
Seasonal Variation of the Proximate Composition, Mineral Content, Fatty Acid Profiles and Other Phytochemical Constituents of Selected Brown Macroalgae

 ,
,  ,
,  , ,
, , 
Abstract
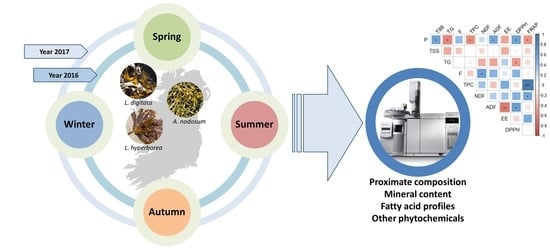
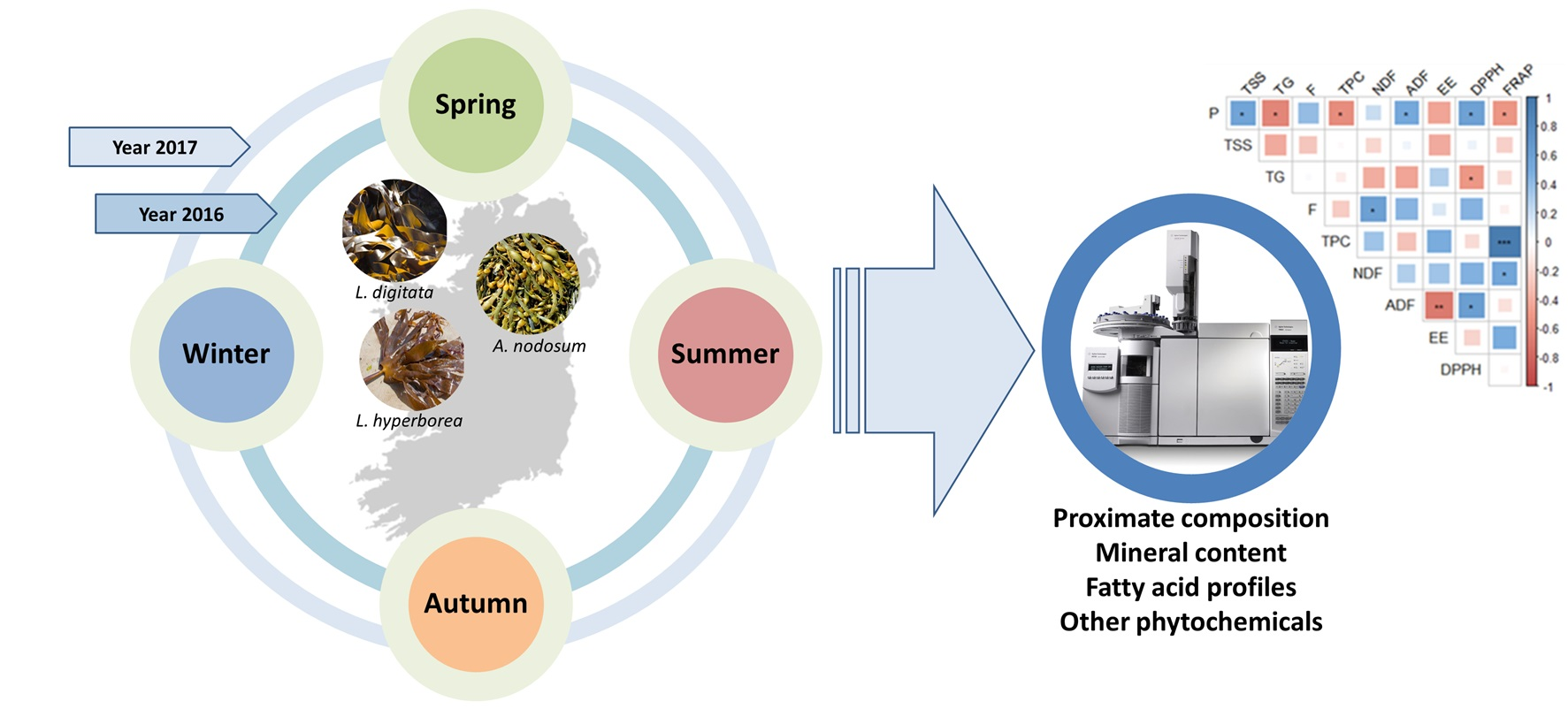
:
1. Introduction
2. Results and Discussion
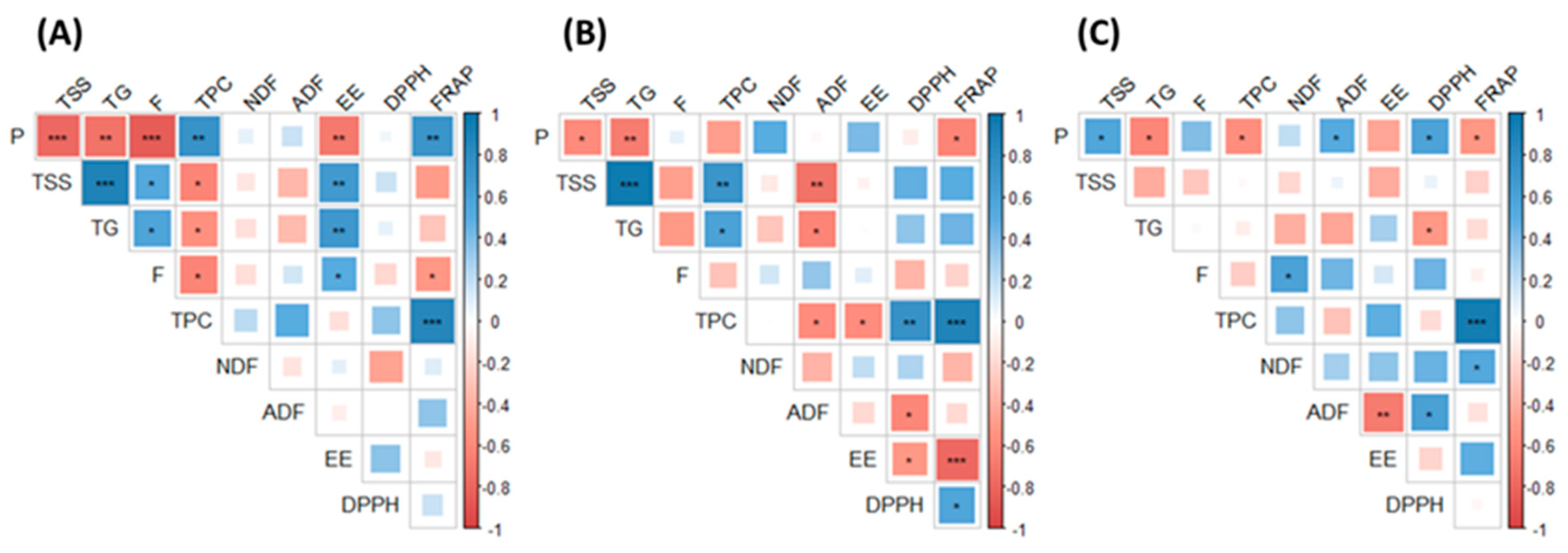
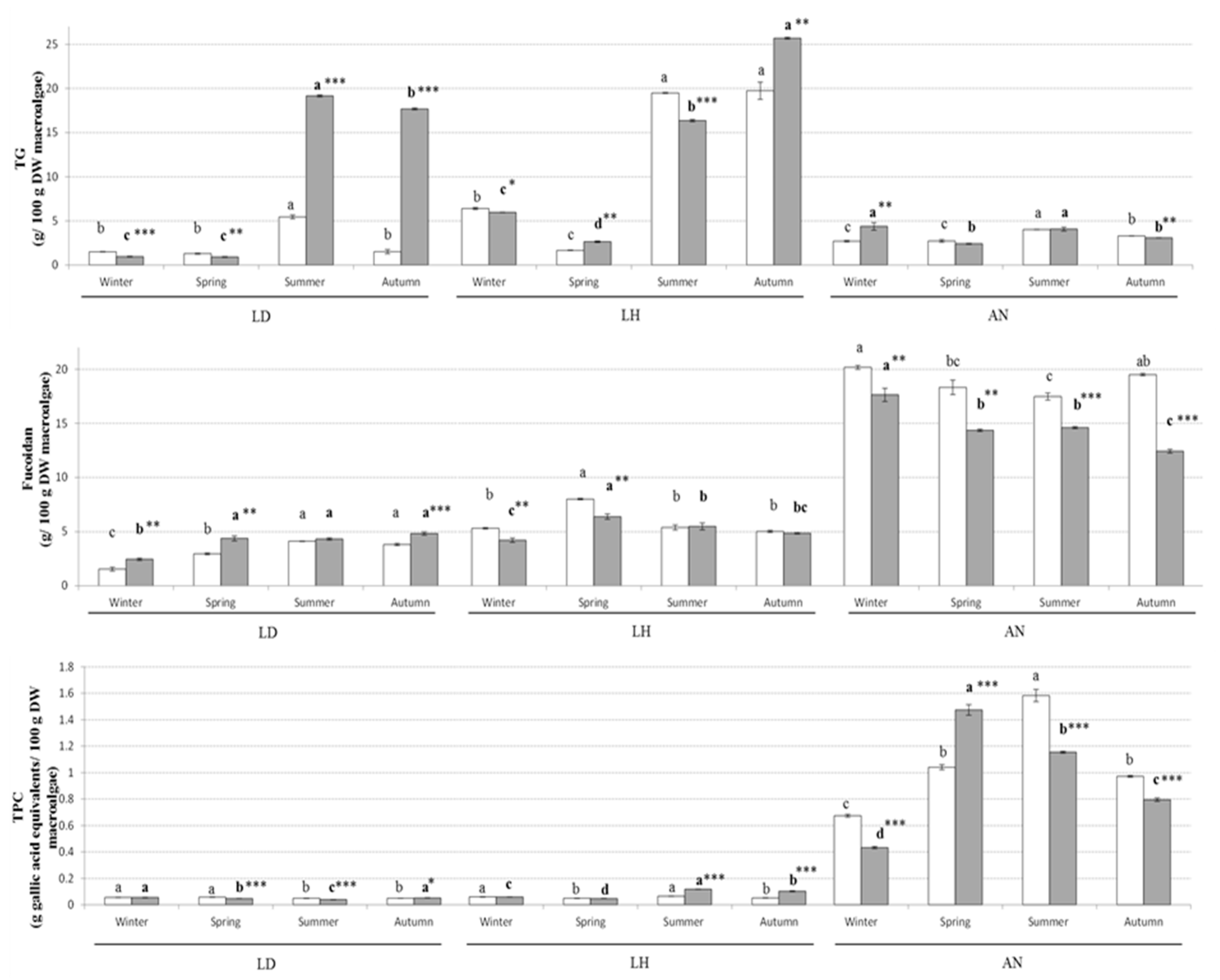
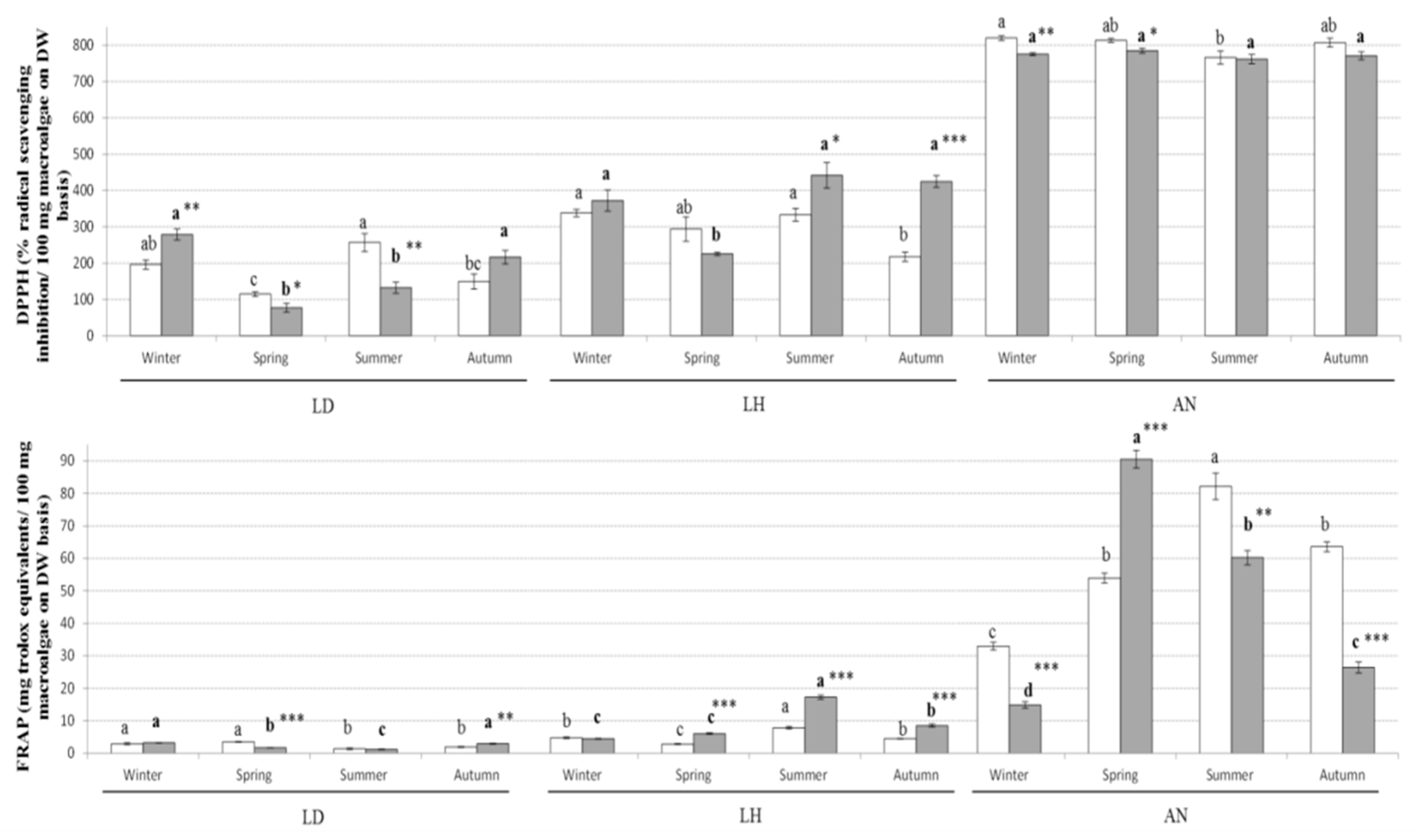
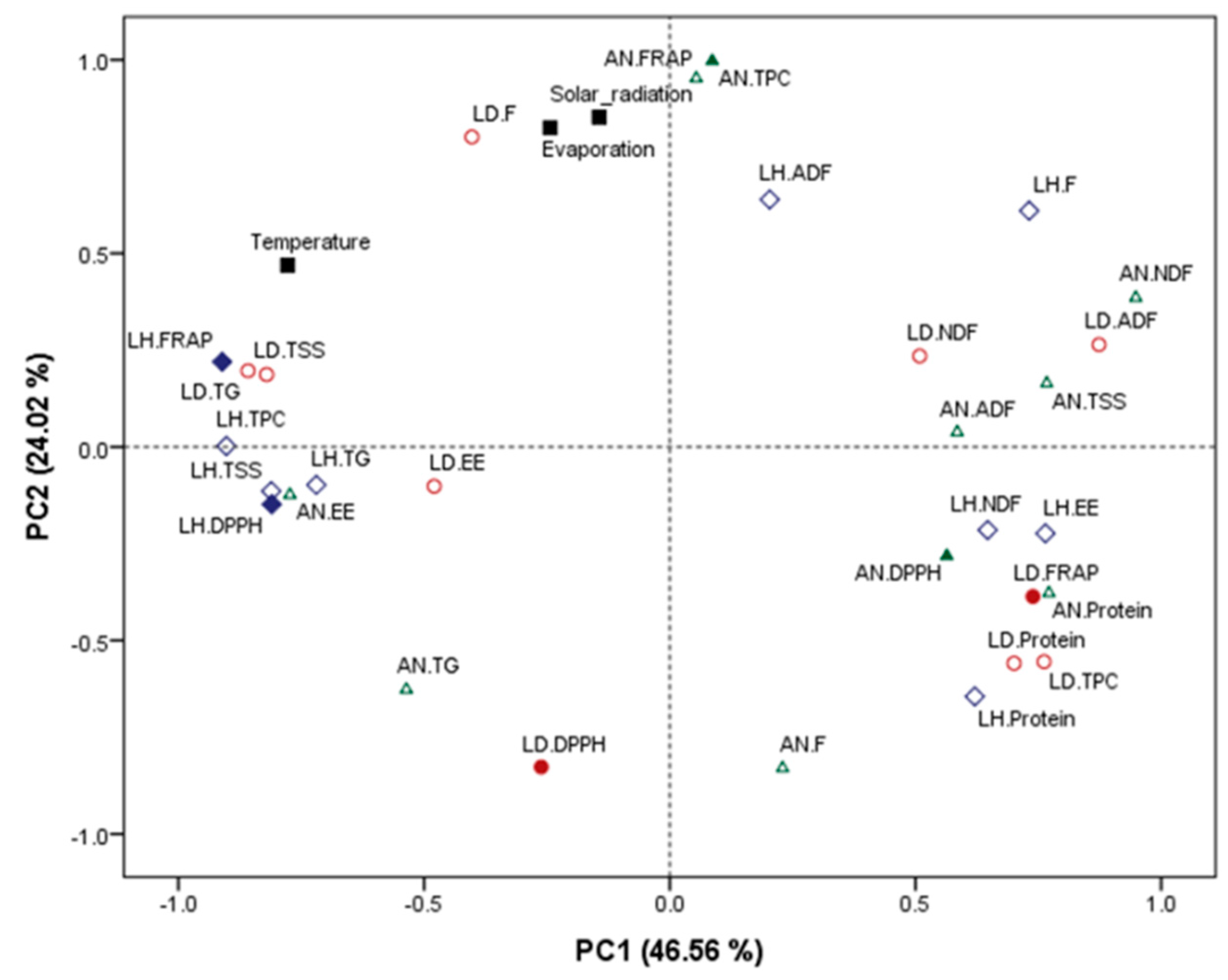
2.1. Proximate Composition, Phytochemical Concentration and Antioxidant Capacity
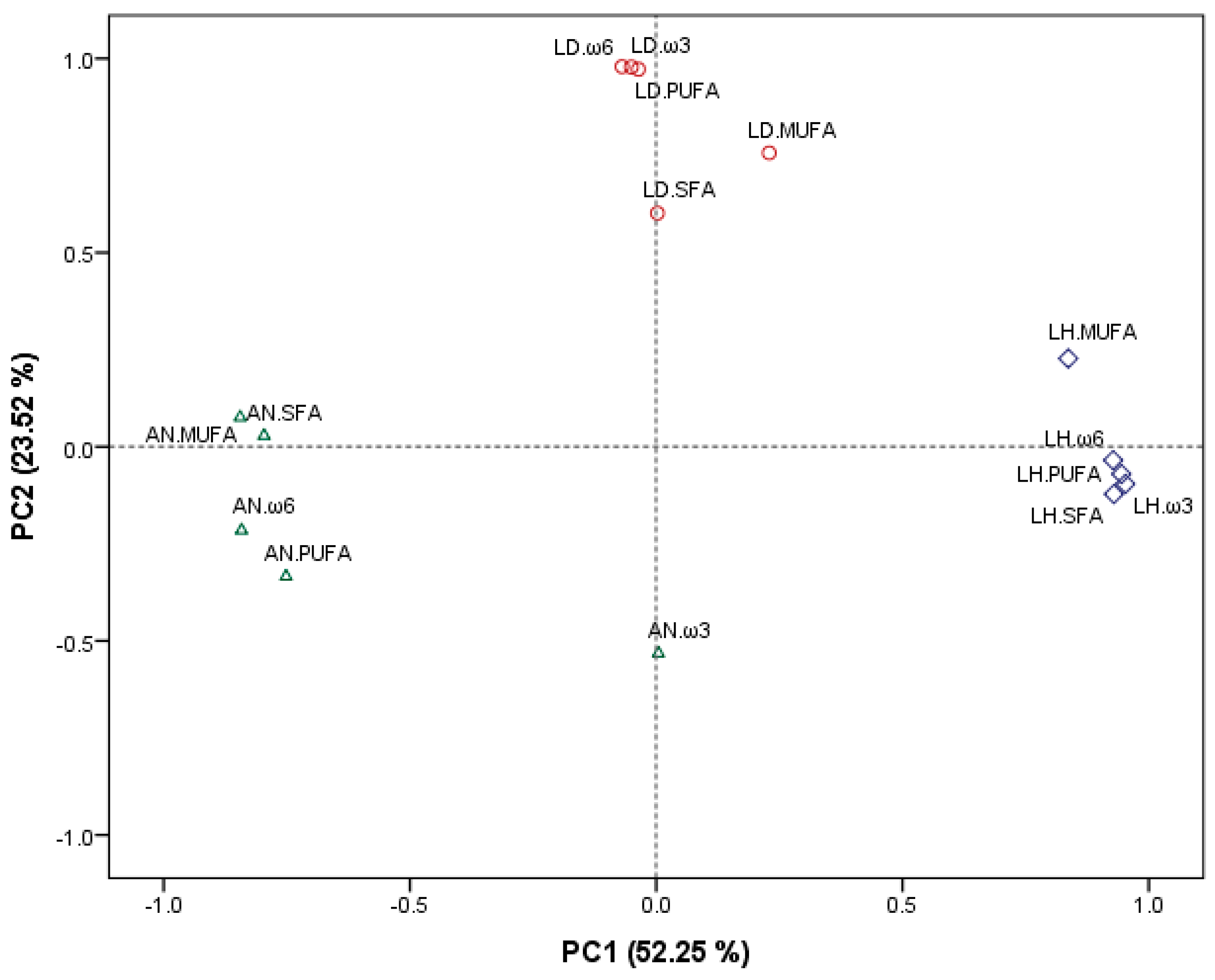
2.2. Fatty Acid Profiling
2.3. Essential and Toxic Trace Metals
3. Materials and Methods
3.1. Macroalgal Biomass, Collection and Preparation
3.2. Chemical Analyses
3.3. Fatty Acid Profiling
3.4. Essential and Toxic Metal Profiling
3.5. Climatological Data
3.6. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chojnacka, K.; Saeid, A.; Witkowska, Z.; Tuhy, L. Biologically Active Compounds in Seaweed Extracts—The Prospects for the Application; The Open Conference Proceedings Journal; Bentham Open: Bussum, The Netherlands, 2012. [Google Scholar]
- Dominguez, H.; Loret, E.P. Ulva lactuca, a source of troubles and potential riches. Mar. Drugs 2019, 17, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, K.G.; Fitzgerald, G.F.; Stanton, C.; Ross, R.P. Looking beyond the terrestrial: The potential of seaweed derived bioactives to treat non-communicable diseases. Mar. Drugs 2016, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roleda, M.Y.; Marfaing, H.; Desnica, N.; Jónsdóttir, R.; Skjermo, J.; Rebours, C.; Nitschke, U. Variations in polyphenol and heavy metal contents of wild-harvested and cultivated seaweed bulk biomass: Health risk assessment and implication for food applications. Food Control 2019, 95, 121–134. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Hayes, M. Red and green macroalgae for fish and animal feed and human functional food development. Food Rev. Int. 2016, 32, 15–45. [Google Scholar] [CrossRef]
