
Influence of Carbohydrate Additives on the Growth Rate of Microalgae Biomass with an Increased Carbohydrate Content


 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Microalgae Identification Results
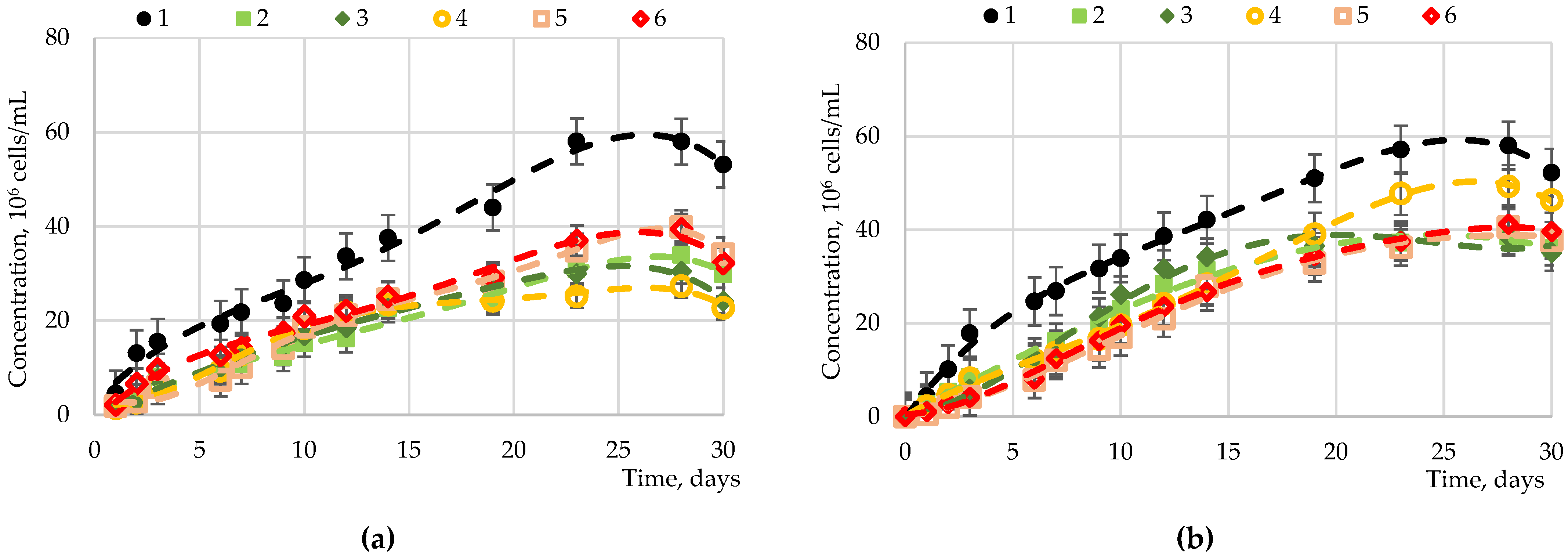
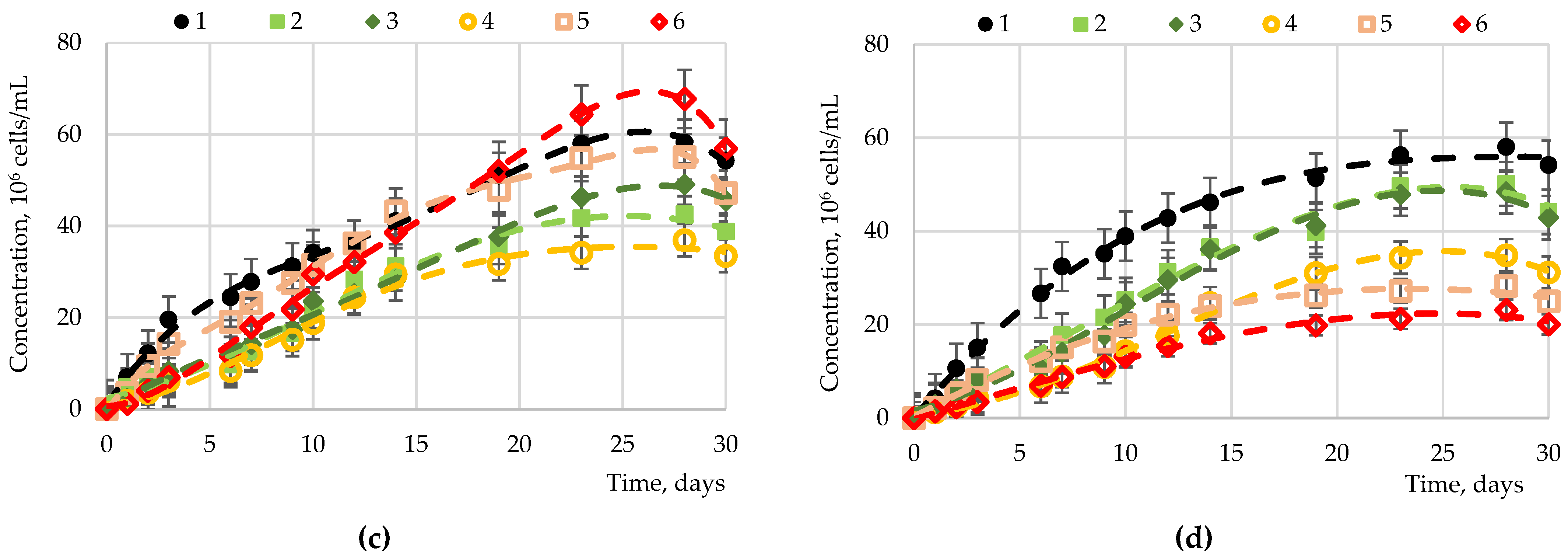
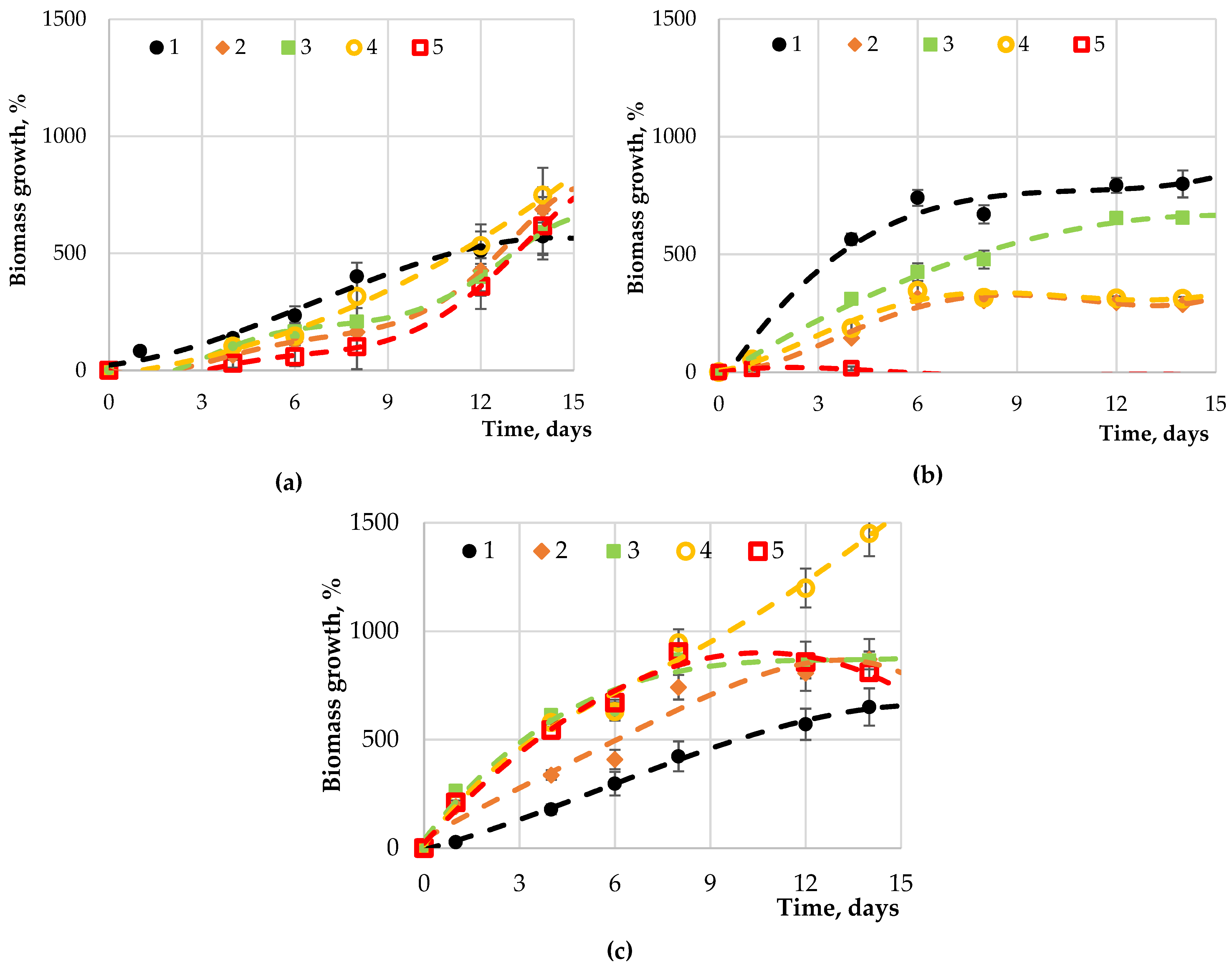
2.2. The Results of the Introduction of Carbohydrates into Nutrient Media for Microalgae Cultivation
2.3. Results of Determining the Content of Carbohydrates in Microalgae
3. Discussion
4. Materials and Methods
4.1. Microalgae Sampling
4.2. Microalgae Biomass Cultivation
4.3. Microalgae Morphology Determination
4.4. Introduction of Carbohydrate Additives to the Nutrient Medium
4.5. Chlorophyll Extraction
4.6. Carbohydrate Fraction Isolation
4.7. Determination of Carbohydrate Content
4.8. Determination of the Residual Amount of Sugars in Nutrient Media
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Sequences of the 18S Ribosomal RNA Gene of the Chlorella vulgaris Beijer
Appendix B. Sequences of the 18S Ribosomal RNA Gene of the Arthrospira platensis Gomont
Appendix C. Sequences of the 18S Ribosomal RNA Gene of the Dunaliella salina Teod
References
- Zhou, X.; Lin, W.; Tong, L.; Liu, X.; Zhong, K.; Liu, L.; Wang, L.; Zhou, S. Hypolipidaemic effects of oat flakes and beta-glucans derived from four Chinese naked oat (Avena nuda) cultivars in Wistar-Lewis rats. J. Sci. Food Agric. 2016, 96, 644–649. [Google Scholar] [CrossRef]
- Kharwar, S.; Bhattacharjee, S.; Mishra, A.K. disentangling the impact of sulfur limitation on exopolysaccharide and functionality of Alr2882 by in silico approaches in Anabaena sp. PCC 7120. Appl. Biochem. Biotechnol. 2021, 193, 1447–1468. [Google Scholar] [CrossRef]
- Vieira, M.V.; Pastrana, L.M.; Fuciños, P. Microalgae encapsulation systems for food, pharmaceutical and cosmetics applications. Mar. Drugs 2020, 18, 644. [Google Scholar] [CrossRef]
- Ma, R.; Wang, B.; Chua, E.T.; Zhao, X.; Lu, K.; Ho, S.H.; Shi, X.; Liu, L.; Xie, Y.; Lu, Y.; et al. Comprehensive utilization of marine microalgae for enhanced co-production of multiple compounds. Mar. Drugs 2020, 18, 467. [Google Scholar] [CrossRef] [PubMed]
