
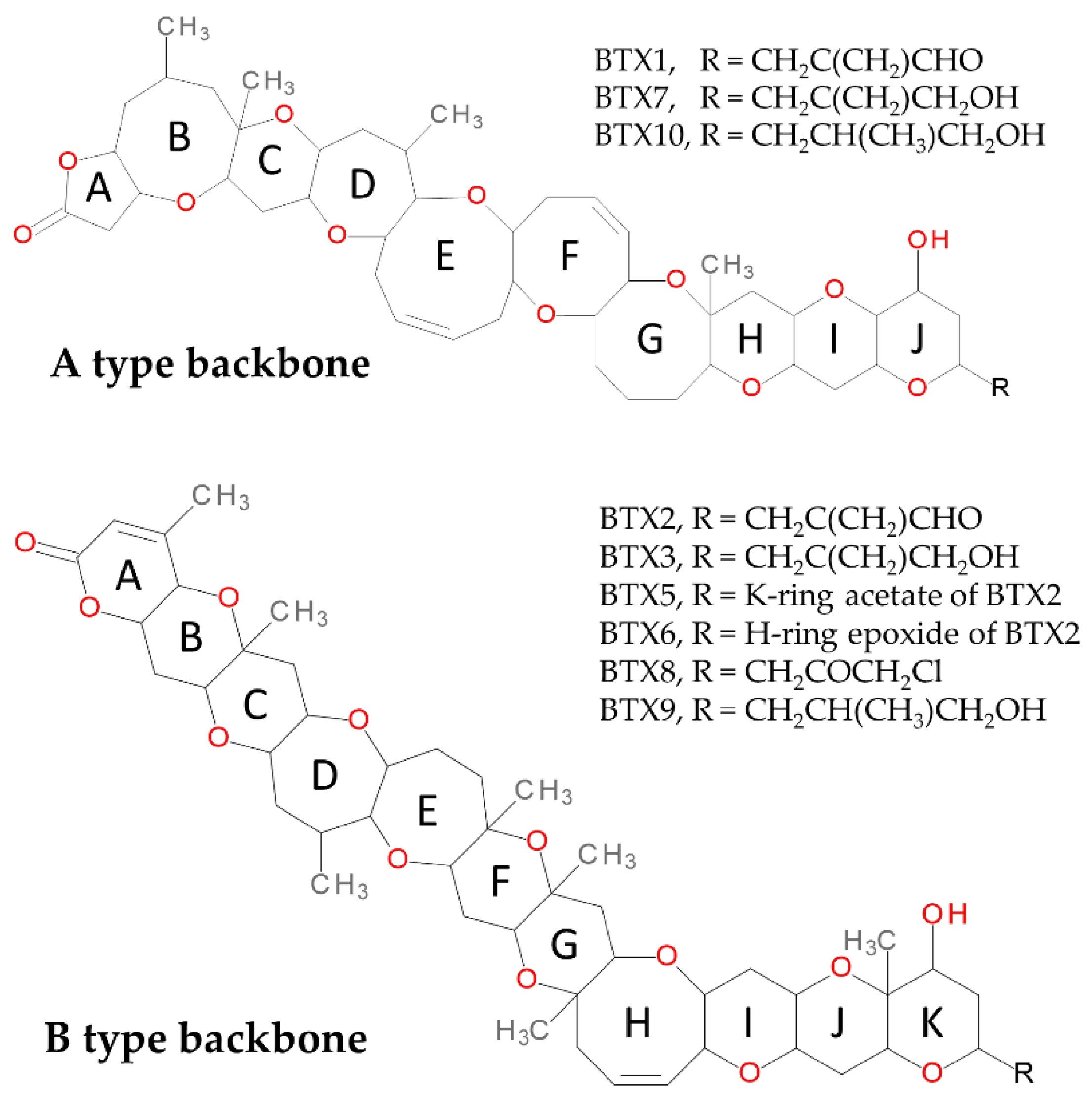
Monitoring the Emergence of Algal Toxins in Shellfish: First Report on Detection of Brevetoxins in French Mediterranean Mussels

 , ,
, ,
Abstract
:1. Introduction
2. Results
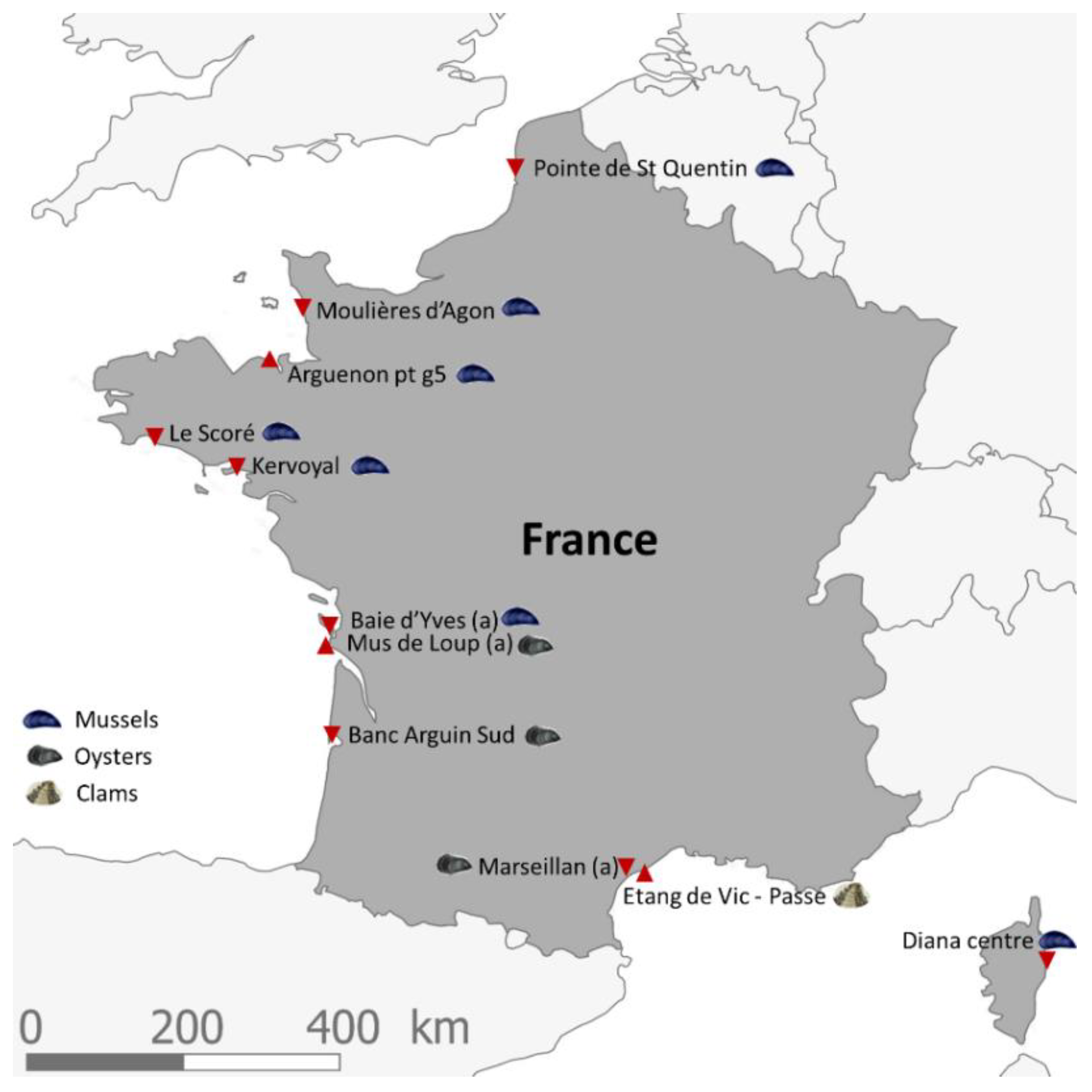
2.1. Criteria for Selecting the Shellfish Sampling Points of the EMERGTOX Network
- their location in shellfish harvesting areas (mussels, oysters or clams) that are active all year round;
- an equal geographical distribution over the French coastlines;
- the existence of historical data for the sampling points, with the aim to obtain long time series;
- the existence of previous mousse bioassays (which was the reference test prior to 2010 for the monitoring of lipophilic toxins) that remain unexplained (short survival time and/or neurological symptoms);
- their location outside of risk areas.