- Déléris, P.; Nazih, H.; Bard, J.-M. Seaweeds in human health. In Seaweed in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2016; pp. 319–367. [Google Scholar]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Vaquero, M.; Rajauria, G.; O’Doherty, J.; Sweeney, T. Polysaccharides from macroalgae: Recent advances, innovative technologies and challenges in extraction and purification. Food Res. Int. 2017, 99, 1011–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapiye, C.; Vahmani, P.; Mlambo, V.; Muchenje, V.; Dzama, K.; Hoffman, L.; Dugan, M. The trans-octadecenoic fatty acid profile of beef: Implications for global food and nutrition security. Food Res. Int. 2015, 76, 992–1000. [Google Scholar] [CrossRef]
- Pelliccia, F.; Marazzi, G.; Greco, C.; Franzoni, F.; Speziale, G.; Gaudio, C. Current evidence and future perspectives on n−3 pufas. Int. J. Cardiol. 2013, 170, S3–S7. [Google Scholar] [CrossRef]
- Committee, E.S. Statement on the benefits of fish/seafood consumption compared to the risks of methylmercury in fish/seafood. Efsa J. 2015, 13, 3982. [Google Scholar] [CrossRef] [Green Version]
- CONTAM Panel. Scientific opinion on arsenic in food. Efsa J. 2009, 7, 1351. [Google Scholar] [CrossRef]
- CONTAM Panel. Scientific opinion on lead in food. Efsa J. 2010, 8, 1570. [Google Scholar]
- EC 488/2014. Commission Regulation (EU) no 488/2014 of 12 May 2014 Amending Regulation (EC) no 1881/2006 as Regards Maximum Levels of Cadmium in Foodstuffs; European Union: Brussels, Belgium, 2014. [Google Scholar]
- EC 1881/2006. Commission Regulation (EC) no 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs; European Union: Brussels, Belgium, 2006. [Google Scholar]
- Shaltout, N.; Shams El Din, N. Investigation of the fatty acid profile in some macroalgae in relation to the environmental conditions for biodiesel production. Am. J. Environ. Sci. 2015, 11, 402–419. [Google Scholar] [CrossRef]