- Rumin, J.; Nicolau, E.; Junior, R.G.O.; Fuentes-Grünewald, C.; Picot, L. Analysis of scientific research driving microalgae market opportunities in Europe. Mar. Drugs 2020, 18, 264. [Google Scholar] [CrossRef] [PubMed]
- Bodachivskyi, I.; Kuzhiumparambil, U.; Bradley, G.; Williams, D. High yielding acid-catalysed hydrolysis of cellulosic polysaccharides and native biomass into low molecular weight sugars in mixed ionic liquid systems. Chem. Open 2019, 8, 1316–1324. [Google Scholar] [CrossRef] [PubMed]
- Koyande, A.K.; Show, P.L.; Guo, R.; Tang, B.; Ogino, C.; Chang, J.S. Bio-processing of algal bio-refinery: A review on current advances and future perspectives. Bioengineered 2019, 10, 574–592. [Google Scholar] [CrossRef] [Green Version]
- Lam, G.P.; Giraldo, J.B.; Vermue, M.H.; Olivieri, G.; Eppink, M.H.; Wijffels, R.H. Understanding the salinity effect on cationic polymers in inducing flocculation of the microalga Neochloris oleoabundans. J. Biotechnol. 2016, 225, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Lakatos, G.E.; Ranglová, K.; Manoel, J.C.; Grivalský, T.; Kopecký, J.; Masojídek, J. Bioethanol production from microalgae polysaccharides. Folia. Microbiol. 2019, 64, 627–644. [Google Scholar] [CrossRef]
- Andreeva, A.; Budenkova, E.; Babich, O.; Sukhikh, S.; Ulrikh, E.; Ivanova, S.; Prosekov, A.; Dolganyuk, V. Production, purification, and study of the amino acid composition of microalgae proteins. Molecules 2021, 26, 2767. [Google Scholar] [CrossRef]
- Klanchui, A.; Dulsawat, S.; Chaloemngam, K.; Cheevadhanarak, S.; Prommeenate, P.; Meechai, A. An improved genome-scale metabolic model of Arthrospira platensis C1 (iAK888) and its application in glycogen overproduction. Metabolites 2018, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Aytenfisu, A.H.; Yang, M.; MacKerell, A.D. Drude polarizable force field for glycosidic linkages involving pyranoses and furanoses. J. Chem. Theory Comput. 2018, 14, 3132–3143. [Google Scholar] [CrossRef]
