
Induction of Apoptotic Cell Death in Human Leukemia U937 Cells by C18 Hydroxy Unsaturated Fatty Acid Isolated from Red Alga Tricleocarpa jejuensis


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
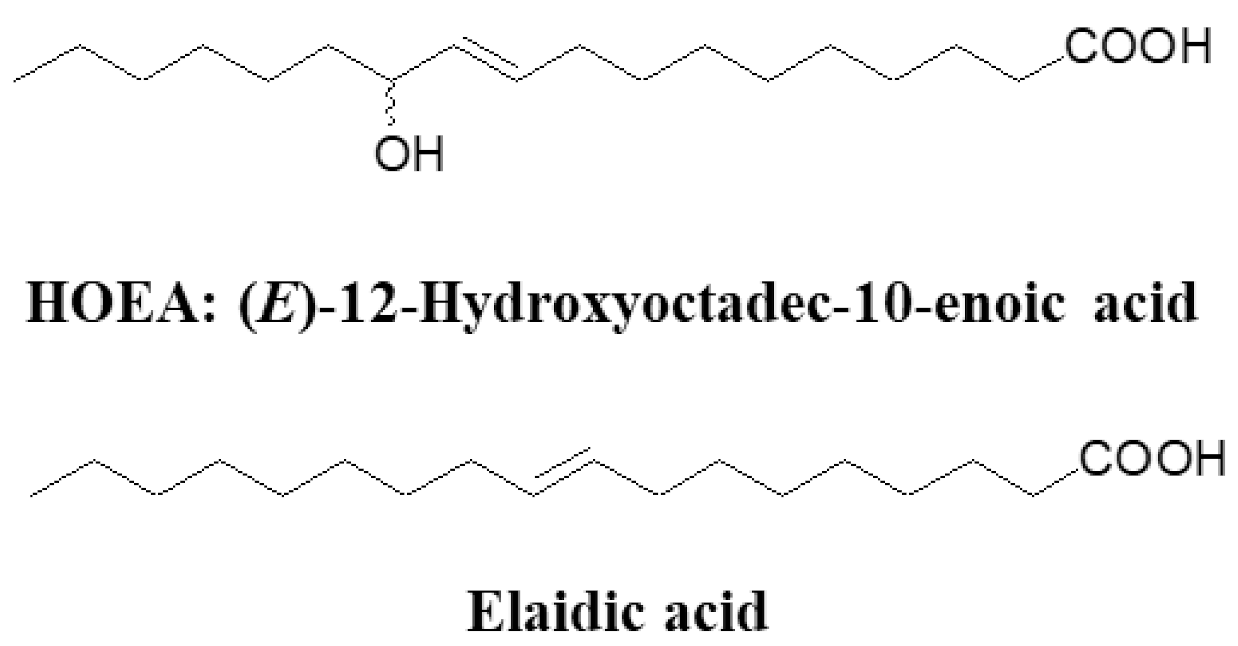
2.1. Chemical Structure of HOEA and Elaidic Acid
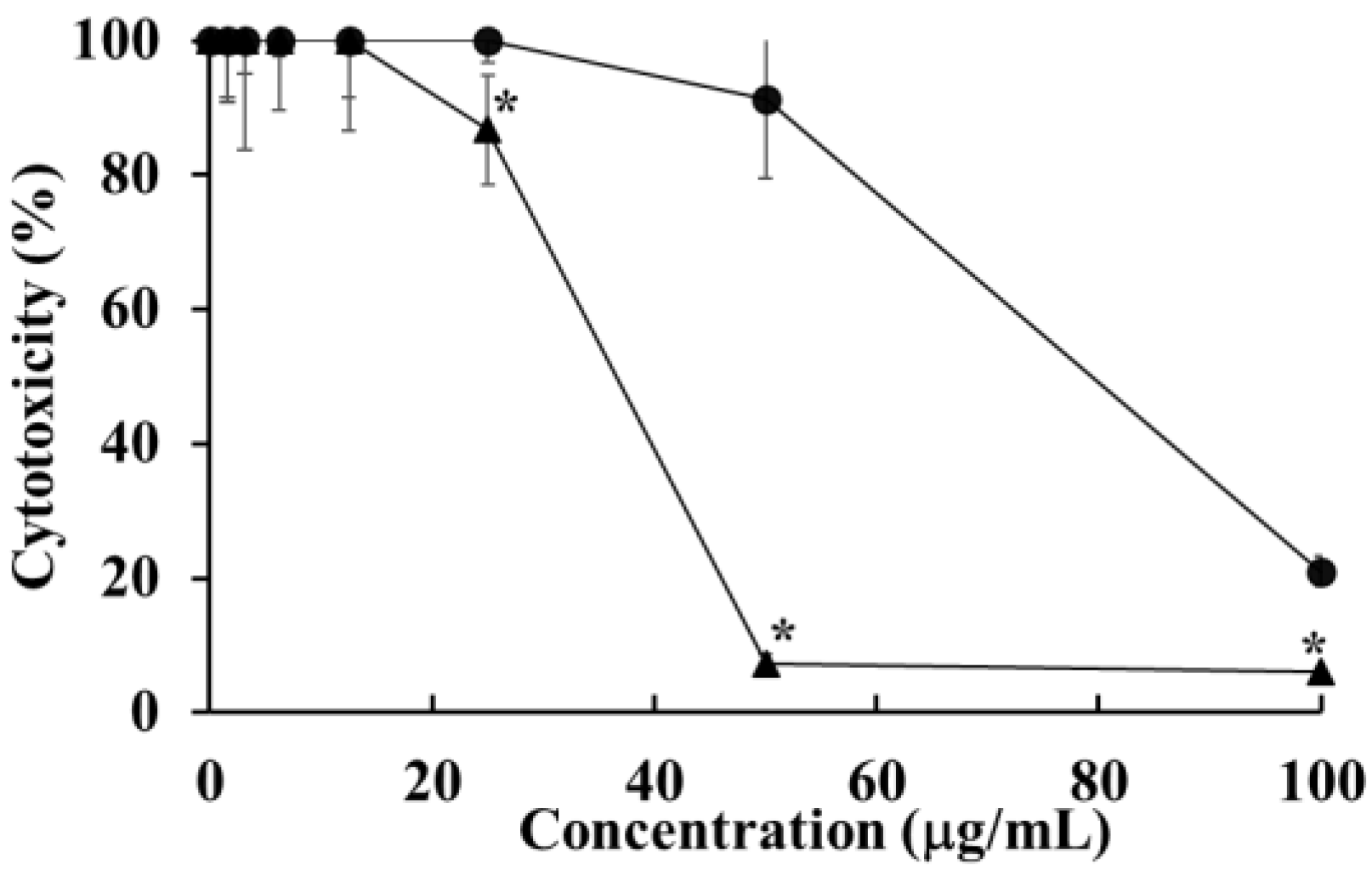
2.2. Cytotoxicity of HOEA and Elaidic Acid on U937 Cells
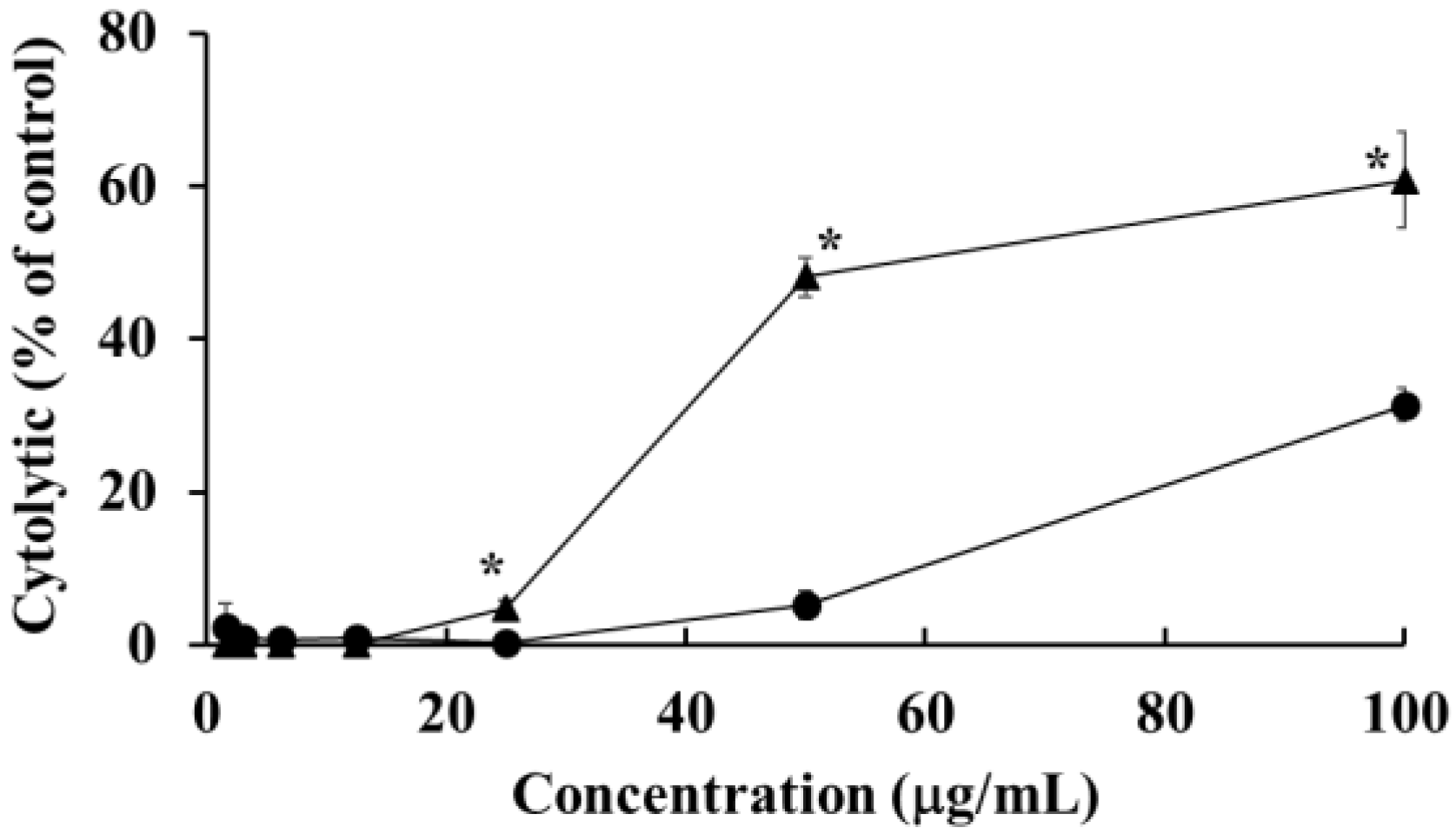
2.3. Cytolytic Effects of HOEA and Elaidic Acid on U937 Cells
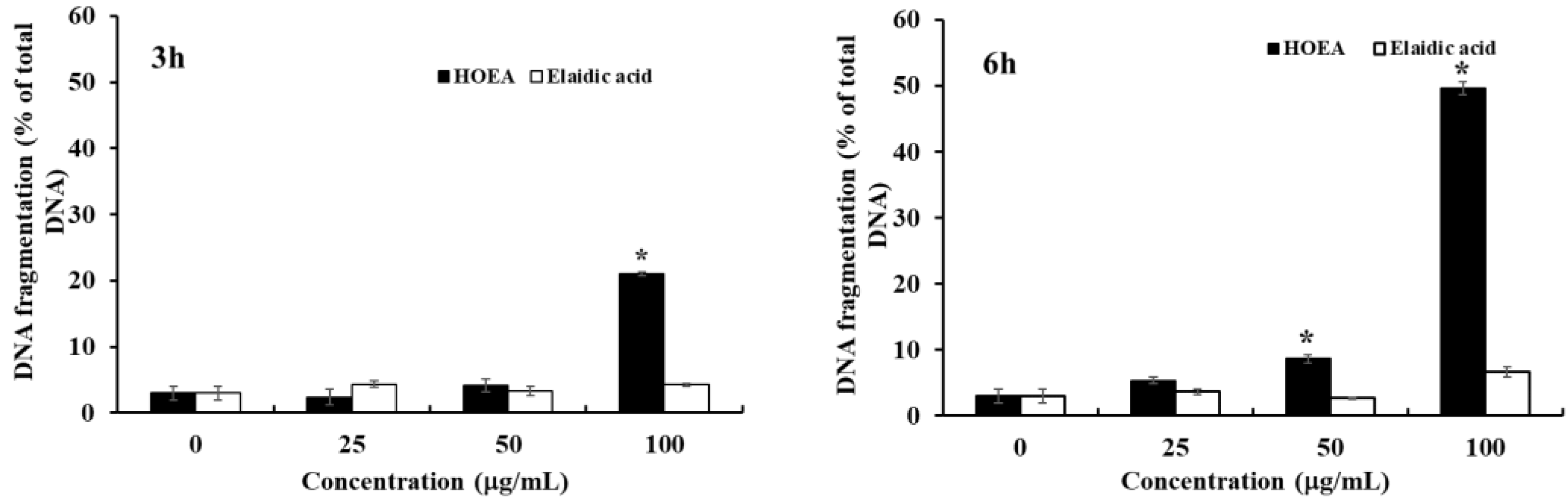
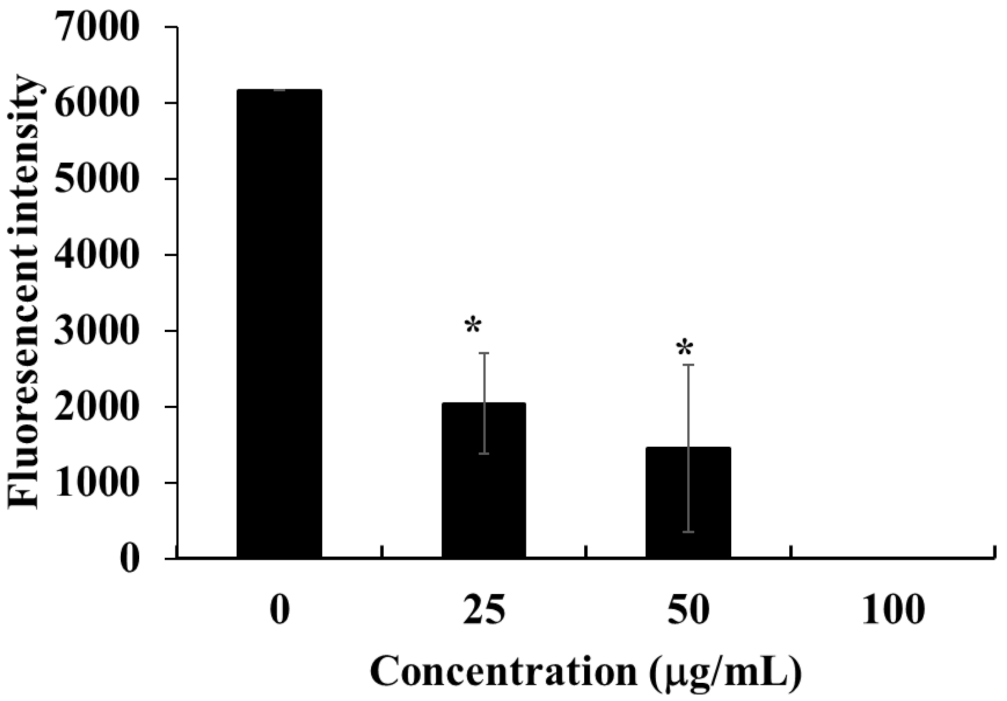
2.4. Detection of DNA Fragmentation in HOEA- and Elaidic Acid- Treated U937 Cells
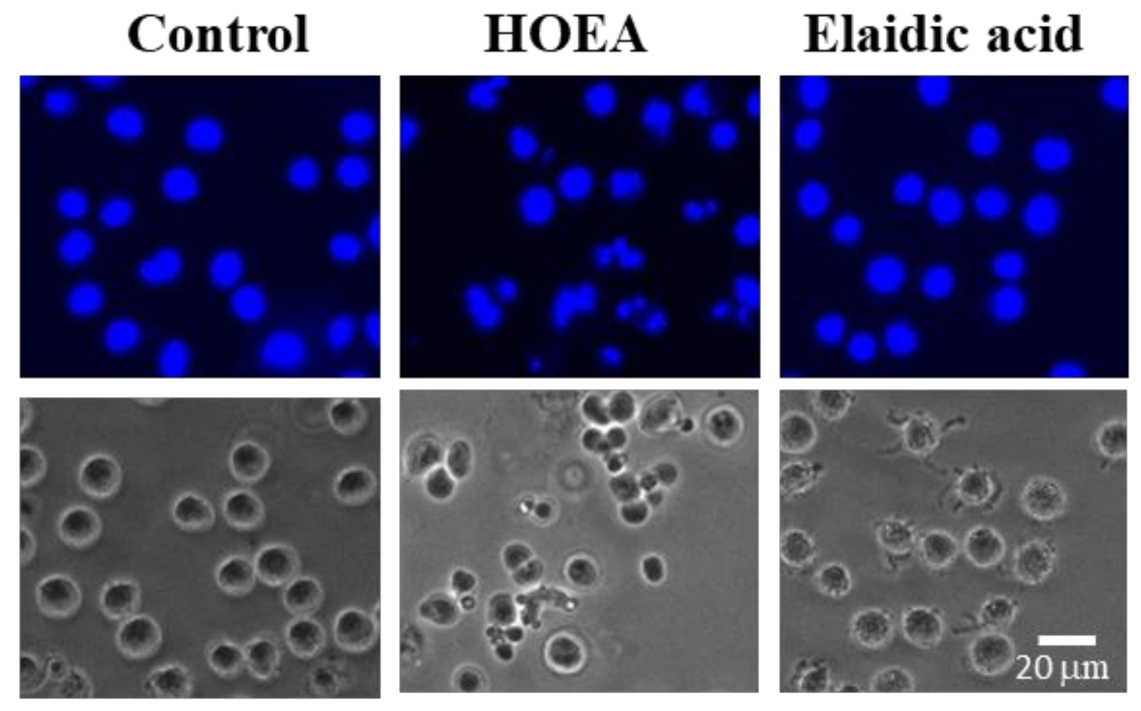
2.5. Nuclear Morophological Changes of U937 Cells Treated with HOEA or Elaidic Acid
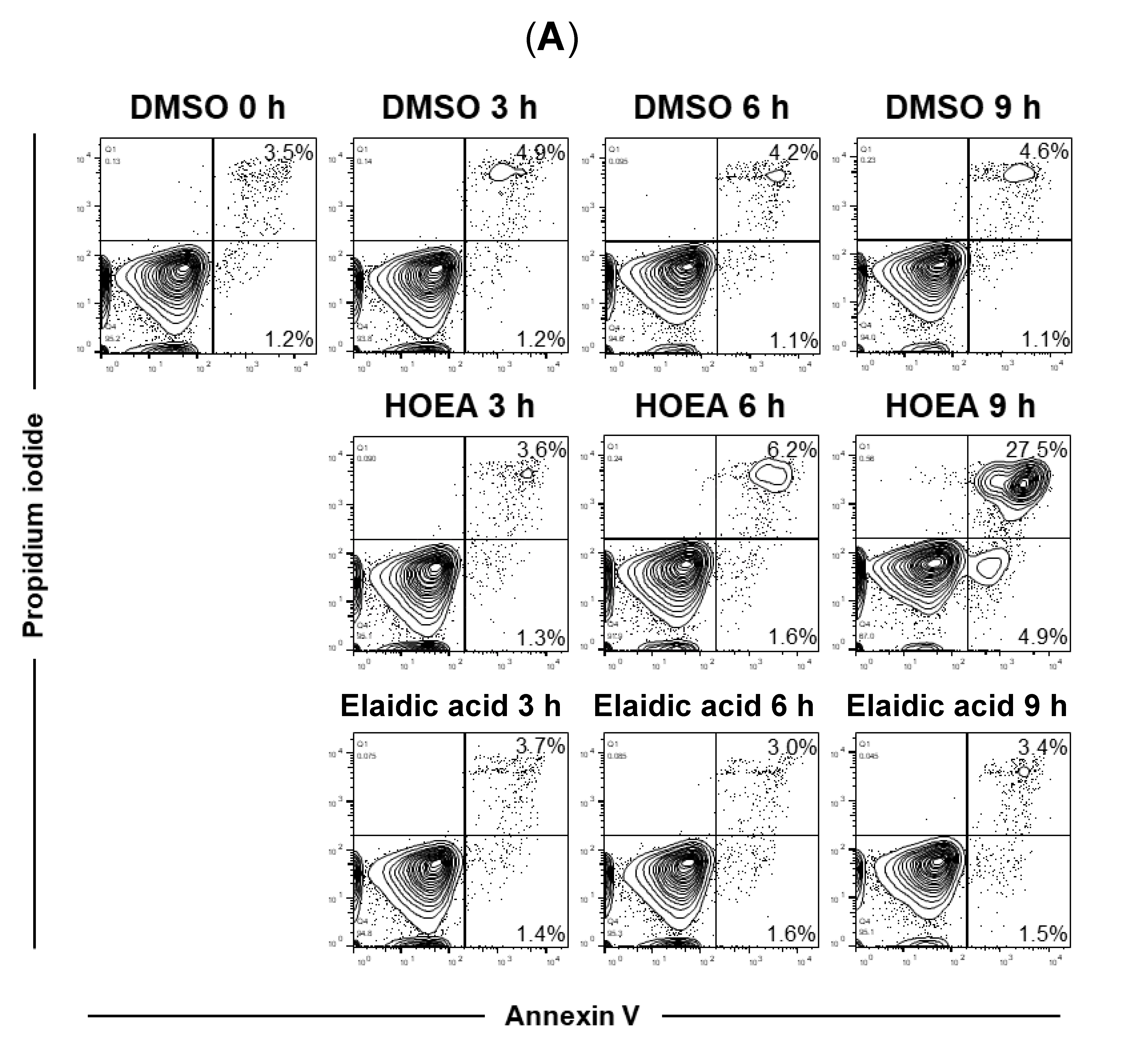
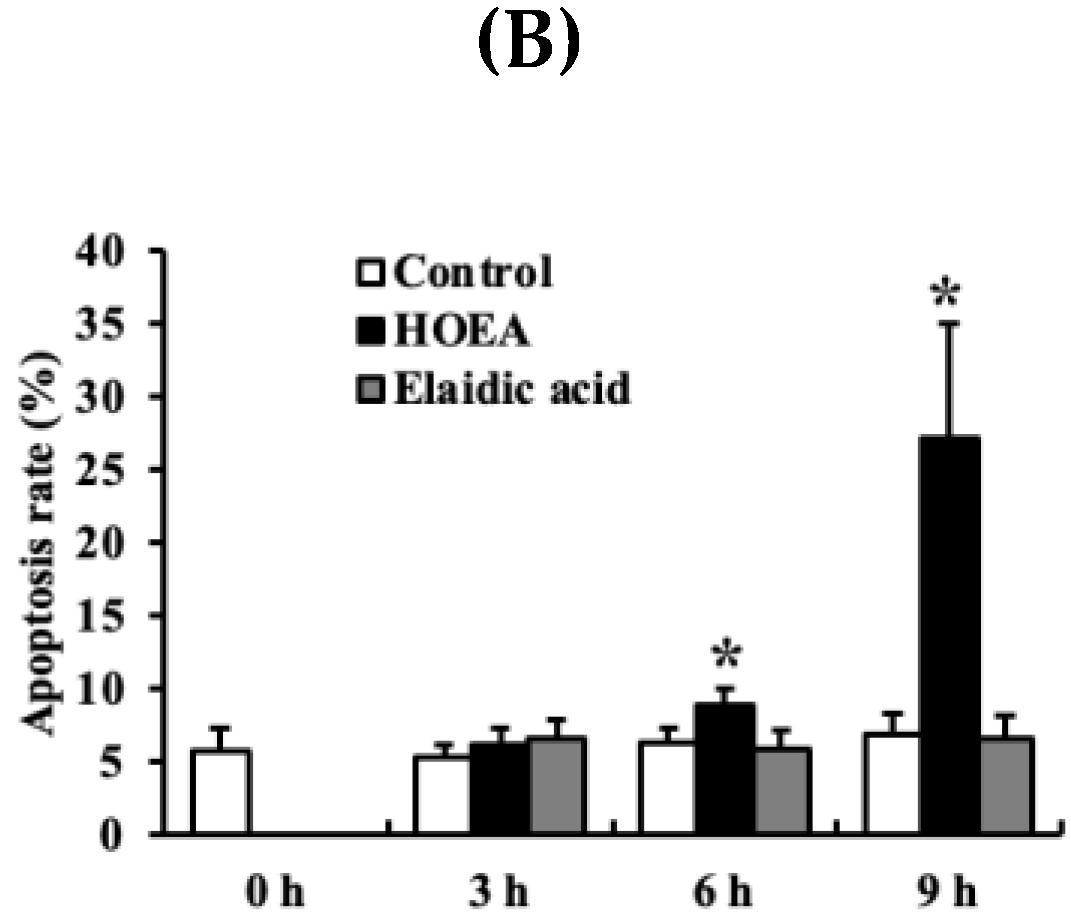
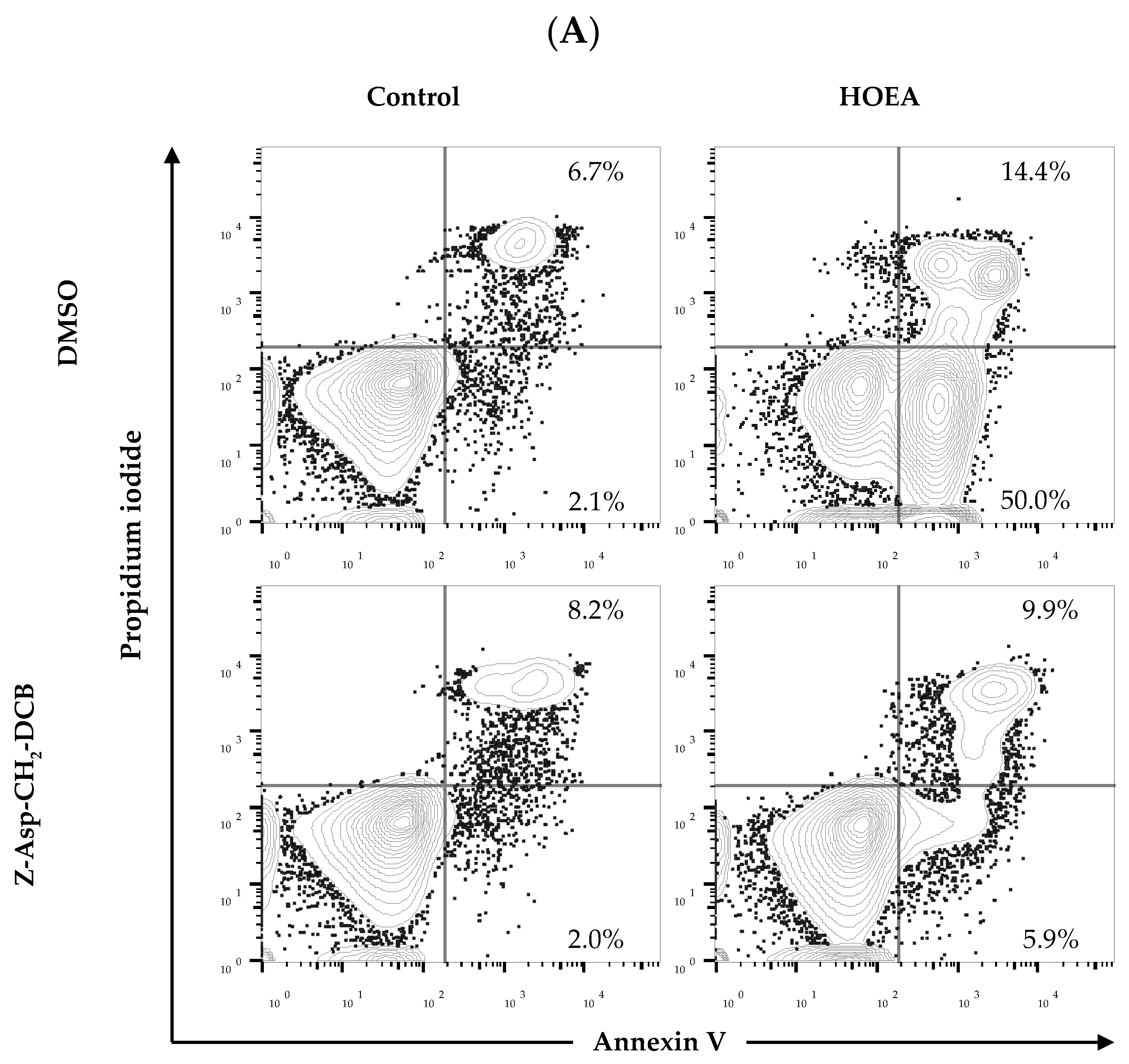
2.6. Flow Cytometry Analyses of HOEA- and Elaidic Acid-Treated U937 Cells
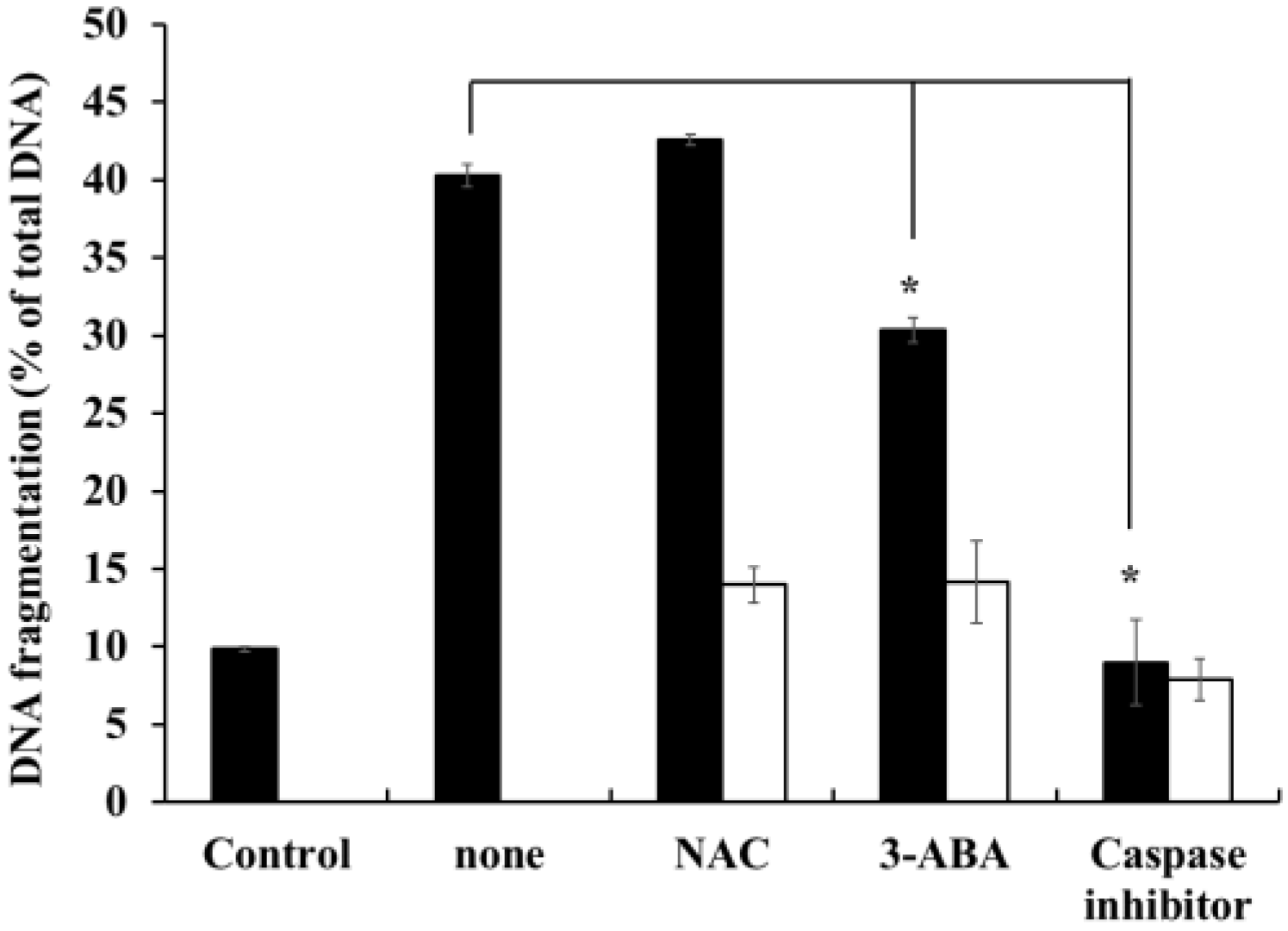
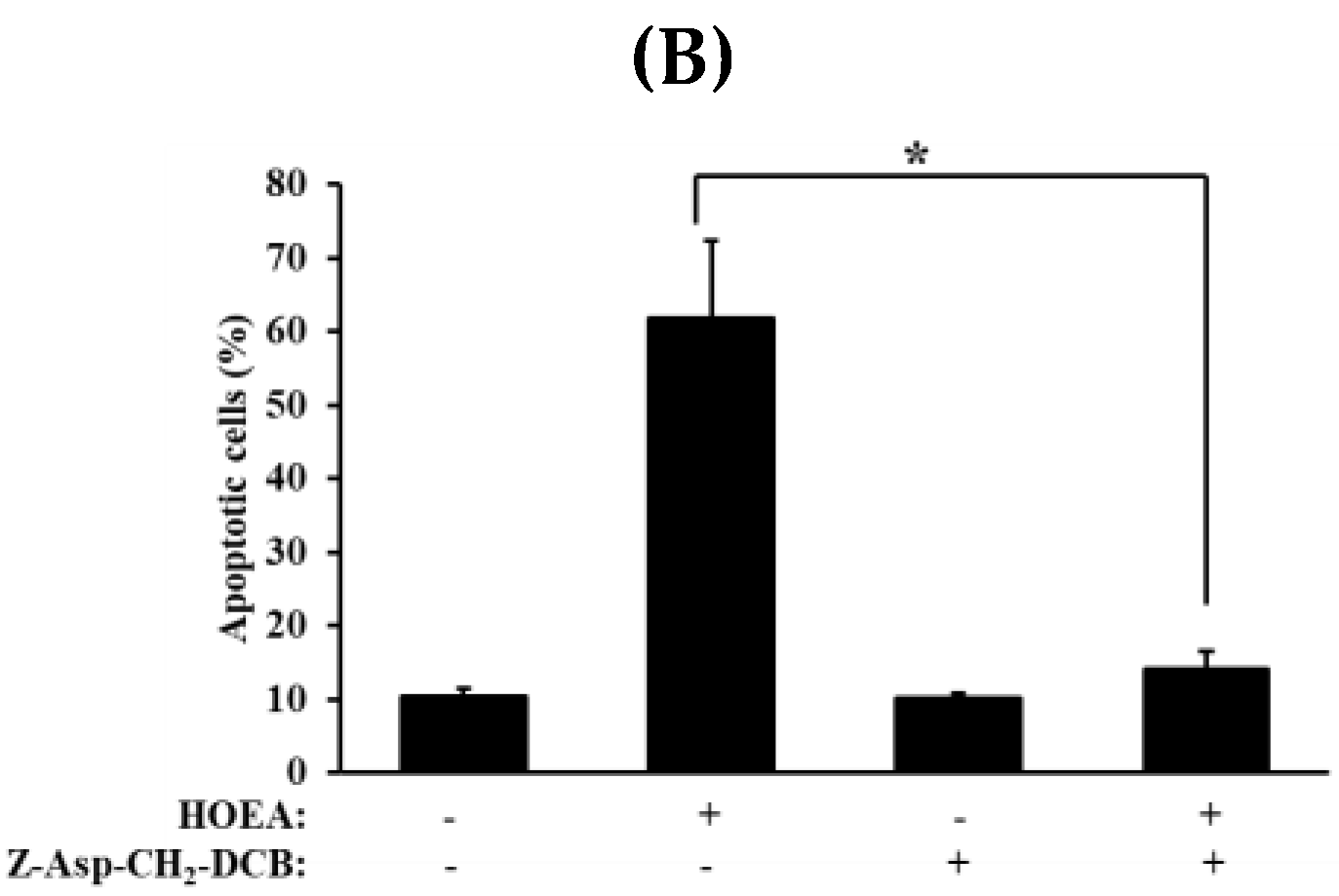
2.7. Effects of Caspase-Family Inhibitor (Z-Asp-CH2-DCB), N-acetyl-l-cysteine (NAC), and 3-aminobenzamide (3-ABA) on DNA-Fragmentation in HOEA-Treated U937 Cells
2.8. Effects of Caspase-Family Inhibitor (Z-Asp-CH2-DCB) on Annexin V+/PI− and Annexin V+/PI+ Cell Populations in the HOEA-Treated U937 Cells
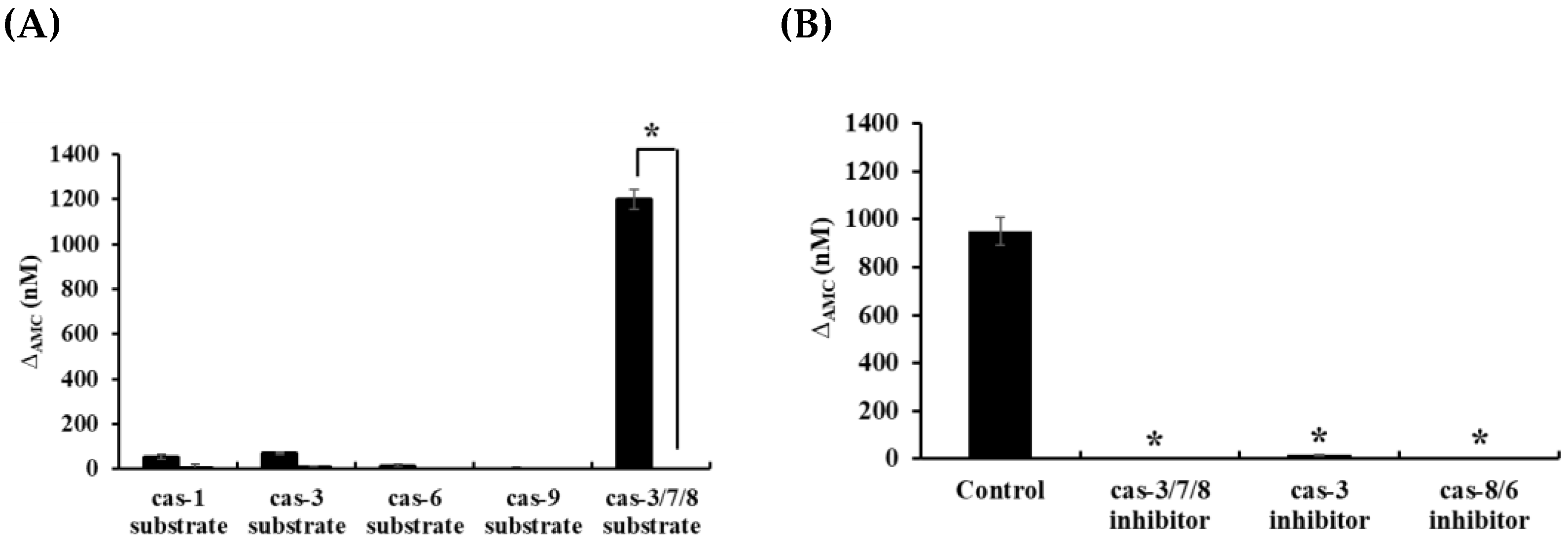
2.9. Analyses of Caspase Activities in HOEA-Treated U937 Cells
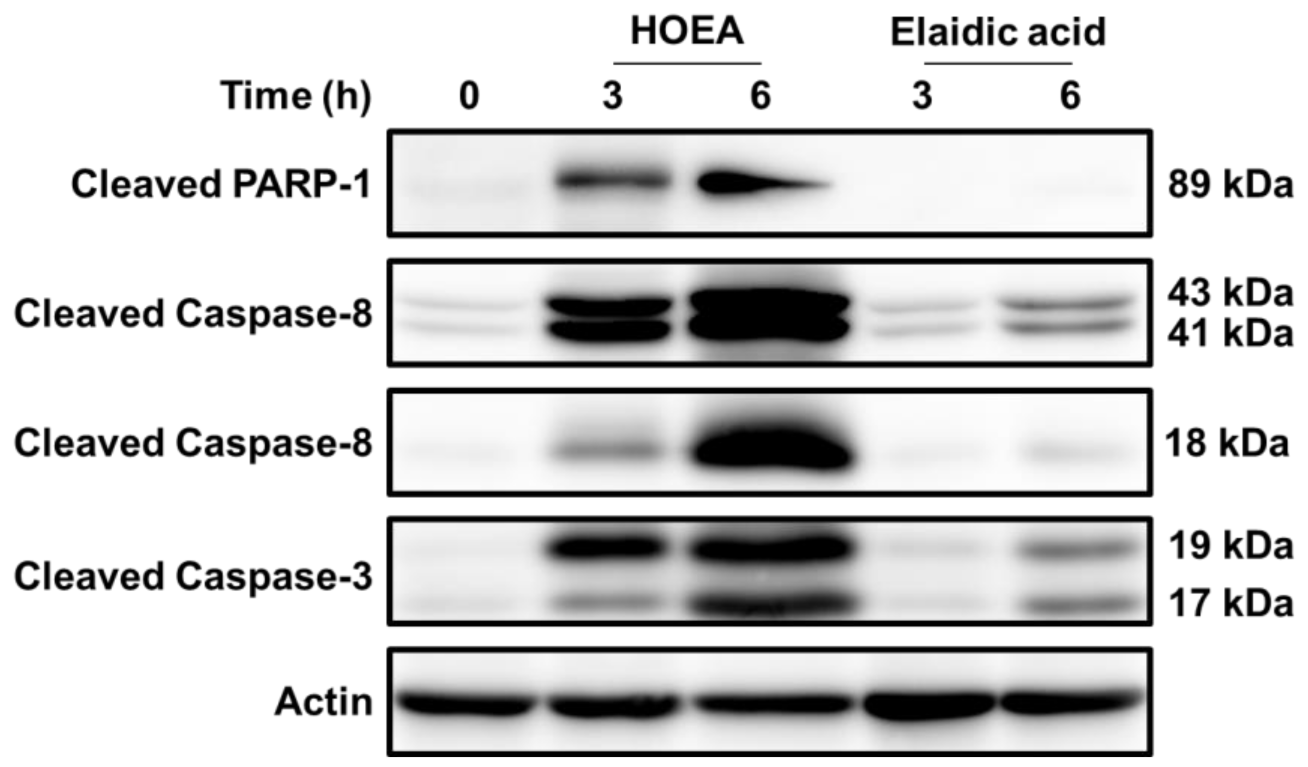
2.10. Effects of HOEA and Elaidic Acid on the Levels of Cleaved Forms of Caspase 3, Caspase 8, and PARP in U937 Cells
2.11. Effect of HOEA on Mitochondrial Membrane Potential in U937 Cells
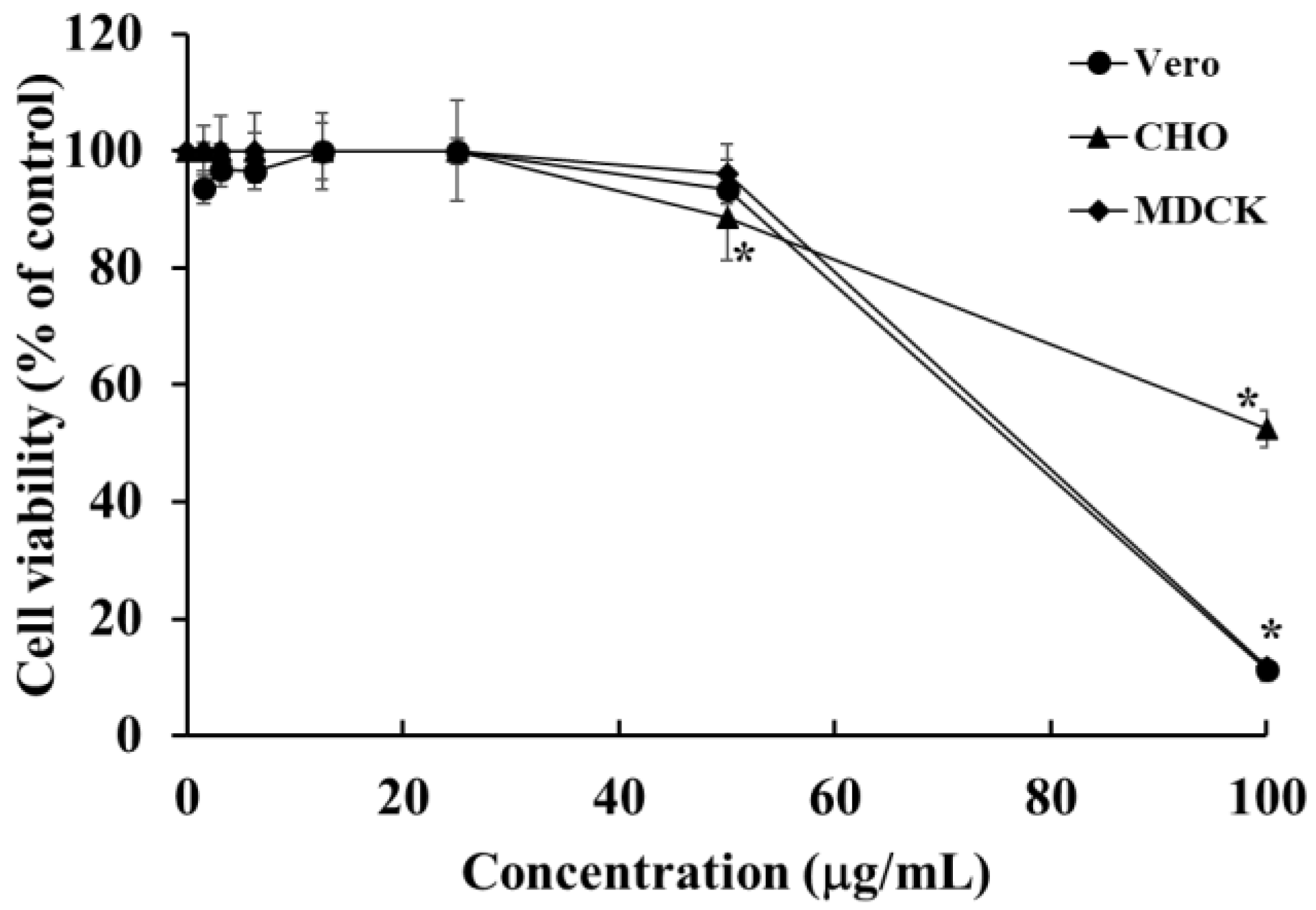
2.12. Cytotoxic Effects of HOEA onVero, CHO, and MDCK Cells
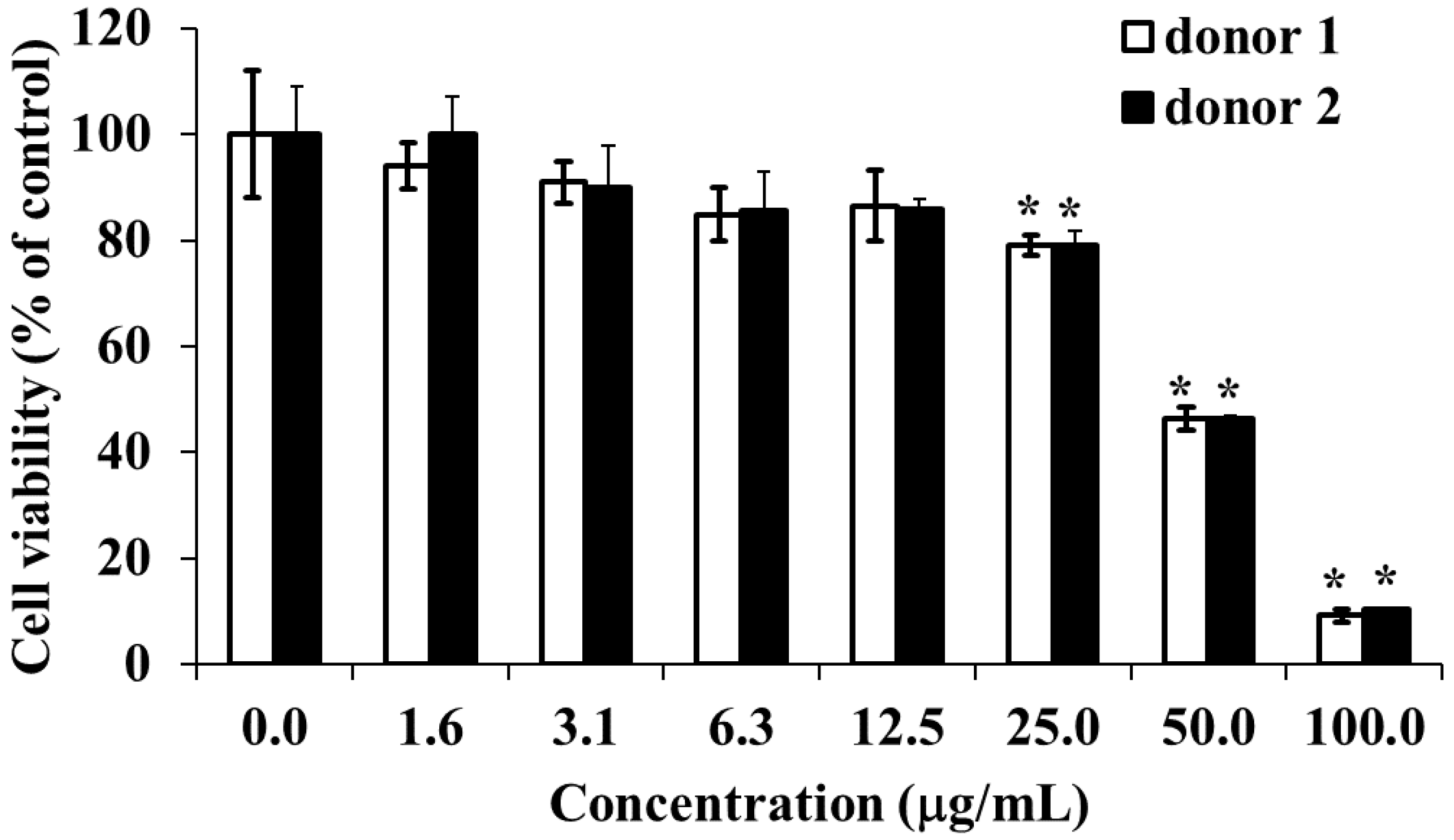
2.13. Cytotoxic Effect of HOEA on Human Peripheral Blood Mononuclear Cells (PBMCs)
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Cytotoxicity Assay
4.4. Cytolytic Assay
4.5. Fluorescence Observation of Nuclear Morphological Changes
4.6. Quantification of DNA Fragmentation
4.7. Measurement of the Effects of Caspase Family Inhibitor (Z-Asp-CH2-DCB), NAC and 3-ABA on HOEA-Induced DNA Fragmentation in U937 Cells