- Galloway, A.W.E.; Britton-Simmons, K.H.; Duggins, D.O.; Gabrielson, P.W.; Brett, M.T. Fatty acid signatures differentiate marine macrophytes at ordinal and family ranks1. J. Phycol. 2012, 48, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from buarcos bay in central west coast of portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Biancarosa, I.; Belghit, I.; Bruckner, C.; Liland, N.; Waagbø, R.; Amlund, H.; Heesch, S.; Lock, E. Chemical characterization of 21 species of marine macroalgae common in norwegian waters: Benefits of and limitations to their potential use in food and feed. J. Sci. Food Agric. 2018, 98, 2035–2042. [Google Scholar] [CrossRef] [Green Version]
- Usov, A.; Smirnova, G.; Klochkova, N. Algae polysaccharides. 55. Polysaccharide composition of some brown Kamchatka algae. Bioorganicheskaia Khimiia 2001, 27, 444–448. [Google Scholar] [PubMed]
- Skriptsova, A.; Shevchenko, N.; Tarbeeva, D.; Zvyagintseva, T. Comparative study of polysaccharides from reproductive and sterile tissues of five brown seaweeds. Mar. Biotechnol. 2011, 14, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine macroalgae as sources of protein and bioactive compounds in feed for monogastric animals. J. Sci. Food Agric. 2019, 99, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikker, P.; Stokvis, L.; van Krimpen, M.M.; van Wikselaar, P.G.; Cone, J.W. Evaluation of seaweeds from marine waters in northwestern europe for application in animal nutrition. Anim. Feed Sci. Technol. 2020, 263, 114460. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Giger-Reverdin, S.; Lessire, M.; Lebas, F.; Ankers, P. Seaweeds for livestock diets: A review. Anim. Feed Sci. Technol. 2016, 212, 1–17. [Google Scholar] [CrossRef]
