Exopolysaccharide from Porphyridium cruentum (purpureum) is Not Toxic and Stimulates Immune Response against Vibriosis: The Assessment Using Zebrafish and White Shrimp Litopenaeus vannamei

 ,
,
Abstract
:1. Introduction
2. Results
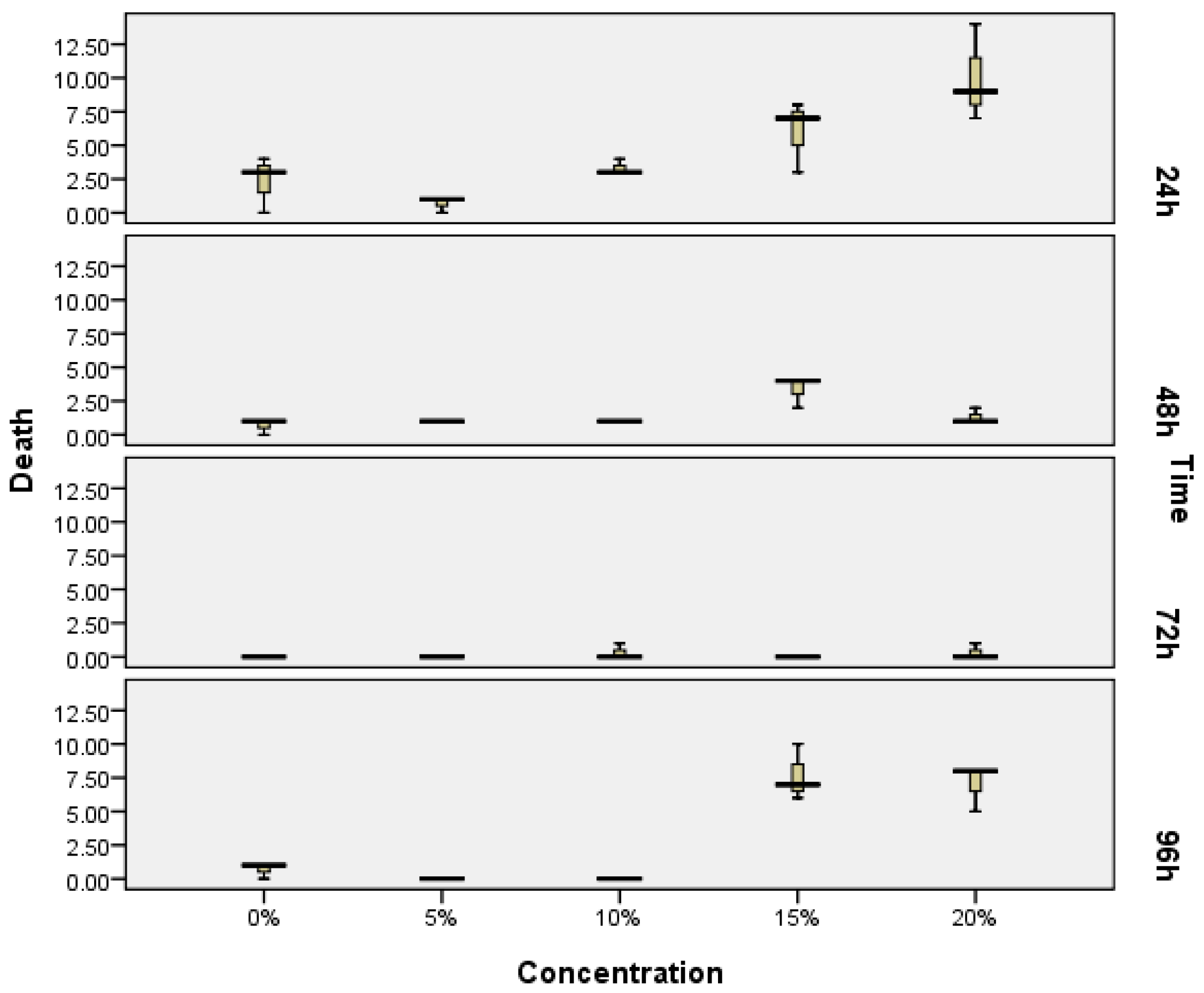
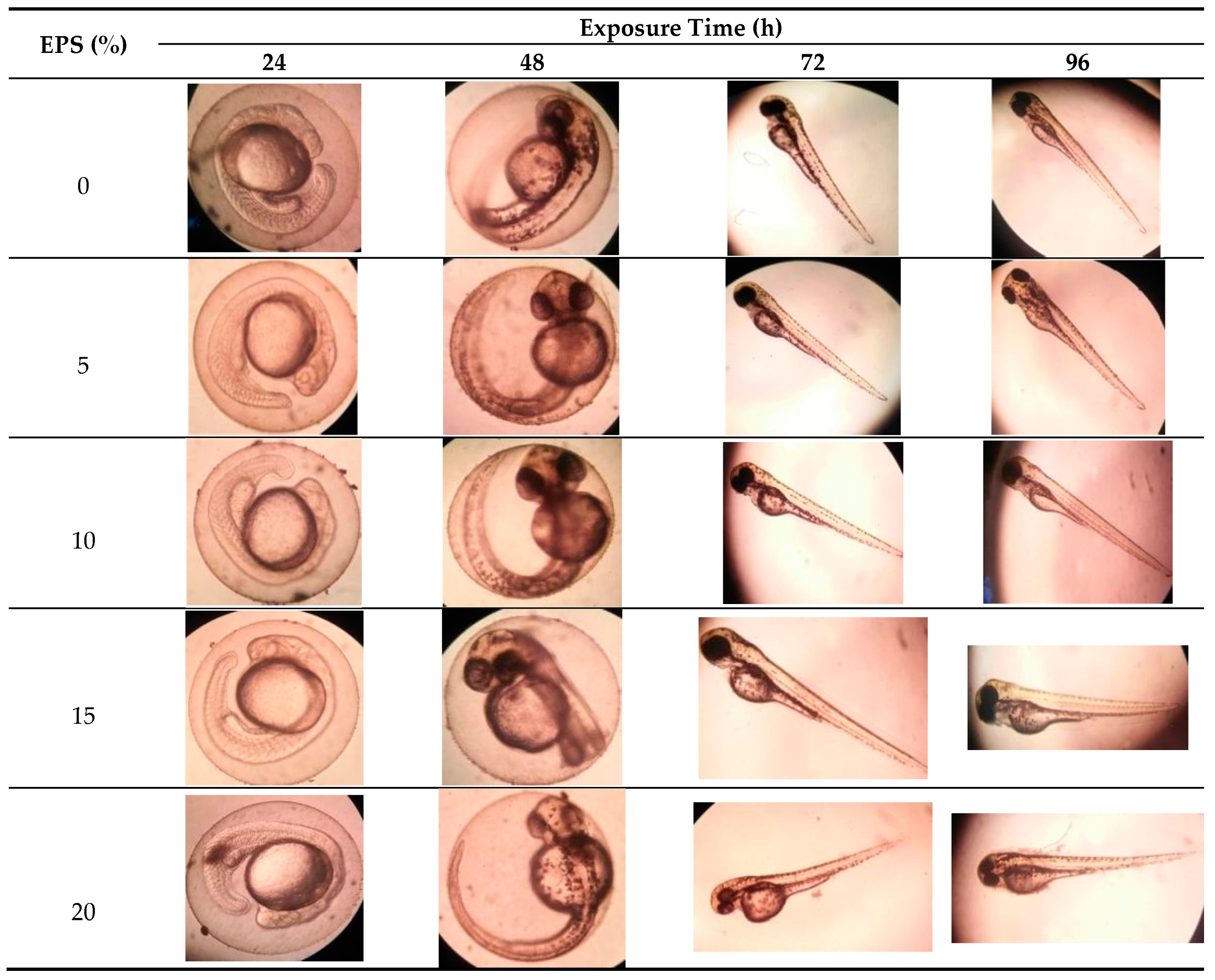
2.1. The Zebrafish Embryotoxicity Test (ZET)
2.2. Morphological View of Vibriosis in the Pacific White Shrimp (Litopenaeus vannamei)
2.3. Shrimp Immune Cells Number
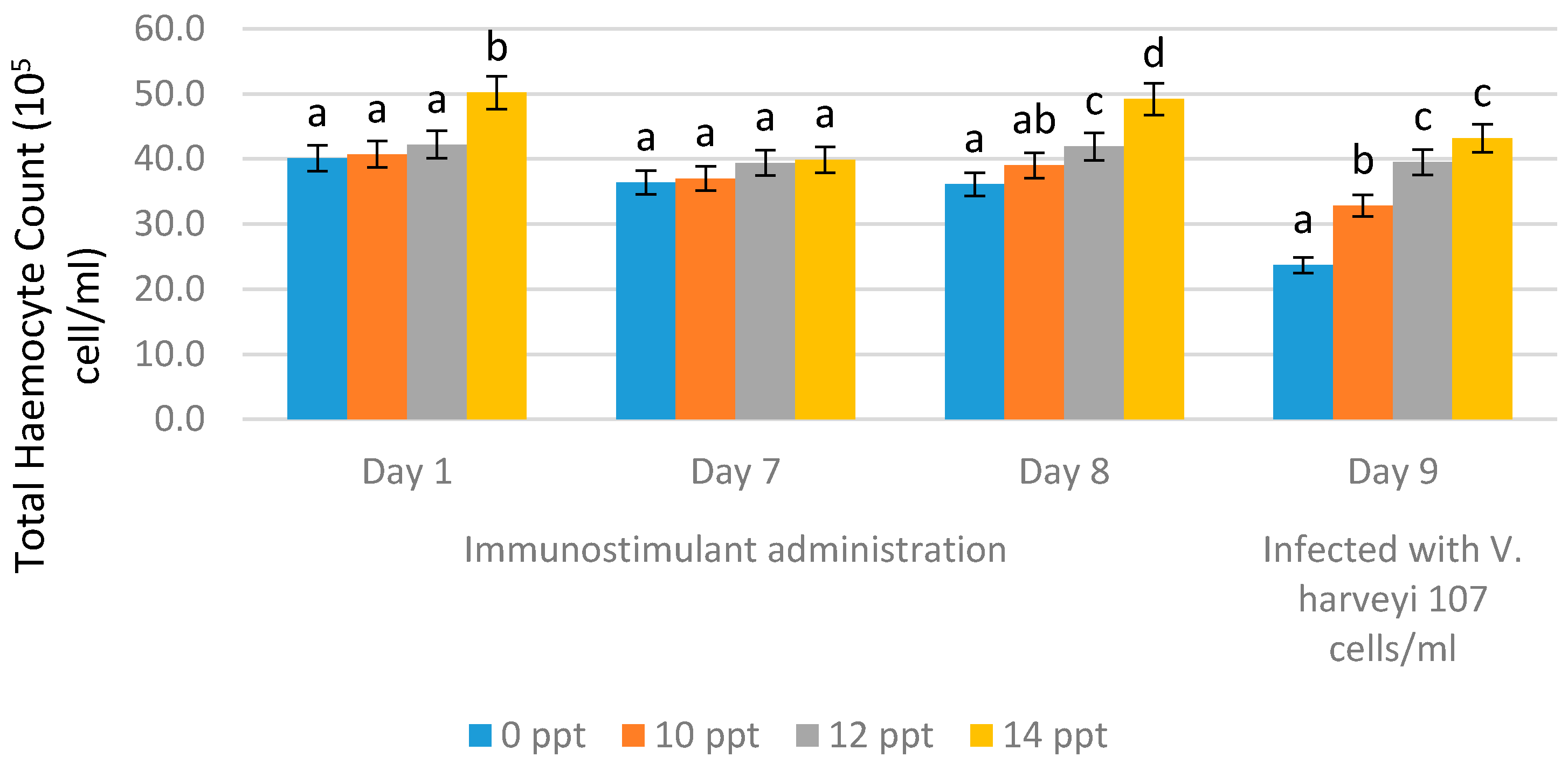
2.3.1. Total Hemocyte Count (THC)
2.3.2. Differential Hemocyte Count (DHC)
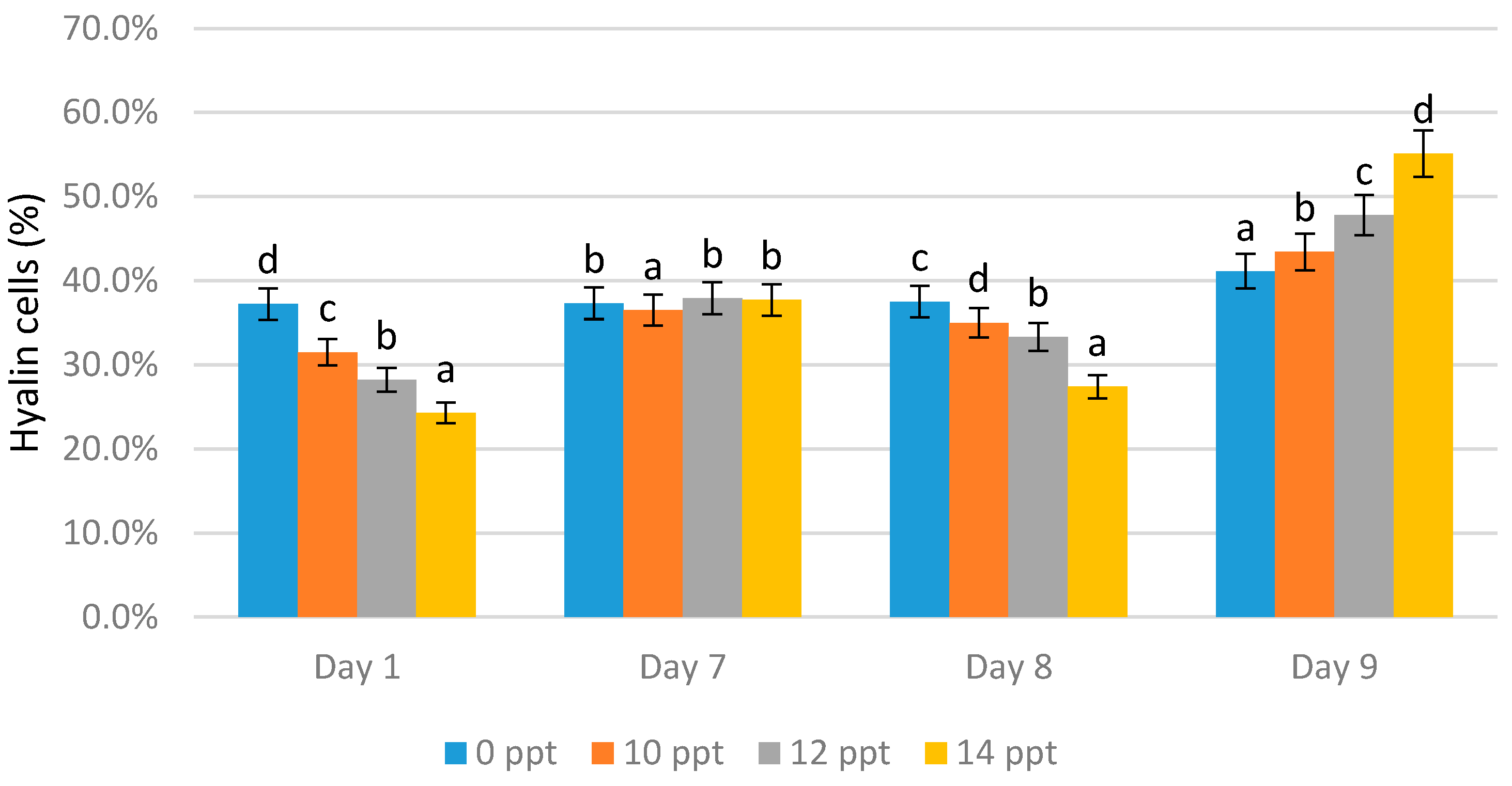
- Hyaline cells
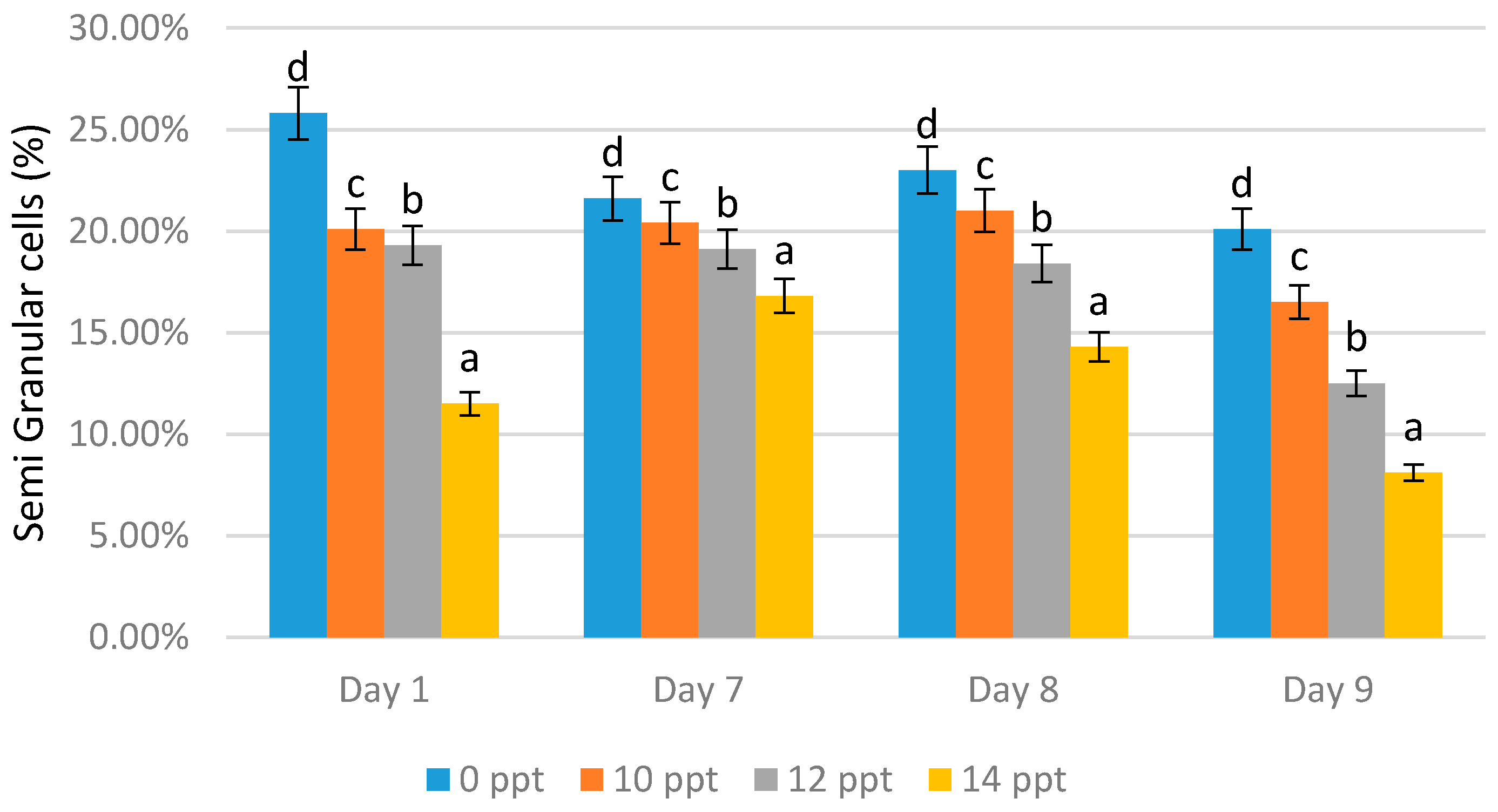
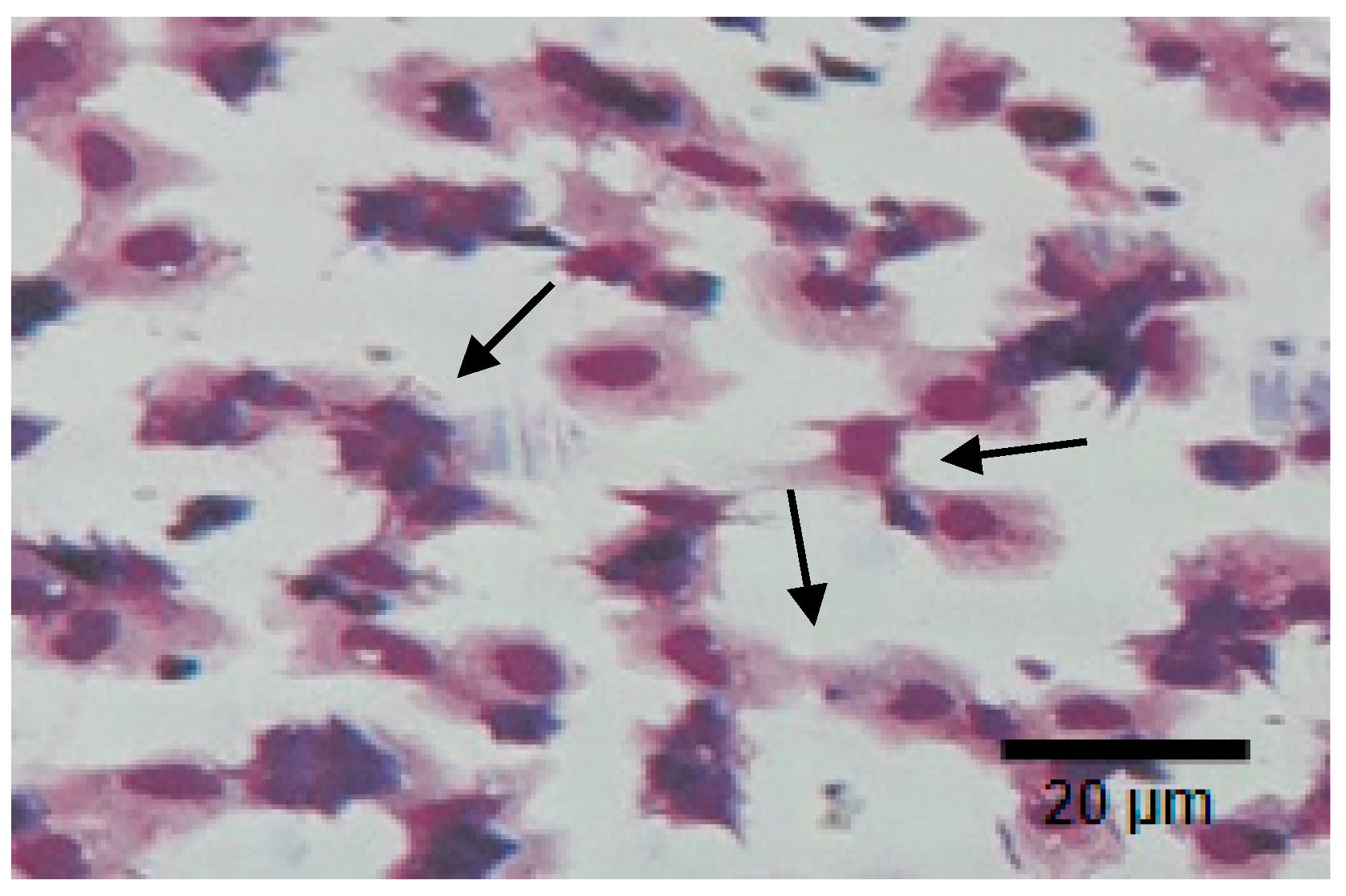
- Semi-granular and granular cells
2.4. Shrimp Immune Activity
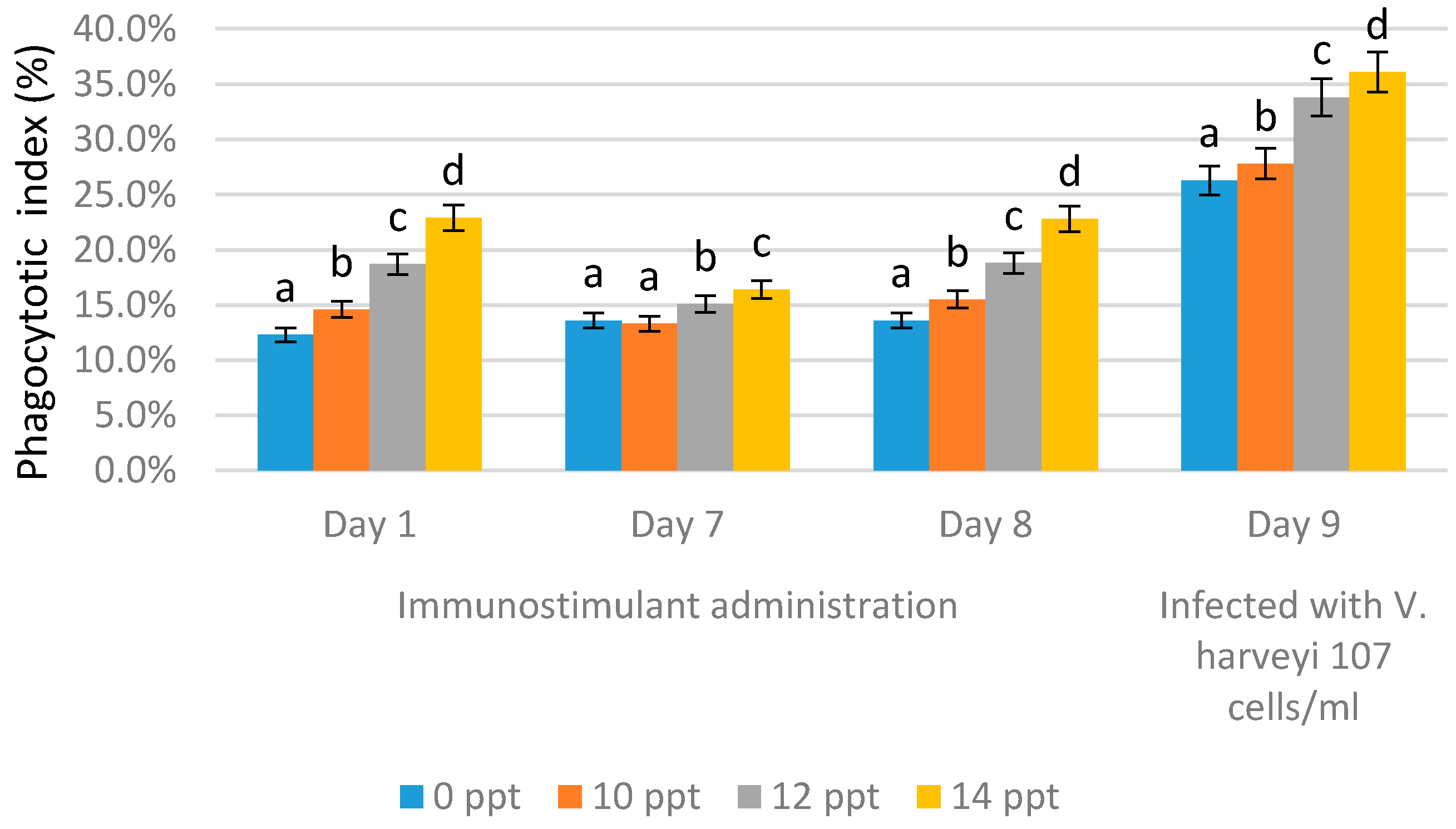
2.4.1. Phagocytic Activity (PA)
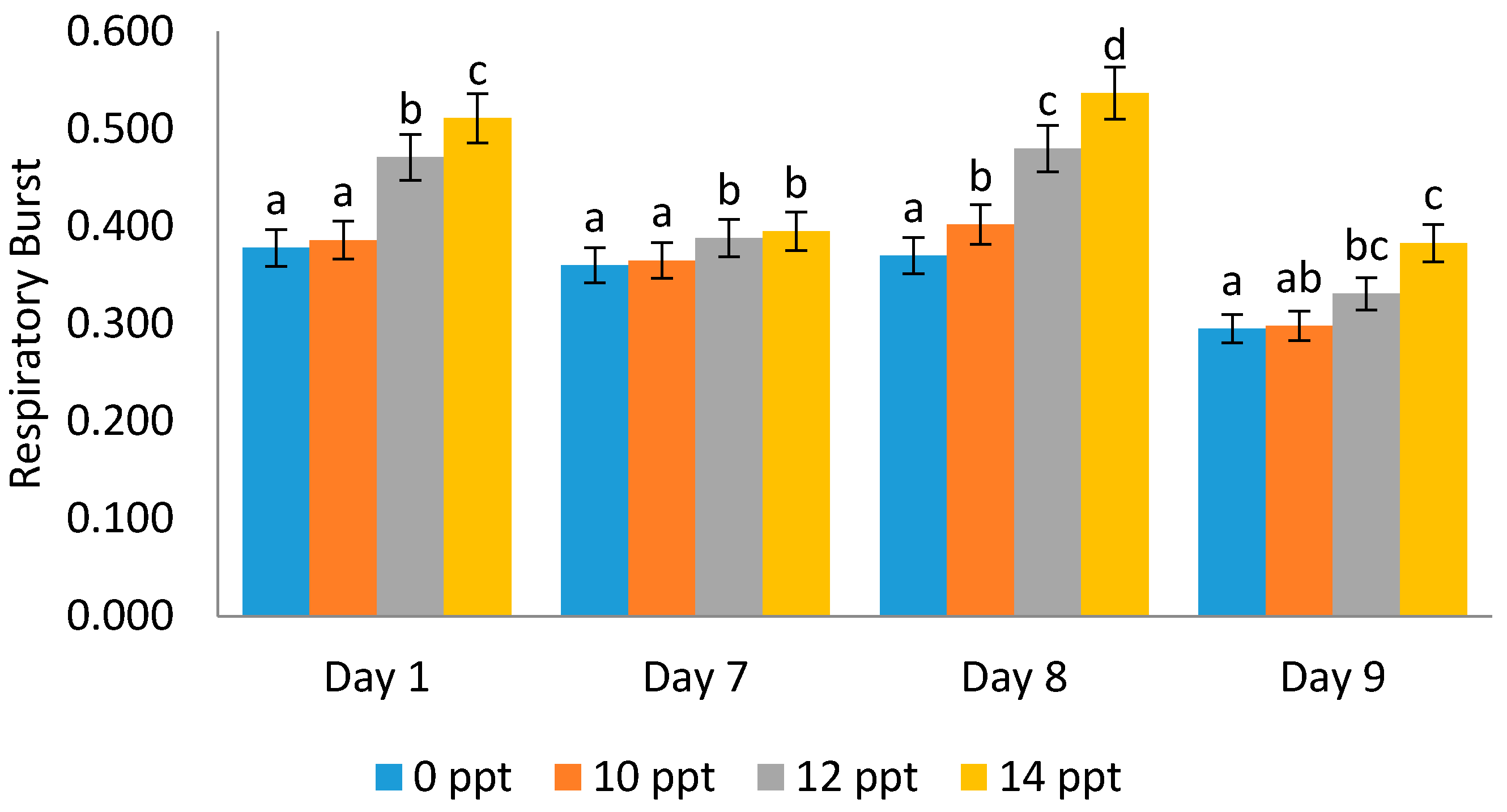
2.4.2. Respiratory Burst (RB) Activity
3. Discussion
4. Materials and Methods
4.1. General
4.2. Culture Condition
4.3. Extracellular Polysaccharides Extraction
4.4. Shrimp Treatment
4.5. Immune Parameters
4.6. Toxicity Test Using the ZET Method
4.7. Data Analyses
4.8. Ethical Consideration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- WWF-Indonesia. Budidaya Udang Vannamei, Tambak Semi Intensif Dengan Instalasi Pengolahan Air Limbah (IPAL); Better Management Practices; WWF-Indonesia: Jakarta, Indonesia, 2014; p. 38. ISBN 978-979-1461-38-2. [Google Scholar]
- Martinez, F.S. The Immune System of Shrimp. In Boletines Nicovita; Nicovita-ALICORP SAA Technical Service; SAA Technical: Johannesburg, South Africa, 2007; pp. 1–6. [Google Scholar]
- Widanarni; Meha, D.; Nuryati, S.; Sukendadan; Suwanto, A. Uji Patogenisitas Vibrio harveyi pada Larva Udang Windu Menggunakan Resistensi Rifampisin Sebagai Penanda Molekuler. J. Akuakultur Indones. 2004, 3, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Supriyadi, H.; Rukyani, A. The Use of Chemotherapuetic Agents for the Treatment of Bacterial Disease of Fish and Shrimp in Indonesia; Asian Fisheries Society: Manila, Philippines, 1992; pp. 515–517. [Google Scholar]