4.8. Caspase Activities
4.9. Flow Cytometry Analysis
4.10. Western Blotting
4.11. Detection of Mitochondrial Membrane Potential
4.12. Preparation of PBMCs and Cell Viability Assay
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hellio, C.; De La Broise, D.; Dufosse, L.; Le Gal, Y.; Bourgougnon, N. Hellio 2001 Anti-Bact.Pdf. Mar. Environ. Res. 2001, 52, 231–247. [Google Scholar] [CrossRef]
- Okai, Y.; Higashi-Okai, K.; Nakamura, S.I.; Yano, Y.; Otani, S. Suppressive effects of the extracts of Japanese edible seaweeds on mutagen-induced umu C gene expression in Salmonella typhimurium (TA 1535/pSK 1002) and tumor promotor-dependent ornithine decarboxylase induction in BALB/c 3T3 fibroblast cells. Cancer Lett. 1994. [Google Scholar] [CrossRef]
- Qi, H.; Zhang, Q.; Zhao, T.; Hu, R.; Zhang, K.; Li, Z. In vitro antioxidant activity of acetylated and benzoylated derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta). Bioorg. Med. Chem. Lett. 2006, 16, 2441–2445. [Google Scholar] [CrossRef]
- Okai, Y.; Higashi-Okai, K. Potent anti-inflammatory activity of pheophytin a derived from edible green alga, Enteromorpha prolifera (Sujiao-nori). Int. J. Immunopharmacol. 1997, 19, 355–358. [Google Scholar] [CrossRef]
- Nan, C.; Zhang, H.; Zhao, G. Allelopathic interactions between the macroalga Ulva pertusa and eight microalgal species. J. Sea Res. 2004, 52, 259–268. [Google Scholar] [CrossRef]
- Kakisawa, H.; Asari, F.; Kusumi, T.; Toma, T.; Sakurai, T.; Oohusa, T.; Hara, Y.; Chiharai, M. An allelopathic fatty acid from the brown alga Cladosiphon okamuranus. Phytochemistry 1988, 27, 731–735. [Google Scholar] [CrossRef]
- Alamsjah, M.A.; Hirao, S.; Ishibashi, F.; Fujita, Y. Isolation and structure determination of algicidal compounds from Ulva fasciata. Biosci. Biotechnol. Biochem. 2005, 69, 2186–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luyen, Q.H.; Cho, J.Y.; Choi, J.S.; Kang, J.Y.; Park, N.G.; Hong, Y.K. Isolation of algal spore lytic C17 fatty acid from the crustose coralline seaweed Lithophyllum yessoense. J. Appl. Phycol. 2009, 21, 423–427. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Y.; Tang, X. Identification of the toxic compounds produced by Sargassum thunbergii to red tide microalgae. Chin. J. Oceanol. Limnol. 2012, 30, 778–785. [Google Scholar] [CrossRef]