2.2. LC-MS/MS Lipophilic Multi-Toxin Quantification in Shellfish
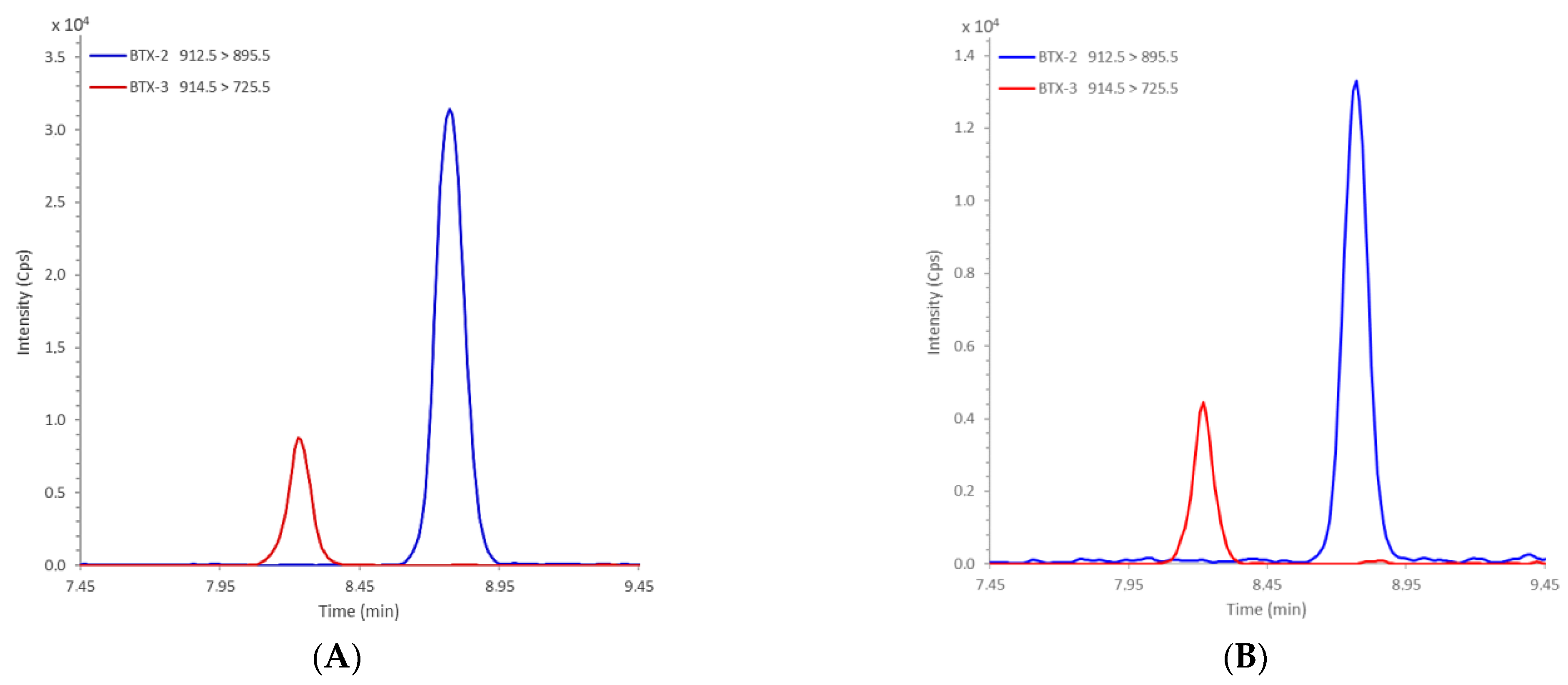
2.2.1. Optimization and Internal Validation of the LC-MS/MS Lipophilic Multi-Toxin Analysis Method in Shellfish
2.2.2. Detection of BTXs at Diana Lagoon
2.3. Identification of Planktonic Species Potentially Producing BTXs in Water Samples from Diana Lagoon
3. Discussion
3.1. Contamination of Corsica Mussels by BTXs
3.2. Phytoplankton Potentially Producing BTXs and Other Metabolites
4. Materials and Methods
4.1. Materials
4.2. Shellfish and Seawater Samples
4.2.1. Seawater for Phytoplankton Observations
4.2.2. Homogenates of Shellfish for Toxin Analysis
4.3. Methods
4.3.1. Microalgae Identification
4.3.2. Extraction of Lipophilic Toxins Including BTXs
4.3.3. LC-MS/MS Analysis of Toxins
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abraham, A.; Wang, Y.; El Said, K.R.; Plakas, S.M. Characterisation of brevetoxin metabolisms in Karenia brevis bloom-exposed clams (Mercenaria sp.) by LC-MS/MS. Toxicon 2012, 60, 1030–1040. [Google Scholar] [CrossRef]
- Chou, H.N.; Shimizu, Y. A new polyether toxin from Gymnodinium breve Davis. Tetrahedron Lett. 1982, 23, 5521–5524. [Google Scholar] [CrossRef]
- Shimizu, Y.; Chou, H.N.; Bando, H.; Van Duyne, G.; Clardy, J. Structure of Brevetoxin A (GB-1 Toxin), the Most Potent Toxin in the Florida Red Tide Organism Gymnodinium breve (Ptychodiscus brevis). J. Am. Chem. Soc. 1986, 108, 514–515. [Google Scholar] [CrossRef]
- Lin, Y.-Y.; Risk, M.; Ray, S.M.; Van Engen, D.; Clardy, J.; Golik, J.; James, J.C.; Nakanishi, K. Isolation and Structure of Brevetoxin B from the “Red Tide” Dinoflagellate Ptychodiscus brevis (Gymnodinium breve). J. Am. Chem. Soc. 1981, 103, 6773–6775. [Google Scholar] [CrossRef]
- Poli, M.A.; Mende, T.J.; Baden, D.G. Brevetoxins, unique activators of voltage-sensitive sodium channels, bind to specific sites in rat brain synaptosomes. Mol. Pharmacol. 1986, 30, 129–135. [Google Scholar]
- Baden, D.G.; Bourdelais, A.J.; Jacocks, H.; Michelliza, S.; Naar, J. Natural and derivative brevetoxins: Historical background, multiplicity, and effects. Environ. Health Perspect. 2005, 113, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Plakas, S.M.; Wang, Z.; Jester, E.L.E.; El Said, K.R.; Granade, H.R.; Henry, M.S.; Blum, P.C.; Pierce, R.H.; Dickey, R.W. Characterization of polar brevetoxin derivatives isolated from Karenia brevis cultures and natural blooms. Toxicon 2006, 48, 104–115. [Google Scholar] [CrossRef]
- Brovedani, V.; Pelin, M.; D’Orlando, E.; Poli, M. Brevetoxins: Toxicological Profile. In Marine and Freshwater Toxins, Toxinology; Gopalakrishnakone, P., Haddad, V., Jr., Tubaro, A., Kim, E., Kem, W.R., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 113–127. [Google Scholar]
- Kusek, K.M.; Vargo, G.; Steidinger, K. Gymnodinium breve in the field, in the lab, and in the newspaper- a scientific and journalistic analysis of Florida red tides. Contrib. Mar. Sci. 1999, 34, 1–228. [Google Scholar]