- Widowati, I.; Zainuri, M.; Kusumaningrum, H.P.; Maesaroh, Y.; Hardivillier, Y.; Leignel, V.; Bourgougnon, N.; Mouget, J.L. Identification of agents causing vibriosis in Litopenaeus vannamei shrimp culture in Kendal, Central Java, Indonesia and application of microalgae Dunaliella salina and Tetraselmis chui as bio-control agents against vibriosis. Aquac. Aquar. Conserv. Legis. 2018, 11, 101–107. [Google Scholar]
- Leung, K.M.; Yeung, K.W.; You, J.; Choi, K.; Zhang, X.; Smith, R.; Zhou, G.-J.; Yung, M.M.; Arias-Barreiro, C.; An, Y.-.J.; et al. Toward Sustainable Environmental Quality: Priority Research Questions for Asia. Environ. Toxicol. Chem. 2020, 39, 1485–1505. [Google Scholar] [CrossRef] [PubMed]
- Salzet, M. Vertebrate innate immunity resembles a mosaic of invertebrate immune responses. Trends Immunol. 2001, 22, 285–288. [Google Scholar] [CrossRef]
- Vargas-Albores, F.; Yepiz-Plascencia, G. Beta Glucan Binding protein and Its Role in Shrimp Immune Response. Aquaculture 2000, 191, 13–21. [Google Scholar] [CrossRef]
- Risjani, Y.; Yunianta Couteau, J.; Minier, C. Cellular immune responses and phagocytic activity of fishes exposed to pollution of volcano mud. Mar. Environ. Res. 2014, 96, 3–80. [Google Scholar] [CrossRef]
- Risjani, Y.; Loppion, G.; Couteau, J.; Yunianta, Y.; Widowati, I.; Hermawati, A.; Minier, C. Genotoxicity in the rivers from the Brantas catchment (East Java, Indonesia): Occurrence in sediments and effects in Oreochromis niloticus (Linnæus 1758). Environ. Sci. Pollut. Res. 2020, 27, 1–9. [Google Scholar] [CrossRef]