- Hirao, S.; Tara, K.; Kuwano, K.; Tanaka, J.; Ishibashi, F. Algicidal activity of glycerolipids from brown alga Ishige sinicola toward red tide microalgae. Biosci. Biotechnol. Biochem. 2012, 76, 372–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.Y.; Wang, H.; Guo, G.L.; Pu, Y.F.; Yan, B.L.; Wang, C.H. Isolation, purification, and identification of antialgal substances in green alga Ulva prolifera for antialgal activity against the common harmful red tide microalgae. Environ. Sci. Pollut. Res. 2016, 23, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Dong, S.; Guo, G.; Guo, L.; Pu, Y. Antialgal Activity of Glycoglycerolipids Derived from a Green Macroalgae Ulva prolifera on Six Species of Red Tide Microalgae. IOP Conf. Ser. Mater. Sci. Eng. 2019, 484. [Google Scholar] [CrossRef]
- Sun, Y.Y.; Meng, K.; Su, Z.X.; Guo, G.L.; Pu, Y.F.; Wang, C.H. Isolation and purification of antialgal compounds from the red alga Gracilaria lemaneiformis for activity against common harmful red tide microalgae. Environ. Sci. Pollut. Res. 2017, 24, 4964–4972. [Google Scholar] [CrossRef]
- Kim, J.Y.; Alamsjah, M.A.; Hamada, A.; Fujita, Y.; Ishibashi, F. Algicidal diterpenes from the brown alga Dictyota dichotoma. Biosci. Biotechnol. Biochem. 2006, 70, 2571–2574. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Xie, H.; Gong, Y.; Wang, Q.; Yang, Y. Secondary metabolites from the seaweed Gracilaria lemaneiformis and their allelopathic effects on Skeletonema costatum. Biochem. Syst. Ecol. 2011, 39, 397–400. [Google Scholar] [CrossRef]
- Ishibashi, F.; Sato, S.; Sakai, K.; Hirao, S.; Kuwano, K. Algicidal sesquiterpene hydroquinones from the brown alga dictyopteris undulata. Biosci. Biotechnol. Biochem. 2013, 77, 1120–1122. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.Y.; Zhou, W.J.; Wang, H.; Guo, G.L.; Su, Z.X.; Pu, Y.F. Antialgal compounds with antialgal activity against the common red tide microalgae from a green algae Ulva pertusa. Ecotoxicol. Environ. Saf. 2018, 157, 61–66. [Google Scholar] [CrossRef]
- Zha, S.; Kuwano, K.; Shibahara, T.; Ishibashi, F. Algicidal hydroxylated C18 unsaturated fatty acids from the red alga Tricleocarpa jejuensis: Identification, synthesis and biological activity. Fitoterapia 2020, 145, 104639. [Google Scholar] [CrossRef] [PubMed]