- Cheng, D.; Li, D.; Yuan, Y.; Zhou, L.; Li, X.; Wu, T.; Wang, L.; Zhao, Q.; Wei, W.; Sun, Y. Improving carbohydrate and starch accumulation in Chlorella sp. AE10 by a novel two-stage process with cell dilution. Biotechnol. Biofuels 2017, 10, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, C.; Wetzel, M.; Reinhardt, J.; Schmidt, M.; Felten, L.; Mundt, S. Screening of microalgae for primary metabolites including β-glucans and the influence of nitrate starvation and irradiance on β-glucan production. J. Appl. Phycol. 2016, 28, 2719–2725. [Google Scholar] [CrossRef]
- Khan, M.I.; Lee, M.G.; Seo, H.J.; Shin, J.H.; Shin, T.S.; Yoon, Y.H.; Kim, M.Y.; Choi, J.I.; Kim, J.D. Enhancing the feasibility of Microcystis aeruginosa as a feedstock for bioethanol production under the influence of various factors. Biomed. Res. Int. 2016, 2016, 4540826. [Google Scholar] [CrossRef] [Green Version]
- Fimbres-Olivarría, D.; López-Elías, J.A.; Carvajal-Millán, E.; Márquez-Escalante, J.A.; Martínez-Córdova, L.R.; Miranda-Baeza, A.; Enríquez-Ocaña, F.; Valdéz-Holguín, J.E.; Brown-Bojórquez, F. Navicula sp. sulfated polysaccharide gels induced by Fe(III): Rheology and microstructure. Int. J. Mol. Sci. 2016, 17, 1238. [Google Scholar] [CrossRef] [Green Version]
- Mooij, P.R.; de Jongh, L.D.; van Loosdrecht, M.C.; Kleerebezem, R. Influence of silicate on enrichment of highly productive microalgae from a mixed culture. J. Appl. Phycol. 2016, 28, 1453–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markou, G.; Depraetere, O.; Vandamme, D.; Muylaert, K. Cultivation of Chlorella vulgaris and Arthrospira platensis with recovered phosphorus from wastewater by means of zeolite sorption. Int. J. Mol. Sci. 2015, 16, 4250–4264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, N.C.; Panwar, A.; Bisht, T.S.; Tamta, S. Coupling of algal biofuel production with wastewater. Sci. World J. 2014, 2014, 210504. [Google Scholar] [CrossRef] [Green Version]
- Patel, D.S.; He, X.; MacKerell, A.D., Jr. Polarizable empirical force field for hexopyranose monosaccharides based on the classical Drude oscillator. J. Phys. Chem. B 2015, 119, 637–652. [Google Scholar] [CrossRef]