- Risjani, Y.; Santoso, D.R.; Couteau, J.; Hermawati, A.; Widowati, I.; Minier, C. Impact of anthropogenic activity and lusi-mud volcano on fish biodiversity at the Brantas Delta, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2020, 493, 012007. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Borowitzka, L.J. Micro-Algal Biotechnology; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Garcia, I.R.; Guerrero, J.L.G. Evaluation of the antioxidant activity of three microalgal species for use as dietary supplements and in the preservation of foods. Food Chem. 2008, 108, 1023–1026. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Influence of sulphate on the composition and antibacterial and antiviral properties of the exopolysaccharide from Porphyridium cruentum. Life Sci. 2014, 101, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Mutmainnah, N.; Risjani, Y.; Hertika, A.M.S. Growth Rate and Chemical Composition of Secondary Metabolite Extracellular Polysaccharide (EPS) Microalga Porphyridium cruentum. J. Exp. Life Sci. 2018, 8, 97–102. [Google Scholar] [CrossRef]
- Raposo, M.F.D.J.; De Morais, R.M.S.C.; Bernardo de Morais, A.M.M. Bioactivity and applications of sulphated polysaccharides from marine microalgae. Mar. Drugs 2013, 11, 233–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parikh, A.; Madamwar, D. Partial characterization of extracellular polysaccharides from cyanobacteria. Bioresour. Technol. 2006, 97, 1822–1827. [Google Scholar] [CrossRef] [PubMed]
- Geresh, S.; Adin, I.; Yarmolinsky, E.; Karpasas, M. Characterization of the extracellular polysaccharide of Porphyridium sp.: Molecular weight determination and rheological properties. Carbohydr. Polym. 2002, 50, 183–189. [Google Scholar] [CrossRef]
- Sun, L. Preparation of Polysaccharides from Porphyridium cruentum and Their Biological Activities. Ph.D. Thesis, Dalian University of Technology, Dalian, China, 2010. [Google Scholar]
- Arad, S.M.; Adda, M.; Cohen, E. The potential of production of sulphated polysaccharides from Porphyridium. Plant Soil 1985, 89, 117–127. [Google Scholar] [CrossRef]