- Zha, S.; Liang, Y.; Oda, T.; Ishibashi, F. Bioactivities of algicidal C18 hydroxy unsaturated fatty acid isolated from the red alga Tricleocarpa jejuensis and its synthesized propargylic derivative. Algal Res. 2020, 52, 102097. [Google Scholar] [CrossRef]
- Ercolano, G.; De Cicco, P.; Ianaro, A. New drugs from the sea: Pro-apoptotic activity of sponges and algae derived compounds. Mar. Drugs 2019, 17, 31. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, D.W.; Thornberry, N.A. Caspases: Killer proteases. Trends Biochem. Sci. 1997, 22, 299–306. [Google Scholar] [CrossRef]
- Villa, P.; Kaufmann, S.H.; Earnshaw, W.C. Caspases and caspase inhibitors. Trends Biochem. Sci. 1997, 22, 388–393. [Google Scholar] [CrossRef]
- Okuma, E.; Saeki, K.; Shimura, M.; Ishizaka, Y.; Yasugi, E.; Yuo, A. Induction of apoptosis in human hematopoietic U937 cells by granulocyte-macrophage colony-stimulating factor: Possible existence of caspase 3-like pathway. Leukemia 2000, 14, 612–619. [Google Scholar] [CrossRef] [Green Version]
- Cohen, G.M. Caspases: The Executioners of Apoptosis—A review. Biochem. J. 1997, 326, 1–116. [Google Scholar] [CrossRef]
- Lazebnik, Y.A.; Kaufmann, S.H.; Desnoyers, S.; Poirier, G.G.; Earnshaw, W.C. Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 1994, 371, 346–347. [Google Scholar] [CrossRef] [PubMed]
- Boulares, A.H.; Yakovlev, A.G.; Ivanova, V.; Stoica, B.A.; Wang, G.; Iyer, S.; Smulson, M. Role of poly(ADP-ribose) polymerase (PARP) cleavage in apoptosis. Caspase 3-resistant PARP mutant increases rates of apoptosis in transfected cells. J. Biol. Chem. 1999, 274, 22932–22940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Alamsjah, M.A.; Ishibe, K.; Kim, D.; Yamaguchi, K.; Ishibashi, F.; Fujita, Y.; Oda, T. Selective toxic effects of polyunsaturated fatty acids derived from Ulva fasciata on red tide phyotoplankter species. Biosci. Biotechnol. Biochem. 2007, 265–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, R.; Noguchi, R.; Ota, T.; Abe, M.; Miyashita, K.; Kawada, T. Cytotoxic effect of conjugated trienoic fatty acids on mouse tumor and human monocytic leukemia cells. Lipids 2001, 36, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, J.; Steinhart, H. Analysis, occurrence, and physiological properties of trans fatty acids (TFA) with particular emphasis on conjugated linoleic acid isomers (CLA)—A review. Eur. J. Lipid Sci. Technol. 1998, 100, 190–210. [Google Scholar] [CrossRef]