- Patel, D.S.; Pendrill, R.; Mallajosyula, S.S.; Widmalm, G.; MacKerell, A.D., Jr. Conformational properties of α- or β-(1→6)-linked oligosaccharides: Hamiltonian replica exchange MD simulations and NMR experiments. J. Phys. Chem. B 2014, 118, 2851–2871. [Google Scholar] [CrossRef]
- Park, D.; Jagtap, S.; Nair, S.K. Structure of a PL17 family alginate lyase demonstrates functional similarities among exotype depolymerases. J. Biol. Chem. 2014, 289, 8645–8655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühlroth, A.; Li, K.; Røkke, G.; Winge, P.; Olsen, Y.; Hohmann-Marriott, M.F.; Vadstein, O.; Bones, A.M. Pathways of lipid metabolism in marine algae, co-expression network, bottlenecks and candidate genes for enhanced production of EPA and DHA in species of Chromista. Mar. Drugs 2013, 11, 4662–4697. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Tan, L.; Jin, X.; Wu, H.; Houbo, W.; Tao, L.; Wenzhou, X. Evaluating the potential of carbohydrate-rich microalga Rhodosorus sp. SCSIO-45730 as a feedstock for biofuel and β-glucans using strategies of phosphate optimization and low-cost harvest. J. Appl. Phycol. 2020, 32, 3051–3061. [Google Scholar] [CrossRef]
- Blockx, J.; Verfaillie, A.; Thielemans, W.; Muylaert, K. Unravelling the mechanism of chitosan-driven flocculation of microalgae in seawater as a function of pH. ACS Sustain. Chem. Eng. 2018, 6, 11273–11279. [Google Scholar] [CrossRef]
- Chng, L.M.; Lee, K.T.; Chan, D.J.C. Synergistic effect of pretreatment and fermentation process on carbohydrate-rich Scenedesmus dimorphus for bioethanol production. Energy Convers. Manag. 2017, 141, 410–419. [Google Scholar] [CrossRef]
- Corrêa, D.O.; Duarte, M.E.R.; Noseda, M.D. Biomass production and harvesting of Desmodesmus subspicatus cultivated in flat plate photobioreactor using chitosan as flocculant agent. J. Appl. Phycol. 2018, 31, 857–866. [Google Scholar] [CrossRef]
- Gerchman, Y.; Vasker, B.; Tavasi, M.; Mishael, Y.; Kinel-Tahan, Y.; Yehoshua, Y. Effective harvesting of microalgae: Comparison of different polymeric flocculants. Bioresour. Technol. 2017, 228, 141–146. [Google Scholar] [CrossRef]