- Ramsdell, J.S. The molecular and integrative basis to brevetoxin toxicity. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; Botana, L., Ed.; Taylor & Francis: London, UK, 2008; pp. 519–550. [Google Scholar]
- Fleming, L.E.; Backer, L.C.; Baden, D.G. Overview of aerosolized Florida red tide toxins: Exposures and effects. Environ. Health Perspect. 2005, 113, 618–620. [Google Scholar] [CrossRef] [Green Version]
- Backer, L.C.; Fleming, L.E.; Rowan, A.; Cheng, Y.-S.; Benson, J.; Pierce, R.H.; Zaias, J.; Bean, J.; Bossart, G.D.; Johnson, D.; et al. Recreational exposure to aerosolized brevetoxins during Florida red tide events. Harmful Algae 2003, 2, 19–28. [Google Scholar] [CrossRef]
- Pierce, R.H.; Henry, M.S.; Blum, P.C.; Lyons, J.; Cheng, Y.S.; Yazzie, D.; Zhou, Y. Brevetoxin concentrations in marine aerosol: Human exposure levels during a Karenia brevis harmful algal bloom. Bull. Environ. Contam. Toxicol. 2003, 70, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.S.; Villareal, T.A.; Zhou, Y.; Gao, J.; Pierce, R.H.; Wetzel, D.; Naar, J.; Baden, D.G. Characterization of red tide aerosol on the Texas coast. Harmful Algae 2005, 4, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Naar, J.P.; Flewelling, L.J.; Lenzi, A.; Abbott, J.P.; Granholm, A.; Jacocks, H.M.; Gannon, D.; Henry, M.; Pierce, R.; Baden, D.G.; et al. Brevetoxins, like ciguatoxins, are potent ichthyotoxic neurotoxins that accumulate in fish. Toxicon 2007, 50, 707–723. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Scientific Opinion on marine biotoxins in shellfish—Emerging toxins: Brevetoxin group. EFSA J. 2010, 8, 1677. [CrossRef]
- Plakas, S.M.; Dickey, R.W. Advances in monitoring and toxicity assessment of brevetoxins in molluscan shellfish. Toxicon 2010, 56, 137–149. [Google Scholar] [CrossRef]
- Watkins, S.M.; Reich, A.; Fleming, L.E.; Hammond, R. Neurotoxic shellfish poisoning. Mar. Drugs 2008, 6, 431–455. [Google Scholar] [CrossRef] [Green Version]
- Gordon, G.; Williams, R.H.; Davis, C.C.; Walton Smith, F.G. Catastrophic mass mortality of marine animals and coincident phytoplankton bloom on the west coast of Florida, November 1946 to August 1947. Ecol. Monogr. 1948, 18, 309–324. [Google Scholar]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Landsberg, J.H.; Flewelling, L.J.; Naar, J. Karenia brevis red tides, brevetoxins in the food web, and impacts on natural resources: Decadal advancements. Harmful Algae 2009, 8, 598–607. [Google Scholar] [CrossRef]
- Lassus, P.; Chomérat, N.; Hess, P.; Nézan, E. Toxic and Harmful Microalgae of the World Ocean/Micro-Algues Toxiques et Nuisibles de L’océan Mondial. Denmark, International Society for the Study of Harmful Algae/Intergovernmental Oceanographic Commission of UNESCO. IOC Manuals and Guides, 68 (Bilinual English/French). 2016, p. 523. Available online: https://archimer.ifremer.fr/doc/00379/49054/ (accessed on 29 June 2021).