- Kohno, H.; Suzuki, R.; Noguchi, R.; Hosokawa, M.; Miyashita, K.; Tanaka, T. Dietary conjugated linolenic acid inhibits azoxymethane-induced colonic aberrant crypt foci in rats. Jpn. J. Cancer Res. 2002, 93, 133–142. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Santalova, E.A.; Gorshkova, I.A.; Dmitrenok, A.S.; Guzii, A.G.; Gorbach, V.I.; Svetashev, V.I.; Stonik, V.A. A new cytotoxic fatty acid (5Z,9Z)-22-methyl-5,9-tetracosadienoic acid and the sterols from the far eastern sponge Geodinella robusta. Lipids 2002, 37, 75–80. [Google Scholar] [CrossRef]
- Schleich, K.; Buchbinder, J.H.; Pietkiewicz, S.; Kähne, T.; Warnken, U.; Öztürk, S.; Schnölzer, M.; Naumann, M.; Krammer, P.H.; Lavrik, I.N. Molecular architecture of the DED chains at the DISC: Regulation of procaspase-8 activation by short DED proteins c-FLIP and procaspase-8 prodomain. Cell Death Differ. 2016, 23, 681–694. [Google Scholar] [CrossRef]
- Mashima, T.; Naito, M.; Kataoka, S.; Kawai, H.; Tsuruo, T. Aspartate-based inhibitor of interleukin-1β-converting enzyme prevents antitumor agent-induced apoptosis in human myeloid leukemia U937 cells+. Biochem. Biophys. Res. Commun. 1995, 209, 907–915. [Google Scholar] [CrossRef]
- Komatsu, N.; Oda, T.; Muramatsu, T. Involvement of both caspase-like proteases and serine proteases in apoptotic cell death induced by ricin, modeccin, diphtheria toxin, and Pseudomonas toxin. J. Biochem. 1998, 124, 1038–1044. [Google Scholar] [CrossRef]
- Cossarizza, A.; Franceschi, C.; Monti, D.; Salvioli, S.; Bellesia, E.; Rivabene, R.; Biondo, L.; Rainaldi, G.; Tinari, A.; Malorni, W. Protective effect of N-acetylcysteine in tumor necrosis factor-α-induced apoptosis in U937 cells: The role of mitochondria. Exp. Cell Res. 1995, 220, 232–240. [Google Scholar] [CrossRef]
- Kawahara, A.; Enari, M.; Talanian, R.V.; Wong, W.W.; Nagata, S. Fas-induced DNA fragmentation and proteolysis of nuclear proteins. Genes Cells 1998. [Google Scholar] [CrossRef]