- Andrew, M.; Jayaraman, G. Structural features of microbial exopolysaccharides in relation to their antioxidant activity. Carbohydr. Res. 2020, 487, 107881. [Google Scholar] [CrossRef]
- Truong, L.; Harper, S.L.; Tanguay, R.L. Evaluation of embryotoxicity using the zebrafish model. In Drug Safety Evaluation; Humana Press: Totowa, NJ, USA, 2011; pp. 271–279. [Google Scholar]
- Parng, C.; Seng, W.L.; Semino, C.; McGrath, P. Zebrafish: A preclinical model for drug screening. Assay Drug Dev. Technol. 2002, 1, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, P.J.; Richardson, M.K.; Champagne, D.L. The use of the zebrafish model in stress research. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1432–1451. [Google Scholar] [CrossRef] [PubMed]
- Segner, H. Zebrafish (Danio rerio) as a model organism for investigating endocrine disruption. Comp. Biochem. Phys. C Toxicol. Pharmacol. 2009, 149, 187–195. [Google Scholar] [CrossRef]
- Dahm, R.; Geisler, R. Learning from small fry: The zebrafish as a genetic model organism for aquaculture fish species. Mar. Biotech. 2006, 8, 329–345. [Google Scholar] [CrossRef]
- Spitsbergen, J.M.; Kent, M.L. The state of the art of the zebrafish model for toxicology and toxicologic pathology research Advantages and current limitations. Toxicol. Pathol. 2003, 31 (Suppl. 1), 62–87. [Google Scholar] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Busquet, F.; Strecker, R.; Rawlings, J.M.; Belanger, S.E.; Braunbeck, T.; Carr, G.J.; Halder, M. OECD validation study to assess intra-and inter-laboratory reproducibility of the zebrafish embryo toxicity test for acute aquatic toxicity testing. Regul. Toxicol. Pharmacol. 2014, 69, 496–511. [Google Scholar] [CrossRef]