- Morohashi, A.; Satake, M.; Murata, K.; Naoki, H.; Kaspar, H.F.; Yasumoto, T. Brevetoxin B3, a New Brevetoxin Analog Isolated from the Greenshell Mussel Perna canaliculus Involved in Neurotoxic Shellfish Poisoning in New Zealand. Tetrahedron Lett. 1995, 36, 8995–8998. [Google Scholar] [CrossRef]
- Morris, P.D.; Campbell, D.S.; Taylor, T.J.; Freeman, J.I. Clinical and epidemiological features of neurotoxic shellfish poisoning in North Carolina. Am. J. Publ. Health 1991, 81, 471–474. [Google Scholar] [CrossRef] [Green Version]
- ANSES. Opinion of the French Agency for Food, Environmental and Occupational Health & Safety of 2 March 2021 on the State of Knowledge on Brevetoxins in Shellfish, Data on Toxicity, Occurrence and Brevetoxin-Producing Microalgae (Request No 2020-SA-0020); The opinion is accompanied by a collective expert appraisal report; ANSES: Maisons-Alfort, France, 2021; p. 18. (In French)
- Derrien, A.; Terre Terrillon, A.; Duval, A.; Martin, C.; Sibat, M.; Rovillon, G.A.; Réveillon, D.; Amzil, Z. From development to implementation of a new LC-MS/MS approach to quantify 43 lipophilic toxins within the French monitoring program. In Proceedings of the 18th International Conference on Harmful Algae “From Ecosystems to Socio-Ecosystems”, Nantes, France, 21–26 October 2018; p. 415. [Google Scholar]
- Abraham, A.; El Said, K.R.; Flewelling, L.J. Role of Biomarkers in Monitoring Brevetoxins in Karenia brevis Exposed Shellfish. Food Saf. 2018, 6, 33–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flewelling, L.J.; Corcoran, A.A.; Granholm, A.A.; Takeuchi, N.Y.; Van Hoeck, R.V.; Zahara, M.L. Validation and Assessment of an Enzyme-Linked Immunosorbent Assay (Elisa) for Use in Monitoring and Managing Neurotoxic Shellfish Poisoning. J. Shellfish Res. 2020, 39, 491–500. [Google Scholar] [CrossRef]