- Varshney, P.; Beardall, J.; Bhattacharya, S.; Wangikar, P.P. Isolation and biochemical characterisation of two thermophilic green algal species—Asterarcys quadricellulare and Chlorella sorokiniana, which are tolerant to high levels of carbon dioxide and nitric oxide. Algal Res. 2018, 30, 28–37. [Google Scholar] [CrossRef]
- Berteotti, S.; Ballottari, M.; Bassi, R. Increased biomass productivity in green algae by tuning nonphotochemical quenching. Sci. Rep. 2016, 6, 21339. [Google Scholar] [CrossRef] [Green Version]
- Schulze, P.S.C.; Pereira, H.G.C.; Santos, T.F.C.; Schueler, L.; Guerra, R.; Barreira, L.A.; Perales, J.A.; Varela, J.C.S. Effect of light quality supplied by light emitting diodes (LEDs) on growth and biochemical profiles of Nannochloropsis oculat and Tetraselmis chuii. Algal Res. 2016, 16, 387–398. [Google Scholar] [CrossRef]
- Gupta, S.K.; Kumar, N.M.; Guldhe, A.; Ansari, F.A.; Rawat, I.; Nasr, M.; Bux, F. Wastewater to biofuels: Comprehensive evaluation of various flocculants on biochemical composition and yield of microalgae. Ecol. Eng. 2018, 117, 62–68. [Google Scholar] [CrossRef]
- Krayesky-Self, S.; Phung, D.; Schmidt, W.; Sauvage, T.; Butler, L.; Fredericq, S. First report of endolithic members of Rhodosorus marinus (Stylonematales, Rhodophyta) growing inside rhodoliths offshore Louisiana, Northwestern Gulf of Mexico. Front. Mar. Sci. 2020, 7, 1–8. [Google Scholar] [CrossRef]
- Li, T.; Xu, J.; Wu, H.; Jiang, P.; Chen, Z.; Xiang, W. Growth and biochemical composition of Porphyridium purpureum SCS-02 under different nitrogen concentrations. Mar. Drugs 2019, 17, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolganyuk, V.; Andreeva, A.; Budenkova, E.; Sukhikh, S.; Babich, O.; Ivanova, S.; Prosekov, A.; Ulrikh, E. Study of morphological features and determination of the fatty acid composition of the microalgae lipid complex. Biomolecules 2020, 10, 1571. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.-H.; Huang, S.-W.; Chen, C.-Y.; Hasunuma, T.; Kondo, A.; Chang, J.-S. Characterization and optimization of carbohydrate production from an indigenous microalga Chlorella vulgaris FSP-E. Bioresour. Techno. 2013, 135, 157–165. [Google Scholar] [CrossRef]