- Jensen, R.B.; Brøkner, C.; Bach Knudsen, K.E.; Tauson, A.-H. A comparative study of the apparent total tract digestibility of carbohydrates in icelandic and danish warmblood horses fed two different haylages and a concentrate consisting of sugar beet pulp and black oats. Arch. Anim. Nutr. 2010, 64, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future protein supply and demand: Strategies and factors influencing a sustainable equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Fletcher, H.R.; Biller, P.; Ross, A.B.; Adams, J.M.M. The seasonal variation of fucoidan within three species of brown macroalgae. Algal Res. 2017, 22, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Parys, S.; Kehraus, S.; Pete, R.; Küpper, F.C.; Glombitza, K.-W.; König, G.M. Seasonal variation of polyphenolics in Ascophyllum nodosum (phaeophyceae). Eur. J. Phycol. 2009, 44, 331–338. [Google Scholar] [CrossRef]
- Häder, D.-P.; Kumar, H.; Smith, R.; Worrest, R. Effects on aquatic ecosystems. J. Photochem. Photobiol. B Biol. 1998, 46, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Henry, B.E.; Van Alstyne, K.L. Effects of uv radiation on growth and phlorotannins in Fucus gardneri (phaeophyceae) juveniles and embryos 1. J. Phycol. 2004, 40, 527–533. [Google Scholar] [CrossRef]
- Pavia, H.; Toth, G.B. Influence of light and nitrogen on the phlorotannin content of the brown seaweeds Ascophyllum nodosum and Fucus vesiculosus. Hydrobiologia 2000, 440, 299–305. [Google Scholar] [CrossRef]
- Barbosa, M.; Fernandes, F.; Pereira, D.M.; Azevedo, I.C.; Sousa-Pinto, I.; Andrade, P.B.; Valentão, P. Fatty acid patterns of the kelps Saccharina latissima, Saccorhiza polyschides and Laminaria ochroleuca: Influence of changing environmental conditions. Arab. J. Chem. 2020, 13, 45–58. [Google Scholar] [CrossRef]