- Roth, P.B.; Twiner, M.J.; Wang, Z.; Bottein Dechraoui, M.-Y.; Doucette, G.J. Fate and distribution of brevetoxin (PbTx) following lysis of Karenia brevis by algicidal bacteria, including analysis of open A-ring derivatives. Toxicon 2007, 50, 1175–1191. [Google Scholar] [CrossRef]
- Fowler, N.; Tomas, C.; Baden, D.; Campbell, L.; Bourdelais, A. Chemical analysis of Karenia papilionacea. Toxicon 2015, 101, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Band-Schmidt, C.J.; Martínez-López, A.; Bustillos-Guzmán, J.J.; Carreón-Palau, L.; Morquecho, L.; Olguín-Monroy, N.O.; Zenteno-Savín, T.; Mendoza-Flores, A.; González-Acosta, B.; Hernández-Sandoval, F.H.; et al. Morphology, biochemistry, and growth of raphidophyte strains from the Gulf of California. Hydrobiologia 2012, 693, 81–97. [Google Scholar] [CrossRef]
- Haque, S.M.; Onoue, Y. Variation in toxin compositions of two harmful raphidophytes, Chattonella antiqua and Chattonella marina, at diff erent salinities. Environ. Toxicol. 2002, 17, 113–118. [Google Scholar] [CrossRef]
- Khan, S.; Arakawa, O.; Onoue, Y. Neurotoxins in a toxic red tide of Heterosigma akashiwo (Raphidophyceae) in Kagoshima Bay, Japan. Aquac. Res. 1997, 28, 9–14. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.F.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Engesmo, A.; Eikrem, W.; Seoane, S.; Smith, K.; Edvardsen, B.; Hofgaard, A.; Tomas, C.R. New insights into the morphology and phylogeny of Heterosigma akashiwo (Raphidophyceae), with the description of Heterosigma minor sp. nov. Phycologia 2016, 55, 279–294. [Google Scholar] [CrossRef]
- Guillou, L.; Nézan, E.; Cueff, V.; Erard-Le Denn, E.; Cambon-Bonavita, M.