- Walisser, J.A.; Thies, R.L. Poly(ADP-ribose) polymerase inhibition in oxidant-stressed endothelial cells prevents oncosis and permits caspase activation and apoptosis. Exp. Cell Res. 1999, 251, 401–413. [Google Scholar] [CrossRef]
- Tanaka, Y.; Yoshihara, K.; Tohno, Y.; Kojima, K.; Kameoka, M.; Kamiya, T. Inhibition and down-regulation of poly(ADP-ribose) polymerase results in a marked resistance of HL-60 cells to various apoptosis-inducers. Cell. Mol. Biol. 1995, 41, 771–781. [Google Scholar] [PubMed]
- Tummers, B.; Green, D.R. Caspase-8: Regulating life and death. Immunol. Rev. 2017, 277, 76–89. [Google Scholar] [CrossRef] [Green Version]
- De, A.K.; Muthiyan, R.; Mondal, S.; Mahanta, N.; Bhattacharya, D.; Ponraj, P.; Muniswamy, K.; Kundu, A.; Kundu, M.S.; Sunder, J.; et al. A natural quinazoline derivative from marine sponge hyrtios erectus induces apoptosis of breast cancer cells via ROS production and intrinsic or extrinsic apoptosis pathways. Mar. Drugs 2019, 17, 658. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Yan, C.; Schor, N.F. Apoptosis in the absence of caspase 3. Oncogene 2001, 20, 6570–6578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasula, S.M.; Fernandes-Alnemri, T.; Zangrilli, J.; Robertson, N.; Armstrong, R.C.; Wang, L.; Trapani, J.A.; Tomaselli, K.J.; Litwack, G.; Alnemri, E.S. The Ced-3/interleukin 1β converting enzyme-like homolog Mch6 and the lamin-cleaving enzyme Mch2α are substrates for the apoptotic mediator CPP32. J. Biol. Chem. 1996, 271, 27099–27106. [Google Scholar] [CrossRef] [Green Version]
- Varfolomeev, E.E.; Schuchmann, M.; Luria, V.; Chiannilkulchai, N.; Beckmann, J.S.; Mett, I.L.; Rebrikov, D.; Brodianski, V.M.; Kemper, O.C.; Kollet, O.; et al. Targeted disruption of the mouse Caspase 8 gene ablates cell death induction by the TNF receptors, Fas/Apo1, and DR3 and is lethal prenatally. Immunity 1998, 9, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Muzio, M.; Chinnaiyan, A.M.; Kischkel, F.C.; O’Rourke, K.; Shevchenko, A.; Ni, J.; Scaffidi, C.; Bretz, J.D.; Zhang, M.; Gentz, R.; et al. FLICE, a novel FADD-homologous ICE/CED-3-like protease, is recruited to the CD95 (Fas/APO-1) death-inducing signaling complex. Cell 1996, 85, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Enari, M.; Sakahira, H.; Yokoyama, H.; Okawa, K.; Iwamatsu, A.; Nagata, S. A caspase-activated DNase that degrades DNA during apoptosis, and its inhibitor ICAD. Nature 1998, 391, 43–50. [Google Scholar] [CrossRef]