- Chou, N.T.; Cheng, C.F.; Wu, H.C.; Lai, C.P.; Lin, L.T.; Pan, I.H.; Ko, C.H. Chlorella sorokiniana-induced activation and maturation of human monocyte-derived dendritic cells through NF-κB and PI3K/MAPK pathways. Evid. Based Complement. Altern. Med. 2012, 2012, 735396. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yao, C.; Sun, Y.; Chen, W.; Tan, H.; Cao, X.; Xue, S.; Yin, H. Production and structural characterization of a new type of polysaccharide from nitrogen-limited Arthrospira platensis cultivated in outdoor industrial-scale open raceway ponds. Biotechnol. Biofuels 2019, 12, 131. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, N.E.; Hussein, M.H.; Shaaban-Dessuuki, S.A.; Dalal, S.R. Production, extraction and characterization of Chlorella vulgaris soluble polysaccharides and their applications in AgNPs biosynthesis and biostimulation of plant growth. Sci. Rep. 2020, 10, 3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaignard, C.; Laroche, C.; Pierre, G.; Dubessay, P.; Delattre, C.; Gardarin, C.; Gourvil, P.; Probert, I.; Dubuffet, A.; Michaud, P. Screening of marine microalgae: Investigation of new exopolysaccharide producers. Algal Res. 2019, 44, 101711. [Google Scholar] [CrossRef]
- Dolganyuk, V.; Belova, D.; Babich, O.; Prosekov, A.; Ivanova, S.; Katserov, D.; Patyukov, N.; Sukhikh, S. Microalgae: A promising source of valuable bioproducts. Biomolecules 2020, 10, 1153. [Google Scholar] [CrossRef] [PubMed]
- Sero, E.T.; Siziba, N.; Bunhu, T.; Shoko, R.; Jonathan, E. Biophotonics for improving algal photobioreactor performance: A review. Int. J. Energy Res. 2020, 44, 5071–5092. [Google Scholar] [CrossRef]
- Toor, M.; Kumar, S.S.; Malyan, S.K.; Bishnoi, N.R.; Mathimani, T.; Rajendran, K.; Pugazhendhi, A. An overview on bioethanol production from lignocellulosic feedstocks. Chemosphere 2019, 242, 125080. [Google Scholar] [CrossRef] [PubMed]