- Tseng, C.C.; Chu, T.W.; Danata, R.H.; Risjani, Y.; Shih, H.T.; Hu, S.Y. Hepcidin-Expressing Fish Eggs as A Novel Food Supplement to Modulate Immunity against Pathogenic Infection in Zebrafish (Daniorerio). Sustainability 2020, 12, 4057. [Google Scholar] [CrossRef]
- Toban, M.H.; Risjani, Y. Administration of marine algae (Gracilaria verrucosa) immunostimulant enhances some innate immune parameters in black tiger shrimp (Penaeus monodon fabricus) against vibrio harveyi infection. J. Appl. Sci. Res. 2012, 8, 1052–1058. [Google Scholar]
- Dangeubun, J.L.; Hardoko, A.S.; Risjani, Y. The Effect of Treatment of Alstonia acuminata Bark-Based Active Compound on the Hematology and Histology of Tiger Grouper Fish (Epinephelus fuscoguttatus). J. Appl. Biotechnol. 2013, 1, 11–24. [Google Scholar] [CrossRef]
- Risjani, Y.; Witkowski, A.; Kryk, A.; Yunianta Górecka, E.; Krzywda, M.; Safitri, I.; Sapar, A.; Dąbek, P.; Arsad, S.; Gusev, E.; et al. Indonesian coral reef habitats reveal exceptionally high species richness and biodiversity of diatom assemblages. Estuar. Coast. Shelf Sci. 2021. under review. [Google Scholar]
- Sun, L.C.; Wang, Q.; Shi, C.M. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef]
- Trianto, A.; Wibowo, E.; Suryono, S.; Sapta, R. Ekstrak Daun Mangrove Aegiceras corniculatum sebagai Antibakteri Vibrio harveyi dan Vibrio parahaemolyticus. Ilmu Kelaut. 2004, 9, 186–189. [Google Scholar]
- Kavitha, M.D.; Shree, M.S.; Vidyashankar, S.; Sarada, R. Acute and subchronic safety assessment of Porphyridium purpureum biomass in the rat model. J. Appl. Phycol. 2016, 28, 1071–1083. [Google Scholar] [CrossRef]
- Jayasree, L.; Janakiram, P.; Madhavi, R. Characterization of Vibrio spp. Associated with Diseased Shrimp from Culture Ponds of Andhra Pradesh (India). J. World Aquac. Soc. 2006, 37, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Rozik, M. Pengaruh Imunostimulan OMP terhadap Histopatologi Hepatopankreas Udang Windu (Peneaus monodon fabricus) pasca Uji Tantang dengan Vibrio Harveyi. J. Trop. Fish. 2014, 10, 750–755. [Google Scholar]