-A.; Gentien, P.; Barbier, G. Genetic diversity and molecular detection of three toxic dinoflagellate genera (Alexandrium, Dinophysis, and Karenia) from French coasts. Protist 2002, 153, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Nézan, E.; Siano, R.; Boulben, S.; Six, C.; Bilien, G.; Chèze, K.; Duval, A.; Le Panse, S.; Quéré, J.; Chomérat, N. Genetic diversity of the harmful family Kareniaceae (Gymnodiniales, Dinophyceae) in France, with the description of Karlodinium gentienii sp.: A new potentially toxic dinoflagellate. Harmful Algae 2014, 40, 75–91. [Google Scholar] [CrossRef] [Green Version]
- Haywood, A.J.; Scholin, C.A.; Marin, R., III; Steidinger, K.A.; Heil, C.; Ray, J. Molecular detection of the brevetoxin-producing dinoflagellate Karenia brevis and closely related species using rRNA-targeted probes and a semiautomated sandwich hybridization assay. J. Phycol. 2007, 43, 1271–1286. [Google Scholar] [CrossRef]
- Moestrup, Ø.; Akselmann-Cardella, R.; Churro, C.; Fraga, S.; Hoppenrath, M.; Iwataki, M.; Larsen, J.; Lundholm, N.; Zingone, A. IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. 2009. Available online: http://www.marinespecies.org/hab (accessed on 28 May 2021).
- Khan, S.; Arakawa, O.; Onoue, V.A. Toxicological study of the marine phytoflagellate, Chattonella antiqua (Raphidophyceae). Phycologia 1996, 35, 239–244. [Google Scholar] [CrossRef]
- Keppler, C.J.; Lewitus, A.J.; Ringwood, A.H.; Hoguet, J.; Staton, T. Sublethal cellular effects of short-term raphidophyte and brevetoxin exposures on the eastern oyster Crassostrea virginica. Mar. Ecol. Prog. Ser. 2006, 312, 141–147. [Google Scholar] [CrossRef]
- Horiguchi, T.; Yoshizawa-Ebata, J.; Nakayama, T. Halostylodinium arenarium, gen. et sp. nov. (Dinophyceae), a coccoid sand-dwelling dinoflagellate from subtropical Japan. J. Phycol. 2000, 36, 960–971. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
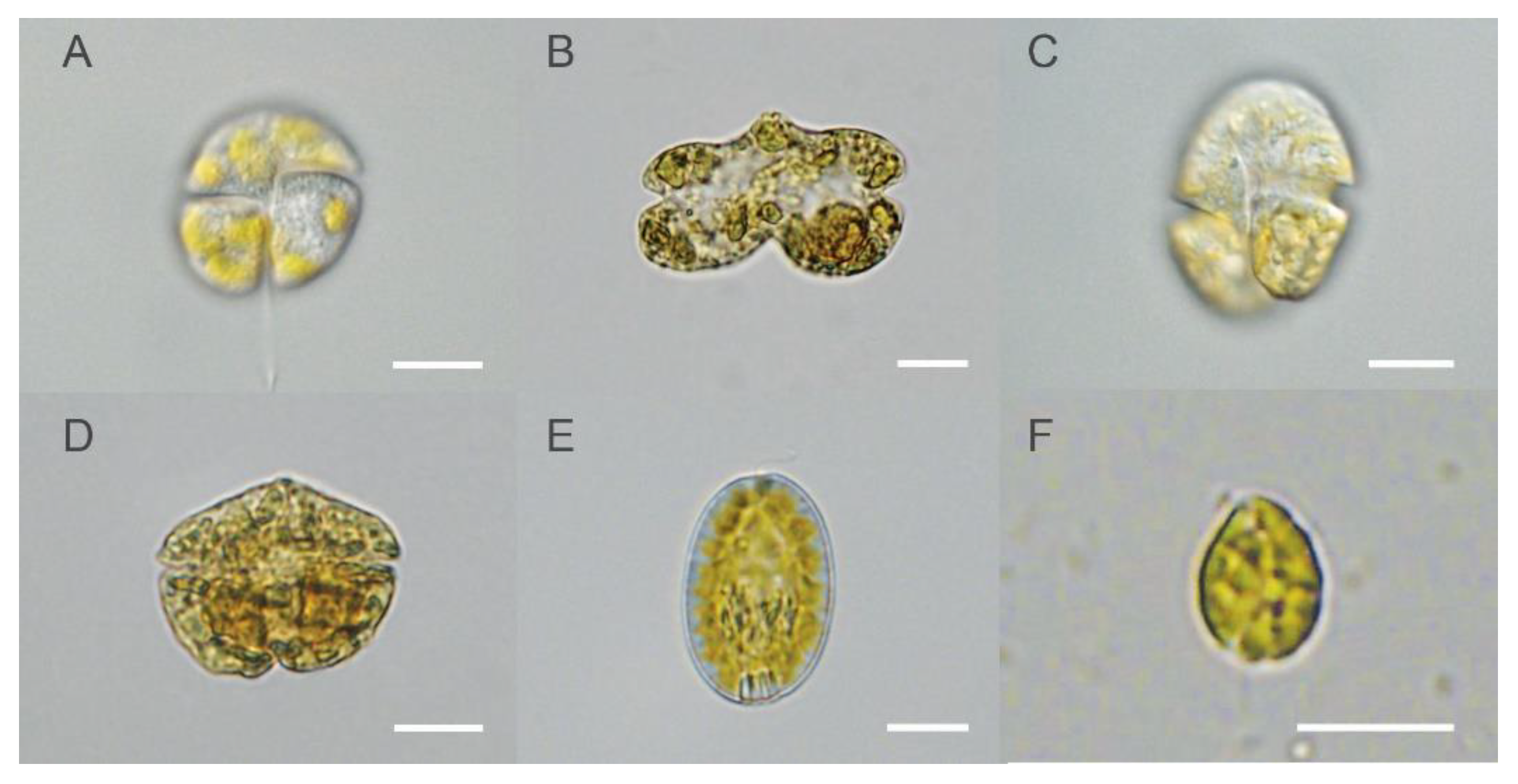{kind=link}
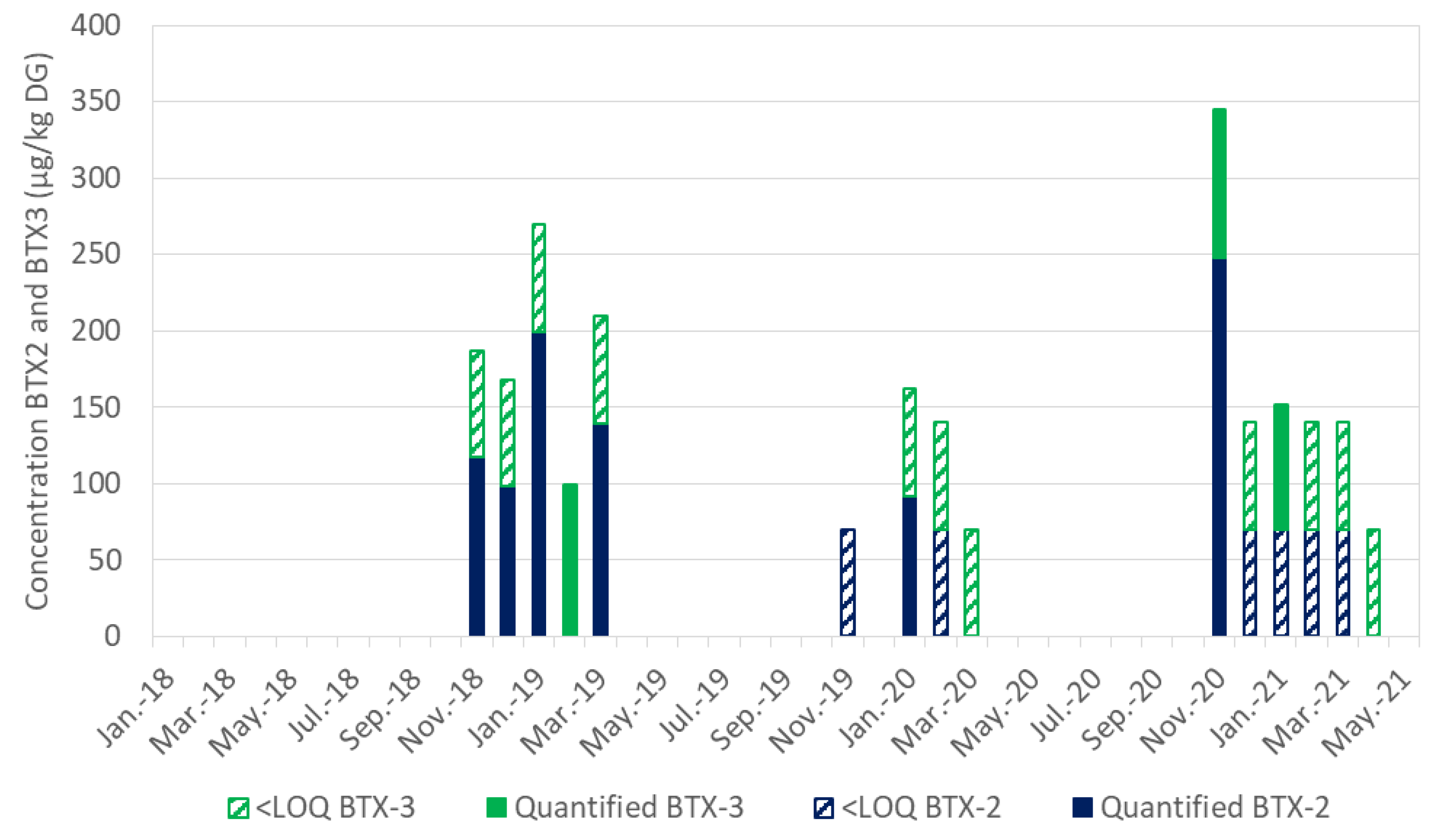
| Month/Year | Targeted BTX Content (µg/kg DG) | ||
|---|---|---|---|
| BTX-2 | BTX-3 | BTX-2 + BTX-3 | |
| Nov-18 | 117.2 | <LOQ * | 117.2 |
