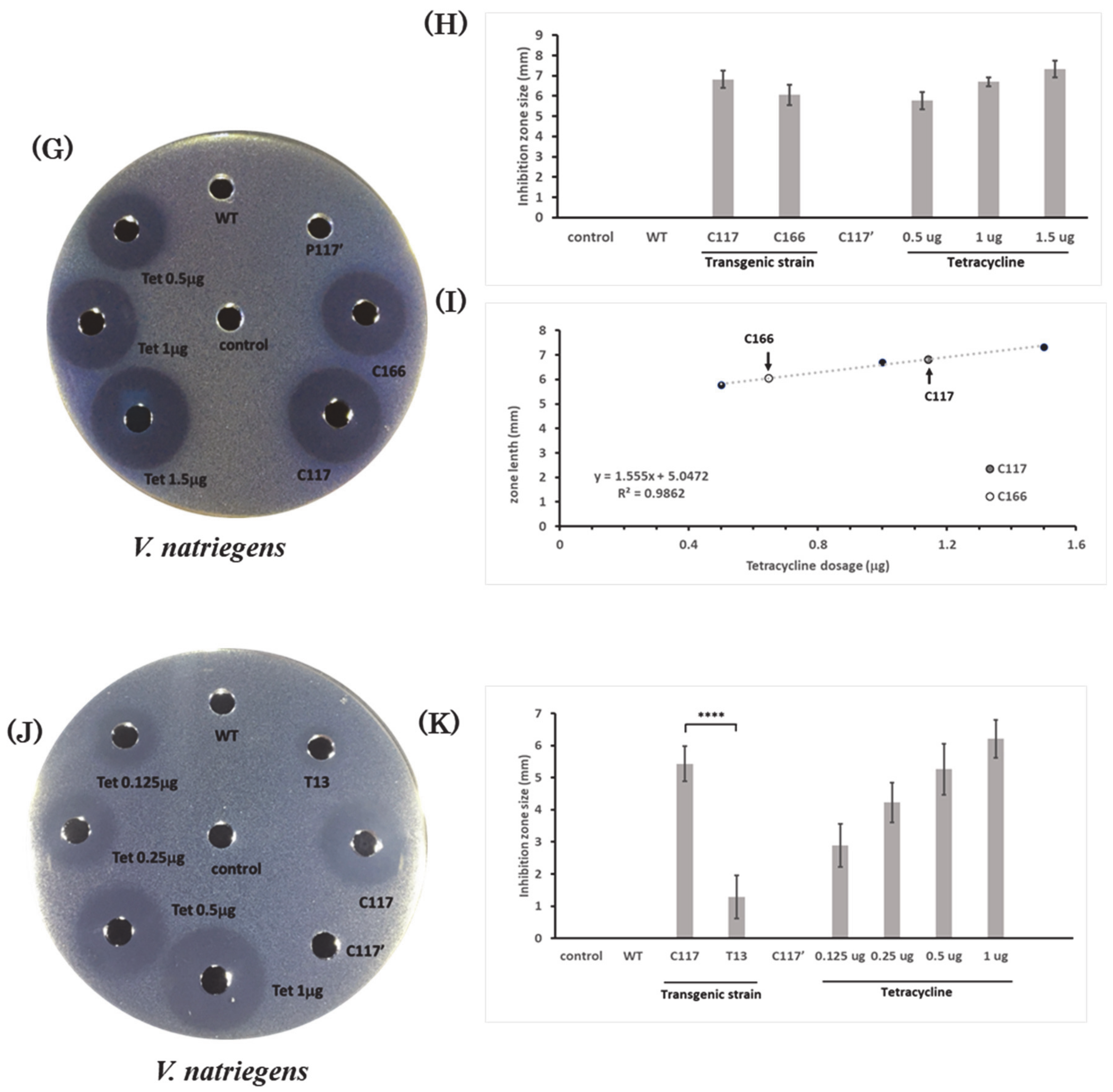
2.7. Bactericidal Activity of the Extracts from Transgenic B. subtilis Strains against Halophilic Pathogens
2.8. Bactericidal Activity of Recombinant CiMAM-Containing Extracts from Transgenic Strains C117 and C166 against the Euryhaline Pathogens
2.9. Bactericidal Activity of Recombinant CiMAM-Containing Extracts from Transgenic Strains C117 and C166 against Common Freshwater Pathogens
3. Discussion
3.1. Recombinant CiMAM -Expressing Transgenic B. subtilis Exhibits Bactericidal Activity against a Variety of Bacteria
3.2. The Effect of Salinity on the Efficacy of Antimicrobial Peptides
3.3. Recombinant Fusion Protein CiMAM-F-CiMAM-GFP Expressed in Transgenic B. subtilis Strains
3.4. The GFP Fragment Cleaved from Recombinant Fusion Protein CiMAM-F-CiMAM-GFP Served as a Selection Marker for Screening Transformants
3.5. Correlation between Plasmid Copy Number and Antimicrobial Ability of Transgenic B. subtilis Strain
4. Materials and Methods
4.1. Bacterial Culture Condition
4.2. Construction of Expression Plasmid
4.3. Preparation of Competent Cells, Electroporation, and PCR Detection
4.4. Dot Blot Analysis and Copy Number of Plasmids
4.5. Protein Analysis by SDS-PAGE
4.6. Western Blot Analysis
4.7. Peptide Sequence of the CiMAM-F-CiMAM-GFP Recombinant Fusion Protein by LC-MS/MS
4.8. Bactericidal Agar Plate Assay
4.9. Comparison of Bactercidal Efficacy against the Halophilic Pathogen between Recombinant Lactoferricin and CiMAM AMPs Produced by Transgenic Strain
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Ethical Statement
Conflicts of Interest
References
- Liu, W.Y.; Huang, S.L.; Chang, C.F.; Hsu, J.P.; Chen, M.L.; Lin, S.H. Guide to Diseases Control for Common Aquaculture Spices. In Fisheries Research Institute Special Publication, 1st ed.; Liu, W.Y., Ed.; Fisheries Research Institute, COA: Keelung, Taiwan, 2003; Volume 2, pp. 1–193. [Google Scholar]
- Moreno, E.; Prats, G.; Sabaté, M.; Pérez, T.; Johnson, J.R.; Andreu, A. Quinolone, fluoroquinolone and trimethoprim/sulfamethoxazole resistance in relation to virulence determinants and phylogenetic background among uropathogenic Escherichia coli. J. Antimicrob. Chemother. 2006, 57, 204–211. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente-Núñez, C.; Reffuveille, F.; Fernández, L.; Hancock, R.E. Bacterial biofilm development as a multicellular adaptation: Antibiotic resistance and new therapeutic strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef]
- Linnaeus, C. Systema Naturae Per Regna Tria Naturae: Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, Locis, 12th ed.; Holmiae: Stockholm, Sweden, 1767; Volume 1, pp. 533–1327. [Google Scholar]
- Leo, G.; Patricolo, E.; Frittitta, G. Fine structure of the tunic of Ciona intestinalis L. II. Tunic morphology, cell distribution and their functional importance. Acta Zool. 1981, 62, 259–271. [Google Scholar] [CrossRef]