- Hsieh, S.L.; Ruan, Y.H.; Li, Y.C.; Hsieh, P.S.; Hu, C.H.; Kuo, C.M. Immune and Physiological Responses in Pacific White Shrimp (Panaeus vannamei) to Vibrio alginolyticus. Aquaculture 2008, 275, 335–341. [Google Scholar] [CrossRef]
- Li, T.; Yan, D.; Wang, X.; Zhang, L.; Chen, P. Hemocyte Changes During Immune Melanization in Bombyx Mori Infected with Escherichia coli. Insects 2019, 10, 301. [Google Scholar] [CrossRef] [Green Version]
- Stoepler, T.M.; Castillo, J.C.; Lill, J.T.; Eleftherianos, I. Hemocyte density increases with developmental stage in an immune-challenged forest caterpillar. PLoS ONE 2013, 8, e70978. [Google Scholar] [CrossRef] [Green Version]
- Sari, A.H.W.; Risjani, Y.; Mahendra, A.P.W. Efek Konsentrasi Sublethal Fenol Terhadap Total Haemocyte Count (THC) dan Histologi Insang Kepiting Bakau (Scylla serata). J. Exp. Life Sci. 2012, 2, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.; Le Moullac, G. State of the Art of Immunological Tools and Health Control of Penaeid Shrimp. Aquaculture 2000, 191, 109–119. [Google Scholar] [CrossRef]
- Johansson, M.W.; Soderhall, K. Cellular immunity in crustaceans and the proPO system. Parasitol. Today 1989, 5, 171–176. [Google Scholar] [CrossRef]
- Supamattaya, K.; Pongmaneerat, J.; Klowklieng, T. The Effect of β-glucan (Macro Gard) on Growth Performance, Immune Response and Disease Resistance in Black Tiger Shrimp, Penaeus monodon Fabricus. Songklanakarin J. Sci. Technol. 2000, 22, 677–688. [Google Scholar]
- Andrade, F.G.D.; Negreiro, M.C.C.D.; Levy, S.M.; Fonseca, I.C.D.B.; Moscardi, F.; Falleiros, Â.M.F. Hemocyte quantitative changes in Anticarsiagemmatalis (Lepidoptera: Noctuidae) larvae infected by AgMNPV. Braz. Arch. Biol. Technol. 2010, 53, 279–284. [Google Scholar] [CrossRef]
- Castro, R.; Zarra, I.; Lamas, J. Water-soluble seaweed extracts modulate the respiratory burst activity of turbot phagocytes. Aquaculture 2004, 229, 67–78. [Google Scholar] [CrossRef]