- da Costa, E.; Domingues, P.; Melo, T.; Coelho, E.; Pereira, R.; Calado, R.; Abreu, M.H.; Domingues, M.R. Lipidomic signatures reveal seasonal shifts on the relative abundance of high-valued lipids from the brown algae Fucus vesiculosus. Mar. Drugs 2019, 17, 335. [Google Scholar] [CrossRef] [Green Version]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Fatty acid contents and profiles of 16 macroalgae collected from the irish coast at two seasons. J. Appl. Phycol. 2014, 26, 451–463. [Google Scholar] [CrossRef]
- Susanto, E.; Fahmi, A.S.; Abe, M.; Hosokawa, M.; Miyashita, K. Lipids, fatty acids, and fucoxanthin content from temperate and tropical brown seaweeds. Aquat. Procedia 2016, 7, 66–75. [Google Scholar] [CrossRef]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Colombo, M.L.; Rise, P.; Giavarini, F.; De Angelis, L.; Galli, C.; Bolis, C. Marine macroalgae as sources of polyunsaturated fatty acids. Plant Foods Hum. Nutr. 2006, 61, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Blouin, N.; Calder, B.L.; Perkins, B.; Brawley, S.H. Sensory and fatty acid analyses of two Atlantic species of porphyra (rhodophyta). J. Appl. Phycol. 2006, 18, 79. [Google Scholar] [CrossRef]
- Drenjančević, I.; Kralik, G.; Kralik, Z.; Mihalj, M.; Stupin, A.; Novak, S.; Grčević, M. The effect of dietary intake of omega-3 polyunsaturated fatty acids on cardiovascular health: Revealing potentials of functional food. In Superfood and Functional Food-The Development of Superfoods and Their Roles as Medicine; Intech: Long Beach, CA, USA, 2017; pp. 207–232. [Google Scholar]
- für die Nährstoffzufuhr, A.R. Referenzwerte für Die Nährstoffzufuhr; Spektrum Akademischer: Heidelberg, Germany, 2000. [Google Scholar]
- Erkkilä, A.T.; Lehto, S.; Pyörälä, K.; Uusitupa, M.I. N−3 fatty acids and 5-y risks of death and cardiovascular disease events in patients with coronary artery disease. Am. J. Clin. Nutr. 2003, 78, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.B.; Willett, W.C. Optimal diets for prevention of coronary heart disease. JAMA 2002, 288, 2569–2578. [Google Scholar] [CrossRef]