- Dahlberg, C.; Auger, H.; Dupont, S.; Sasakura, Y.; Thorndyke, M.; Joly, J.S. Refining the Ciona intestinalis model of central nervous system regeneration. PLoS ONE 2009, 4, e4458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedders, H.; Michalek, M.; Grötzinger, J.; Leippe, M. An exceptional salt-tolerant antimicrobial peptide derived from a novel gene family of haemocytes of the marine invertebrate Ciona intestinalis. Biochem. J. 2008, 416, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedders, H.; Podschun, R.; Leippe, M. The antimicrobial peptide Ci-MAM-A24 is highly active against multidrug-resistant and anaerobic bacteria pathogenic for humans. Int. J. Antimicrob. Agents 2010, 36, 264–266. [Google Scholar] [CrossRef] [Green Version]
- Jena, P.; Mishra, B.; Leippe, M.; Hasilik, A.; Griffiths, G.; Sonawane, A. Membrane-active antimicrobial peptides and human placental lysosomal extracts are highly active against mycobacteria. Peptides 2011, 32, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Schlusselhuber, M.; Humblot, V.; Casale, S.; Méthivier, C.; Verdon, J.; Leippe, M.; Berjeaud, J.M. Potent antimicrobial peptides against Legionella pneumophila and its environmental host, Acanthamoeba castellanii. Appl. Microbiol. Biotechnol. 2015, 99, 4879–4891. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.A.; Khaneja, R.; Tam, N.M.; Cazzato, A.; Tan, S.; Urdaci, M.; Brisson, A.; Gasbarrini, A.; Barnes, I.; Cutting, S.M. Bacillus subtilis isolated from the human gastrointestinal tract. Res. Microbiol. 2009, 160, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions: Bacillus subtilis antibiotics. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Lin, S.C.; Chen, C.W.; Tzeng, D.S. The effect of microorganism bioagent for biological control of rice leaf blight disease and influence of rice agriculture characters. Bull. Taichung Dist. Agric. Res. Ext. Stat. 2003, 81, 65–77. [Google Scholar]
- Hsieh, F.C.; Li, M.C.; Kao, S.S. Evaluation of the inhibition activity of Bacillus subtilis-based products and their related metabolites against pathogenic fungi in Taiwan. Plant Protect. Bull. 2003, 45, 155–162. [Google Scholar]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.C.; Hung, C.W.; Lin, C.Y.; Shih, C.H.; Tsai, H.J. Oral administration of transgenic biosafe microorganism containing antimicrobial peptide enhances the survival of tilapia fry infected bacterial pathogen. Fish Shellfish. Immunol. 2019, 95, 606–616. [Google Scholar] [CrossRef]
- Wakabayashi, H.; Bellamy, W.; Takase, M.; Tomita, M. Inactivation of Listeria monocytogenes by Lactoferricin, a potent antimicrobial peptide derived from cow’s milk. J. Food Prot. 1992, 55, 238–240. [Google Scholar] [CrossRef]
- Huber, C.; Klimant, I.; Krause, C.; Werner, T.; Mayr, T.; Wolfbeis, O.S. Optical sensor for seawater salinity. Anal. Bioanal. Chem. 2000, 368, 196–202. [Google Scholar] [CrossRef]