- Velmurugan, B.K.; Jiang, I.F.; Shih, H.Y.; Lee, D.N.; Weng, C.F. Respiratory Burst Activity in Head Kidney and Spleen Leukocytes of Tilapia (Oreochromis mossambicus) under Acute Osmotic Stress. Zool. Stud. 2012, 51, 1290–1297. [Google Scholar]
- Kanjana, K.; Radtanatip, T.; Asuvapongpatana, S.; Withyachumnarnkul, B.; Wongprasert, K. Solvent extracts of the red seaweed Gracilariafisheri prevent Vibrio harveyi infections in the black tiger shrimp Penaeusmonodon. Fish Shellfish Immunol. 2011, 30, 389–396. [Google Scholar] [CrossRef]
- Esquer-Miranda, E.; Nieves-Soto, M.; Rivas-Vega, M.E.; Miranda-Baeza, A.; Piña-Valdez, P. Effects of methanolicmacroalgae extracts from Caulerpasertularioides and Ulvalactuca on Litopenaeusvannamei survival in the presence of Vibrio bacteria. Fish Shellfish Immunol. 2016, 51, 346–350. [Google Scholar] [CrossRef]
- Risjani, Y.; Abidin, G. Genetic diversity and similarity between green and brown morphotypes of Kappaphycus alvarezii using RAPD. J. Appl. Phycol. 2020, 32, 2253–2260. [Google Scholar] [CrossRef]
- Wootton, E.C.; Dyrynda, E.A.; Pipe, R.K.; Ratcliffe, N.A. Comparisons of PAH-induced immunomodulation in three bivalve molluscs. Aquat. Toxicol. 2003, 65, 13–25. [Google Scholar] [CrossRef]
- Anderson, D.P.; Siwicki, A.K. Basic haematology and serology for fish health programs. In Diseases in Asian Aquaculture II; Shariff, M., Arthur, J.R., Subasinghe, R.P., Eds.; Fish Health Section, Asian Fisheries Society: Manila, Philippines, 1995; pp. 185–202. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EPS (%) | Mean | 5% | 0% | 10% | 15% | 20% | Notation |
|---|---|---|---|---|---|---|---|
| 5% | 1.7 | a | |||||
| 0% | 3.7 | 2 | a | ||||
| 10% | 4.7 | 3 | 1 | b | |||
| 15% | 17 | 15.3 | 13.3 | 12.3 | c | ||
| 20% | 18 | 16.3 | 14.3 | 13.3 | 1 | c |