- Nestel, P.; Shige, H.; Pomeroy, S.; Cehun, M.; Abbey, M.; Raederstorff, D. The n−3 fatty acids eicosapentaenoic acid and docosahexaenoic acid increase systemic arterial compliance in humans. Am. J. Clin. Nutr. 2002, 76, 326–330. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.V.; Tocher, D.R. Biosynthesis of polyunsaturated fatty acids in aquatic ecosystems: General pathways and new directions. In Lipids in Aquatic Ecosystems; Springer: New York City, NY, USA, 2009; pp. 211–236. [Google Scholar]
- Gladyshev, M.; Arts, M.; Sushchik, N.i. Preliminary estimates of the export of omega-3 highly unsaturated fatty acids (epa+ dha) from aquatic to terrestrial ecosystems. In Lipids in Aquatic Ecosystems; Springer: New York City, NY, USA, 2009; pp. 179–210. [Google Scholar]
- Khaled, A.; Hessein, A.; Abdel-Halim, A.M.; Morsy, F.M. Distribution of heavy metals in seaweeds collected along marsa-matrouh beaches, egyptian mediterranean sea. Egypt. J. Aquat. Res. 2014, 40, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Mæhre, H.K.; Malde, M.K.; Eilertsen, K.-E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef]
- Gall, E.A.; Küpper, F.C.; Kloareg, B. A survey of iodine content in laminaria digitata. Bot. Mar. 2004, 47, 30–37. [Google Scholar] [CrossRef]
- Küpper, F.C.; Carpenter, L.J.; McFiggans, G.B.; Palmer, C.J.; Waite, T.J.; Boneberg, E.-M.; Woitsch, S.; Weiller, M.; Abela, R.; Grolimund, D. Iodide accumulation provides kelp with an inorganic antioxidant impacting atmospheric chemistry. Proc. Natl. Acad. Sci. USA 2008, 105, 6954–6958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almela, C.; Algora, S.; Benito, V.; Clemente, M.J.; Devesa, V.; Suner, M.A.; Velez, D.; Montoro, R. Heavy metal, total arsenic, and inorganic arsenic contents of algae food products. J. Agric. Food Chem. 2002, 50, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on Undesirable Substances in Animal Feed—Council Statement; European Union: Brussels, Belgium, 2002.