- Aslan, B.; Ozpolat, B.; Sood, A.K.; Lopez-Berestein, G. Nanotechnology in cancer therapy. J. Drug Target. 2013, 21, 904–913. [Google Scholar] [CrossRef] [Green Version]
- Bildstein, L.; Dubernet, C.; Couvreur, P. Prodrug-based intracellular delivery of anticancer agents. Adv. Drug Deliv. Rev. 2011, 63, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Tsukigawa, K.; Fang, J. A Retrospective 30 Years after Discovery of the Enhanced Permeability and Retention Effect of Solid Tumors: Next-Generation Chemotherapeutics and Photodynamic Therapy—Problems, Solutions, and Prospects. Microcirculation 2016, 23, 173–182. [Google Scholar] [CrossRef]
- Maeda, H.; Nakamura, H.; Fang, J. The EPR effect for macromolecular drug delivery to solid tumors: Improvement of tumor uptake, lowering of systemic toxicity, and distinct tumor imaging in vivo. Adv. Drug Deliv. Rev. 2013, 65, 71–79. [Google Scholar] [CrossRef]
- Islam, W.; Matsumoto, Y.; Fang, J.; Harada, A.; Niidome, T.; Ono, K.; Tsutsuki, H.; Sawa, T.; Imamura, T.; Sakurai, K.; et al. Polymer-conjugated glucosamine complexed with boric acid shows tumor-selective accumulation and simultaneous inhibition of glycolysis. Biomaterials 2021, 269, 120631. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Islam, W. 3-overcoming barriers for tumor-targeted drug delivery: The power of macromolecular anticancer drugs with the EPR effect and the modulation of vascular physiology. In Polymer-Protein Conjugates; Elsevier: Amsterdam, The Netherlands, 2020; pp. 41–58. [Google Scholar]
- Desai, N.; Trieu, V.; Yao, Z.; Louie, L.; Ci, S.; Yang, A.; Tao, C.; De, T.; Beals, B.; Dykes, D.; et al. Increased antitumor activity, intratumor paclitaxel concentrations, and endothelial cell transport of cremophor-free, albumin-bound paclitaxel, ABI-007, compared with cremophor-based paclitaxel. Clin. Cancer Res. 2006, 12, 1317–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Luo, C.; Cui, W.; Sun, J.; He, Z. Chemotherapy agent-unsaturated fatty acid prodrugs and prodrug-nanoplatforms for cancer chemotherapy. J. Control Release 2017, 264, 145–159. [Google Scholar] [CrossRef]