- Yamin, W.A.; Shaleh, S.R.M.; Ching, F.F.; Othman, R.; Manjaji-Matsumoto, M.; Mustafa, S.; Shigeharu, S.; Kandasamy, G. Harvesting Chaetoceros gracilis by flocculation using chitosan. IOP Conf. Ser. Earth Environ. Sci. 2019, 236, 012123. [Google Scholar] [CrossRef]
- Zhu, Q.; Wu, S. Water-soluble beta-1,3-glucan prepared by degradation of curdlan with hydrogen peroxide. Food Chem. 2019, 283, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Rojo-Cebreros, A.H.; Ibarra-Castro, L.; Martínez-Brown, J.M.; Velasco-Blanco, G.; Martínez-Téllez, M.A.; Medina-Jasso, M.A.; Nieves-Soto, M.; Quintana-Zavala, D. Potential of Nannochloropsis in beta glucan production. In Nannochloropsis: Biology, Biotechnological, Potential and Challenges; Jan, M., Kazik, P., Eds.; Nova Sciences Publishers Inc.: New York, NY, USA, 2017; pp. 181–225. [Google Scholar]
- Welham, S.J.; Gezan, S.A.; Clark, S.J.; Mead, A. Statistical Methods in Biology: Design and Analysis of Experiments and Regression; CRC Press (Taylor & Francis Group): Boca Raton, FL, USA, 2014. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbohydrates | Concentration, % of Dry Matter | |||||
|---|---|---|---|---|---|---|
| I | II | III | ||||
| exp. | Control | exp. | Control | exp. | Control | |
| 1 | 0.12 ± 0.02 a | 0.10 ± 0.01 | 0.39 ± 0.08 a | 0.42 ± 0.05 | 0.38 ± 0.01 a | 0.33 ± 0.01 |
| 2 | 11.26 ± 0.72 b | 9.78 ± 0.22 | 3.41 ± 0.62 b | 2.32 ± 0.30 | 1.78 ± 0.29 b | 1.60 ± 0.22 |
| 3 | 0.12 ± 0.03 a | 0.11 ± 0.02 | 0.21 ± 0.02 a | 0.20 ± 0.02 | 0.21 ± 0.01 a | 0.22 ± 0.01 |
| 4 | 0.33 ± 0.06 a | 0.27 ± 0.03 | 0.31 ± 0.09 a | 0.32 ± 0.07 | 0.11 ± 0.06 a | 0.10 ± 0.02 |
| 5 | 16.97 ± 0.64 c | 15.12 ± 0.48 | 9.59 ± 0.20 c | 7.81 ± 0.76 | 8.68 ± 0.74 c | 7.54 ± 0.30 |
| Carbohydrates | Concentration, % of Dry Matter | |||||
|---|---|---|---|---|---|---|
| I | II | III | ||||
| exp. | Control | exp. | Control | exp. | Control | |
| 1 | 0.11 ± 0.02 a | 0.10 ± 0.01 | 0.37 ± 0.08 a | 0.42 ± 0.05 | 0.33 ± 0.01 a | 0.33 ± 0.01 |
| 2 | 10.48 ± 0.72 b | 9.78 ± 0.22 | 3.40 ± 0.62 b | 2.32 ± 0.30 | 1.76 ± 0.29 b | 1.60 ± 0.22 |