- Hanaoka, K.i.; Yosida, K.; Tamano, M.; Kuroiwa, T.; Kaise, T.; Maeda, S. Arsenic in the prepared edible brown alga hijiki, Hizikia fusiforme. Appl. Organomet. Chem. 2001, 15, 561–565. [Google Scholar] [CrossRef]
- Ichikawa, S.; Nozawa, S.; Hanaoka, K.i.; Kaise, T. Ingestion and excretion of arsenic compounds present in edible brown algae, Hijikia fusiforme, by mice. Food Chem. Toxicol. 2010, 48, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, W. Official Methods of Analysis of AOAC International. Volume I, Agricultural Chemicals, Contaminants, Drugs/Edited by William Horwitz; AOAC International: Gaithersburg, MD, USA, 2010. [Google Scholar]
- Biancarosa, I.; Espe, M.; Bruckner, C.; Heesch, S.; Liland, N.; Waagbø, R.; Torstensen, B.; Lock, E. Amino acid composition, protein content, and nitrogen-to-protein conversion factors of 21 seaweed species from norwegian waters. J. Appl. Phycol. 2017, 29, 1001–1009. [Google Scholar] [CrossRef]
- Brummer, Y.; Cui, S.W. Understanding carbohydrate analysis. Food Carbohydr. Chem. Phys. Prop. Appl. 2005, 1–38. [Google Scholar] [CrossRef]
- Van Soest, P.v.; Robertson, J.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; O’Doherty, J.V.; Tiwari, B.K.; Sweeney, T.; Rajauria, G. Enhancing the extraction of polysaccharides and antioxidants from macroalgae using sequential hydrothermal-assisted extraction followed by ultrasound and thermal technologies. Mar. Drugs 2019, 17, 457. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, P.; Kumar, C.S.; Bhaskar, N. Antioxidant properties of methanol extract and its solvent fractions obtained from selected indian red seaweeds. Bioresour. Technol. 2008, 99, 2717–2723. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using folin–ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Bolanos de la Torre, A.A.S.; Henderson, T.; Nigam, P.S.; Owusu-Apenten, R.K. A universally calibrated microplate ferric reducing antioxidant power (frap) assay for foods and applications to manuka honey. Food Chem. 2015, 174, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Nicklisch, S.C.; Waite, J.H. Optimized dpph assay in a detergent-based buffer system for measuring antioxidant activity of proteins. MethodsX 2014, 1, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vaquero, M.; Rajauria, G.; Tiwari, B.; Sweeney, T.; O’Doherty, J. Extraction and yield optimisation of fucose, glucans and associated antioxidant activities from laminaria digitata by applying response surface methodology to high intensity ultrasound-assisted extraction. Mar. Drugs 2018, 16, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunton, N.P.; Mason, C.; Collins, M.J. Rapid microwave assisted preparation of fatty acid methyl esters for the analysis of fatty acid profiles in foods. J. Anal. Chem. 2015, 70, 1218–1224. [Google Scholar] [CrossRef]
- López-Alonso, M.; Miranda, M.; Benedito, J.L.; Pereira, V.; García-Vaquero, M. Essential and toxic trace element concentrations in different commercial veal cuts in spain. Meat Sci. 2016, 121, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EN. EN 15111:2007. Foodstuffs–Determination of Trace Elements–Determination of Iodine by icp-ms (Inductively Coupled Plasma Mass Spectrometry). 2007, pp. 1–12. Available online: https://0-doi-org.brum.beds.ac.uk/10.3403/30153589 (accessed on 9 March 2021).
- Available online: https://www.met.ie/climate/available-data/monthly-data (accessed on 13 February 2018).
- Available online: http://www.r-project.org (accessed on 11 February 2021).
- Friendly, M. Corrgrams: Exploratory displays for correlation matrices. Am. Stat. 2002, 56, 316–324. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximate Composition * | Macroalgae sp. | Year 2016 | Year 2017 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Winter | Spring | Summer | Autumn | Winter | Spring | Summer | Autumn | ||
| DM | L. digitata | 91.39 ± 0.01 Bc | 90.42 ± 0.01 Bd | 91.88 ± 0.04 Bb | 94.44 ± 0.05 Ba | 93.98 ± 0.00 Ac | 92.83 ± 0.04 Ad | 94.13 ± 0.01 Ab | 95.93 ± 0.01 Aa |
| Ash | 34.84 ± 0.08 Aa | 32.55 ± 0.05 Bc | 19.92 ± 0.02 Ad | 34.28 ± 0.01 Ab | 32.42 ± 0.03 Bb | 36.00 ± 0.03 Aa | 18.96 ± 0.01 Bd | 21.82 ± 0.00 Bc | |
| GE | 11.03 ± 0.03 Bc | 11.13 ± 0.00 Ab | 13.82 ± 0.01 Aa | 11.11 ± 0.01 Bbc | 11.52 ± 0.01 Ac | 10.78 ± 0.03 Bd | 13.78 ± 0.01 Aa | 13.56 ± 0.00 Ab | |