- Chou, K.C.; Maggiora, G.M.; Nemethy, G.; Scheraga, H.A. Energetics of the structure of the four-alpha-helix bundle in proteins. Proc. Natl. Acad. Sci. USA 1988, 85, 4295–4299. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.C.; Yeung, J.C.; Duan, Y.; Ye, R.; Szarka, S.J.; Habibi, H.R.; Wong, S.L. Functional production and characterization of a fibrin-specific single-chain antibody fragment from Bacillus subtilis: Effects of molecular chaperones and a wall-bound protease on antibody fragment production. Appl. Environ. Microbiol. 2002, 68, 3261–3269. [Google Scholar] [CrossRef] [Green Version]
- Liew, P.X.; Wang, C.L.C.; Wong, S.L. Functional characterization and localization of a Bacillus subtilis sortase and its substrate and use of this sortase system to covalently anchor a heterologous protein to the B. subtilis cell wall for surface display. J. Bacteriol. 2012, 194, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, C.; Kobayashi, S.; Orii, H.; Watanabe, K.; Agata, K. Identification of two distinct muscles in the Planarian Dugesia japonica by their expression of myosin heavy chain genes. Zool. Sci. 1998, 15, 861–869. [Google Scholar] [CrossRef] [Green Version]
- Atkins, T. Biodegradation of poly(ethylene adipate) microcapsules in physiological media. Biomaterials 1998, 19, 61–67. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogens | Transgenic Strains: C117/C166 |
|---|---|
| LRE | |
| V. parahaemolyticus | y = 1.6417x + 4.9794, R2 = 0.9585 |
| V. alginolyticus | y = 2.3789x + 3.5492, R2 = 0.9223 |
| V. natriegens | y = 1.555x + 5.0472, R2 = 0.9862 |
| E. tarda | y = 2.2293x + 1.8928, R2 = 0.9588 |
| S. iniae | y = 1.71x + 3.9767, R2 = 0.994 |
| S. epidermis | y = 1.5157x + 0.7757, R2 = 0.9952 |
| Pathogens | Strains | |||
| C117 a | C166 a | T1 b | T13 b | |
| Promoter | ||||
| P43 | P43 | P43 | P43 | |
| Tandem repeatsc | ||||
| 2× CiMAM | 2× CiMAM | 3× LFB | 3× LFB | |
| Plasmid copy numberd | ||||
| 1057 | 349 | 931 | 647 | |
| S. epidermidis | 48 (Amp) | 41 (Amp) | 154 (Amp) | 130 (Amp) |
| E. tarda | 39 (Amp) | 37 (Amp) | 79 (Amp) | 33 (Amp) |
| V. parahaemolyticus | 47 (Tet) | 25 (Tet) | 30 (Tet) | 26 (Tet) |
| V. natriegens | 57 (Tet) | NA | NA | 9 (Tet) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.-C.; Tsai, J.-C.; Lin, C.-Y.; Hung, C.-W.; Sheu, J.-C.; Tsai, H.-J. Using Bacillus subtilis as a Host Cell to Express an Antimicrobial Peptide from the Marine Chordate Ciona intestinalis. Mar. Drugs 2021, 19, 111. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020111
Lee B-C, Tsai J-C, Lin C-Y, Hung C-W, Sheu J-C, Tsai H-J. Using Bacillus subtilis as a Host Cell to Express an Antimicrobial Peptide from the Marine Chordate Ciona intestinalis. Marine Drugs. 2021; 19(2):111. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020111
Chicago/Turabian StyleLee, Bing-Chang, Jui-Che Tsai, Cheng-Yung Lin, Chun-Wei Hung, Jin-Chuan Sheu, and Huai-Jen Tsai. 2021. "Using Bacillus subtilis as a Host Cell to Express an Antimicrobial Peptide from the Marine Chordate Ciona intestinalis" Marine Drugs 19, no. 2: 111. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020111