| EPS. Concentration | Replication | Exposure Time | |||||||
|---|---|---|---|---|---|---|---|---|---|
| N | 24 h | N | 48 h | N | 72 h | N | 96 h | ||
| 1 | 0 | 0 | 0 | 0 | 12 | 81 | 19 | 76 | |
| 0% | 2 | 0 | 0 | 4 | 70 | 14 | 72 | 15 | 86 |
| 3 | 0 | 0 | 0 | 0 | 7 | 83 | 15 | 84 | |
| 1 | 0 | 0 | 0 | 0 | 13 | 86 | 19 | 83 | |
| 5% | 2 | 0 | 0 | 4 | 85 | 17 | 85 | 18 | 84 |
| 3 | 0 | 0 | 2 | 85 | 18 | 87 | 18 | 83 | |
| 1 | 0 | 0 | 2 | 72 | 17 | 70 | 17 | 84 | |
| 10% | 2 | 0 | 0 | 0 | 0 | 13 | 79 | 14 | 84 |
| 3 | 0 | 0 | 4 | 78 | 16 | 76 | 16 | 85 | |
| 1 | 0 | 0 | 0 | 0 | 9 | 60 | 9 | 82 | |
| 15% | 2 | 0 | 0 | 1 | 80 | 8 | 79 | 8 | 87 |
| 3 | 0 | 0 | 0 | 0 | 14 | 79 | 16 | 88 | |
| 1 | 0 | 0 | 0 | 0 | 8 | 53 | 10 | 78 | |
| 20% | 2 | 0 | 0 | 0 | 0 | 7 | 68 | 12 | 78 |
| 3 | 0 | 0 | 0 | 0 | 0 | 70 | 5 | 77 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Risjani, Y.; Mutmainnah, N.; Manurung, P.; Wulan, S.N.; Yunianta. Exopolysaccharide from Porphyridium cruentum (purpureum) is Not Toxic and Stimulates Immune Response against Vibriosis: The Assessment Using Zebrafish and White Shrimp Litopenaeus vannamei. Mar. Drugs 2021, 19, 133. https://0-doi-org.brum.beds.ac.uk/10.3390/md19030133
Risjani Y, Mutmainnah N, Manurung P, Wulan SN, Yunianta. Exopolysaccharide from Porphyridium cruentum (purpureum) is Not Toxic and Stimulates Immune Response against Vibriosis: The Assessment Using Zebrafish and White Shrimp Litopenaeus vannamei. Marine Drugs. 2021; 19(3):133. https://0-doi-org.brum.beds.ac.uk/10.3390/md19030133
Chicago/Turabian StyleRisjani, Yenny, Nurul Mutmainnah, Praprianita Manurung, Siti Narsito Wulan, and Yunianta. 2021. "Exopolysaccharide from Porphyridium cruentum (purpureum) is Not Toxic and Stimulates Immune Response against Vibriosis: The Assessment Using Zebrafish and White Shrimp Litopenaeus vannamei" Marine Drugs 19, no. 3: 133. https://0-doi-org.brum.beds.ac.uk/10.3390/md19030133