| Dec-18 | 98.2 | <LOQ | 98.2 |
| Jan-19 | 199.3 | <LOQ | 199.3 |
| Feb-19 | <LOD ** | 99.2 | 99.2 |
| Mar-19 | 139.5 | <LOQ | 139.5 |
| Nov-19 | <LOQ | <LOD | <LOQ |
| Jan-20 | 92 | <LOQ | 92 |
| Feb-20 | <LOQ | <LOQ | <LOQ |
| Nov-20 | 247.8 | 96.8 | 344.6 |
| Dec-20 | <LOQ | <LOQ | <LOQ |
| Jan-21 | <LOQ | 82 | 82 |
| Feb-21 | <LOQ | <LOQ | <LOQ |
| Mar-21 | <LOQ | <LOQ | <LOQ |
| Apr-21 | <LOD | <LOQ | <LOQ |
| May-21 | <LOD | <LOD | <LOD |
| Transitions | ID | Ion | Dwell (ms) | EP (V) | DP (V) | CE (V) | CXP (V) | |
|---|---|---|---|---|---|---|---|---|
| Q1 Mass (Da) | Q3 Mass (Da) | |||||||
| 912.5 | 319.3 | BTX2 | [M + NH4]+ | 10 | 10 | 81 | 37 | 7 |
| 912.5 | 895.5 | 10 | 10 | 81 | 19 | 22 | ||
| 914.5 | 807.5 | BTX3 | [M + NH4]+ | 10 | 10 | 84 | 24 | 22 |
| 914.5 | 725.5 | 10 | 10 | 84 | 33 | 18 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amzil, Z.; Derrien, A.; Terre Terrillon, A.; Duval, A.; Connes, C.; Marco-Miralles, F.; Nézan, E.; Mertens, K.N. Monitoring the Emergence of Algal Toxins in Shellfish: First Report on Detection of Brevetoxins in French Mediterranean Mussels. Mar. Drugs 2021, 19, 393. https://0-doi-org.brum.beds.ac.uk/10.3390/md19070393
Amzil Z, Derrien A, Terre Terrillon A, Duval A, Connes C, Marco-Miralles F, Nézan E, Mertens KN. Monitoring the Emergence of Algal Toxins in Shellfish: First Report on Detection of Brevetoxins in French Mediterranean Mussels. Marine Drugs. 2021; 19(7):393. https://0-doi-org.brum.beds.ac.uk/10.3390/md19070393
Chicago/Turabian StyleAmzil, Zouher, Amélie Derrien, Aouregan Terre Terrillon, Audrey Duval, Coralie Connes, Françoise Marco-Miralles, Elisabeth Nézan, and Kenneth Neil Mertens. 2021. "Monitoring the Emergence of Algal Toxins in Shellfish: First Report on Detection of Brevetoxins in French Mediterranean Mussels" Marine Drugs 19, no. 7: 393. https://0-doi-org.brum.beds.ac.uk/10.3390/md19070393