| 3 | 0.11 ± 0.03 a | 0.11 ± 0.02 | 0.19 ± 0.02 a | 0.20 ± 0.02 | 0.20 ± 0.01 a | 0.22 ± 0.01 |
| 4 | 0.29 ± 0.06 a | 0.27 ± 0.03 | 0.30 ± 0.09 a | 0.32 ± 0.07 | 0.10 ± 0.06 a | 0.10 ± 0.02 |
| 5 | 15.99 ± 0.64 c | 15.12 ± 0.48 | 8.98 ± 0.20 c | 7.81 ± 0.76 | 8.66 ± 0.74 c | 7.54 ± 0.30 |
| Carbohydrates | Concentration, % of Dry Matter | ||
|---|---|---|---|
| I | II | III | |
| 1 | 1.0 ± 0.2 a | 3.0 ± 0.6 a | 2.3 ± 0.3 a |
| 2 | 1.3 ± 0.2 a | 4.1 ± 0.5 b | 9.1 ± 0.6 b |
| 3 | 2.1 ± 0.3 b | 1.2 ± 0.2 c | 2.0 ± 0.1 a |
| 4 | 2.0 ± 0.3 b | 2.0 ± 0.3 c | 1.1 ± 0.2 c |
| 5 | 1.9 ± 0.4 b | 1.1 ± 0.2 c | 0.9 ± 0.4 c |
| Cultivation Time, Days | Concentration, % of Dry Matter | ||
|---|---|---|---|
| I | II | III | |
| 0 | 0.08 ± 0.02 a | 0.09 ± 0.03 a | 0.02 ±0.06 a |
| 4 | 2.43 ± 0.31 b | 1.59 ± 0.05 b | 0.45 ± 0.14 b |
| 8 | 9.88 ± 0.46 c | 5.29 ± 0.18 c | 0.78 ± 0.23 b |
| 12 | 16.97 ± 0.64 d | 9.59 ± 0.20 d | 1.13 ± 0.51 c |
| 16 | 4.58 ± 0.34 e | 2.62 ± 0.12 e | 1.27 ± 0.62 c |
| 20 | - | - | 1.69 ± 0.66 c |
| 24 | - | - | 2.37 ±0.71 d |
| 28 | - | - | 8.68 ± 0.74 e |
| 30 | - | - | 0.97 ± 0.47 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andreeva, A.; Budenkova, E.; Babich, O.; Sukhikh, S.; Dolganyuk, V.; Michaud, P.; Ivanova, S. Influence of Carbohydrate Additives on the Growth Rate of Microalgae Biomass with an Increased Carbohydrate Content. Mar. Drugs 2021, 19, 381. https://0-doi-org.brum.beds.ac.uk/10.3390/md19070381
Andreeva A, Budenkova E, Babich O, Sukhikh S, Dolganyuk V, Michaud P, Ivanova S. Influence of Carbohydrate Additives on the Growth Rate of Microalgae Biomass with an Increased Carbohydrate Content. Marine Drugs. 2021; 19(7):381. https://0-doi-org.brum.beds.ac.uk/10.3390/md19070381
Chicago/Turabian StyleAndreeva, Anna, Ekaterina Budenkova, Olga Babich, Stanislav Sukhikh, Vyacheslav Dolganyuk, Philippe Michaud, and Svetlana Ivanova. 2021. "Influence of Carbohydrate Additives on the Growth Rate of Microalgae Biomass with an Increased Carbohydrate Content" Marine Drugs 19, no. 7: 381. https://0-doi-org.brum.beds.ac.uk/10.3390/md19070381