- Bougnoux, P.; Hajjaji, N.; Maheo, K.; Couet, C.; Chevalier, S. Fatty acids and breast cancer: Sensitization to treatments and prevention of metastatic re-growth. Prog. Lipid Res. 2010, 49, 76–86. [Google Scholar] [CrossRef]
- Hajjaji, N.; Bougnoux, P. Selective sensitization of tumors to chemotherapy by marine-derived lipids: A review. Cancer Treat. Rev. 2013, 39, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Jóźwiak, M.; Filipowska, A.; Fiorino, F.; Struga, M. Anticancer activities of fatty acids and their heterocyclic derivatives. Eur. J. Pharmacol. 2020, 871, 172937. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, P.Y.; Hettiarachchi, S.D.; Zhou, Y.; Ouhtit, A.; Seven, E.S.; Oztan, C.Y.; Celik, E.; Leblanc, R.M. Nanoparticle-mediated targeted drug delivery for breast cancer treatment. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 419–433. [Google Scholar] [CrossRef]
- Komatsu, N.; Nakagawa, M.; Oda, T.; Muramatsu, T. Depletion of intracellular NAD+ and ATP levels during ricin-induced apoptosis through the specific ribosomal inactivation results in the cytolysis of U937 cells. J. Biochem. 2000. [Google Scholar] [CrossRef]















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zha, S.; Ueno, M.; Liang, Y.; Okada, S.; Oda, T.; Ishibashi, F. Induction of Apoptotic Cell Death in Human Leukemia U937 Cells by C18 Hydroxy Unsaturated Fatty Acid Isolated from Red Alga Tricleocarpa jejuensis. Mar. Drugs 2021, 19, 138. https://0-doi-org.brum.beds.ac.uk/10.3390/md19030138
Zha S, Ueno M, Liang Y, Okada S, Oda T, Ishibashi F. Induction of Apoptotic Cell Death in Human Leukemia U937 Cells by C18 Hydroxy Unsaturated Fatty Acid Isolated from Red Alga Tricleocarpa jejuensis. Marine Drugs. 2021; 19(3):138. https://0-doi-org.brum.beds.ac.uk/10.3390/md19030138
Chicago/Turabian StyleZha, Shijiao, Mikinori Ueno, Yan Liang, Seiji Okada, Tatsuya Oda, and Fumito Ishibashi. 2021. "Induction of Apoptotic Cell Death in Human Leukemia U937 Cells by C18 Hydroxy Unsaturated Fatty Acid Isolated from Red Alga Tricleocarpa jejuensis" Marine Drugs 19, no. 3: 138. https://0-doi-org.brum.beds.ac.uk/10.3390/md19030138