| EE | 0.26 ± 0.05 Ab | 0.51 ± 0.00 Aab | 0.87 ± 0.02 Aa | 0.66 ± 0.19 Aab | 0.38 ± 0.03 Ac | 0.03 ± 0.00 Bd | 0.74 ± 0.01 Bb | 1.12 ± 0.05 Aa | |
| NDF | 54.41 ± 0.40 Ab | 64.01 ± 0.55 Aa | 46.35 ± 0.55 Ac | 55.57 ± 0.87 Ab | 27.01 ± 1.37 Bc | 39.23 ± 0.32 Bb | 44.98 ± 0.55 Aa | 42.15 ± 0.01 Bab | |
| ADF | 16.06 ± 0.02 Bc | 23.79 ± 0.14 Aa | 22.28 ± 0.28 Ab | 15.57 ± 0.12 Bc | 20.65 ± 0.26 Ab | 24.69 ± 0.58 Aa | 10.75 ± 0.04 Bc | 23.25 ± 0.93 Aab | |
| Protein | 11.12 ± 0.76 Aa | 9.15 ± 0.00 Aa | 3.63 ± 0.04 Ab | 5.54 ± 0.03 Ab | 9.98 ± 0.09 Aa | 6.92 ± 0.02 Bb | 2.88 ± 0.06 Bd | 4.01 ± 0.04 Bc | |
| TSS | 11.88 ± 0.13 Ac | 12.41 ± 0.00 Ab | 21.03 ± 0.11 Ba | 10.88 ± 0.06 Bd | 11.75 ± 0.07 Ac | 11.19 ± 0.01 Bc | 26.94 ± 0.05 Aa | 20.39 ± 0.56 Ab | |
| DM | L. hyperborea | 90.83 ± 0.00 Bd | 92.84 ± 0.03 Bc | 94.28 ± 0.02 Ab | 95.17 ± 0.02 Ba | 95.81 ± 0.04 Aa | 95.01 ± 0.00 Ab | 94.19 ± 0.00 Ac | 95.75 ± 0.02 Aa |
| Ash | 30.01 ± 0.03 Ab | 35.64 ± 0.04 Aa | 18.36 ± 0.02 Bd | 22.34 ± 0.32 Ac | 30.33 ± 0.34 Ab | 32.79 ± 0.04 Ba | 20.82 ± 0.02 Ac | 18.91 ± 0.16 Bd | |
| GE | 12.77 ± 0.04 Ac | 12.00 ± 0.01 Ad | 14.60 ± 0.02 Aa | 13.91 ± 0.05 Bb | 11.25 ± 0.11 Bc | 12.28 ± 0.12 Ab | 14.44 ± 0.01 Aa | 14.68 ± 0.01 Aa | |
| EE | 0.76 ± 0.07 Aa | 0.75 ± 0.10 Aa | 0.65 ± 0.12 Aa | 0.80 ± 0.01 Aa | 0.60 ± 0.01 Aab | 0.56 ± 0.02 Aab | 0.34 ± 0.07 Ab | 0.69 ± 0.06 Aa | |
| NDF | 51.56 ± 0.77 Bb | 67.17 ± 0.66 Aa | 40.29 ± 0.95 Ac | 40.21 ± 0.05 Bc | 64.17 ± 0.71 Aa | 48.90 ± 0.21 Bb | 44.68 ± 0.24 Ac | 66.09 ± 1.20 Aa | |
| ADF | 19.81 ± 1.04 Aa | 21.49 ± 0.42 Ba | 22.48 ± 0.15 Aa | 21.65 ± 0.08 Aa | 21.30 ± 0.46 Ab | 35.12 ± 0.53 Aa | 20.06 ± 0.67 Ab | 10.84 ± 0.00 Bc | |
| Protein | 9.98 ± 0.01 Aa | 7.22 ± 0.01 Ab | 2.15 ± 0.13 Ad | 3.93 ± 0.06 Ac | 8.02 ± 0.04 Ba | 4.16 ± 0.04 Bb | 2.22 ± 0.27 Ac | 3.57 ± 0.00 Ab | |
| TSS | 14.49 ± 0.11 Ac | 11.85 ± 0.03 Ad | 19.21 ± 0.09 Bb | 20.15 ± 0.21 Ba | 13.73 ± 0.07 Bc | 11.23 ± 0.03 Bd | 20.42 ± 0.06 Ab | 26.69 ± 0.05 Aa | |
| DM | A. nodosum | 90.38 ± 0.01 Bd | 92.73 ± 0.08 Ab | 91.02 ± 0.02 Bc | 93.94 ± 0.04 Ba | 95.30 ± 0.08 Aa | 91.89 ± 0.01 Ad | 92.88 ± 0.01 Ac | 94.69 ± 0.00 Ab |
| Ash | 23.31 ± 0.30 Aa | 20.04 ± 0.08 Ac | 17.91 ± 0.06 Bd | 21.87 ± 0.09 Ab | 23.98 ± 0.05 Aa | 19.97 ± 0.12 Ac | 19.76 ± 0.14 Ac | 21.28 ± 0.22 Ab | |
| GE | 14.36 ± 0.02 Bc | 15.30 ± 0.06 Ab | 15.99 ± 0.05 Aa | 14.56 ± 0.02 Ac | 14.53 ± 0.03 Ab | 14.96 ± 0.02 Aa | 14.81 ± 0.04 Ba | 14.44 ± 0.13 Ab | |
| EE | 3.33 ± 0.00 Ab | 2.54 ± 0.04 Ac | 3.82 ± 0.10 Aa | 2.73 ± 0.03 Ac | 3.17 ± 0.02 Aa | 3.49 ± 0.28 Aa | 3.50 ± 0.10 Aa | 2.20 ± 0.01 Bb | |
| NDF | 53.47 ± 0.58 Aa | 56.10 ± 0.55 Aa | 52.67 ± 1.23 Aa | 53.44 ± 0.80 Aa | 49.50 ± 0.75 Ab | 56.43 ± 0.36 Aa | 46.64 ± 0.18 Ac | 41.11 ± 0.41 Bd | |
| ADF | 21.95 ± 0.37 Ab | 30.35 ± 0.34 Aa | 12.15 ± 0.39 Bc | 29.76 ± 0.00 Aa | 16.37 ± 0.16 Bb | 16.05 ± 0.56 Bb | 16.26 ± 0.21 Ab | 20.04 ± 0.30 Ba | |
| Protein | 6.14 ± 0.01 Aa | 5.22 ± 0.09 Ab | 2.68 ± 0.00 Ad | 3.52 ± 0.01 Ac | 4.03 ± 0.05 Bb | 3.60 ± 0.02 Bc | 2.25 ± 0.06 Ad | 4.44 ± 0.11 Aa | |
| TSS | 13.66 ± 0.08 Ab | 14.27 ± 0.07 Aa | 13.34 ± 0.03 Ab | 11.63 ± 0.06 Bc | 12.43 ± 0.02 Bc | 13.02 ± 0.15 Bb | 12.33 ± 0.11 Ac | 14.63 ± 0.01 Aa | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Vaquero, M.; Rajauria, G.; Miranda, M.; Sweeney, T.; Lopez-Alonso, M.; O’Doherty, J. Seasonal Variation of the Proximate Composition, Mineral Content, Fatty Acid Profiles and Other Phytochemical Constituents of Selected Brown Macroalgae. Mar. Drugs 2021, 19, 204. https://0-doi-org.brum.beds.ac.uk/10.3390/md19040204
Garcia-Vaquero M, Rajauria G, Miranda M, Sweeney T, Lopez-Alonso M, O’Doherty J. Seasonal Variation of the Proximate Composition, Mineral Content, Fatty Acid Profiles and Other Phytochemical Constituents of Selected Brown Macroalgae. Marine Drugs. 2021; 19(4):204. https://0-doi-org.brum.beds.ac.uk/10.3390/md19040204
Chicago/Turabian StyleGarcia-Vaquero, Marco, Gaurav Rajauria, Marta Miranda, Torres Sweeney, Marta Lopez-Alonso, and John O’Doherty. 2021. "Seasonal Variation of the Proximate Composition, Mineral Content, Fatty Acid Profiles and Other Phytochemical Constituents of Selected Brown Macroalgae" Marine Drugs 19, no. 4: 204. https://0-doi-org.brum.beds.ac.uk/10.3390/md19040204