
Bacillus amyloliquefaciens-9 Reduces Somatic Cell Count and Modifies Fecal Microbiota in Lactating Goats

 and
and 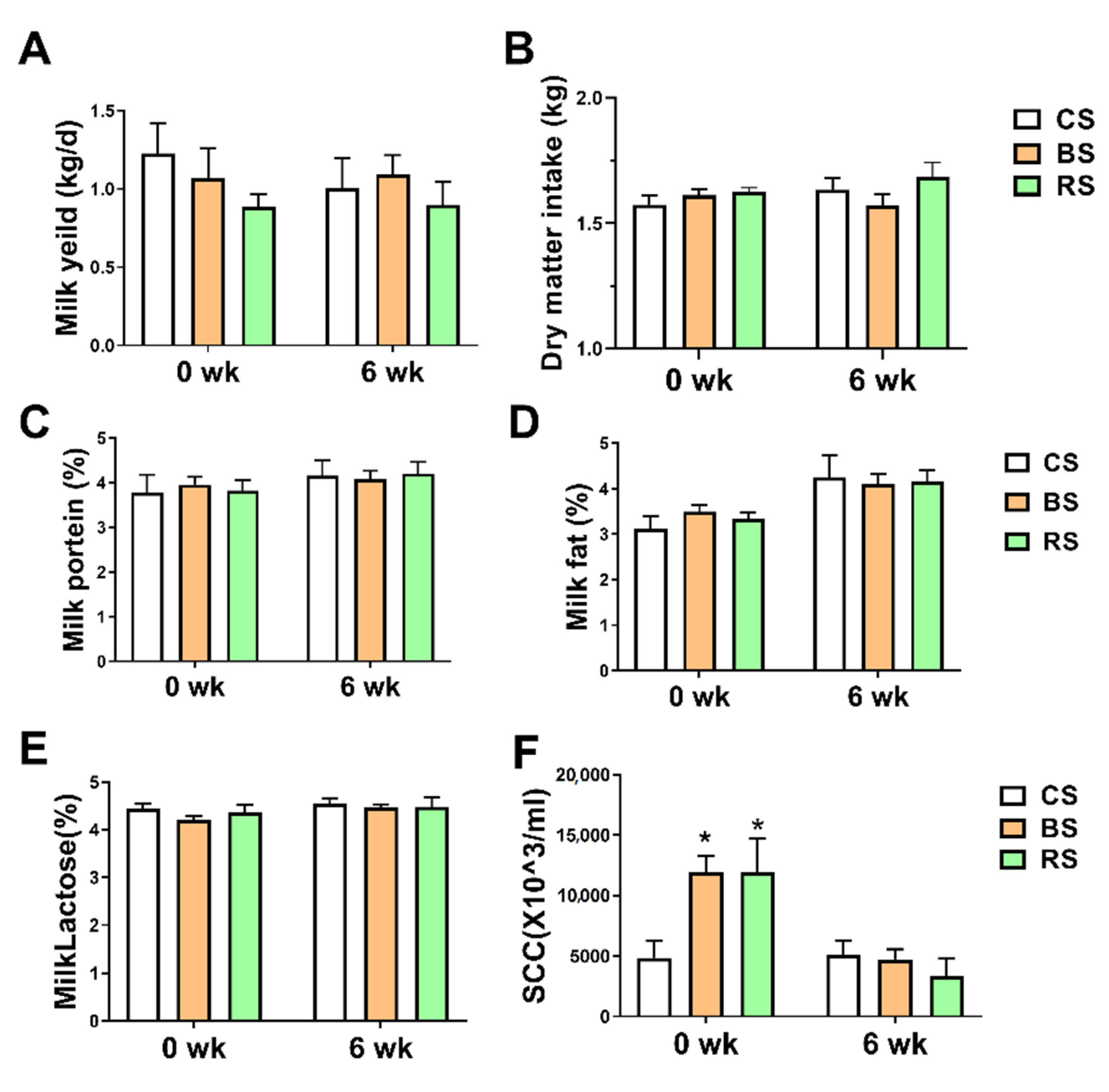{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. GB-9 and RT Supplementation Decreased SCC
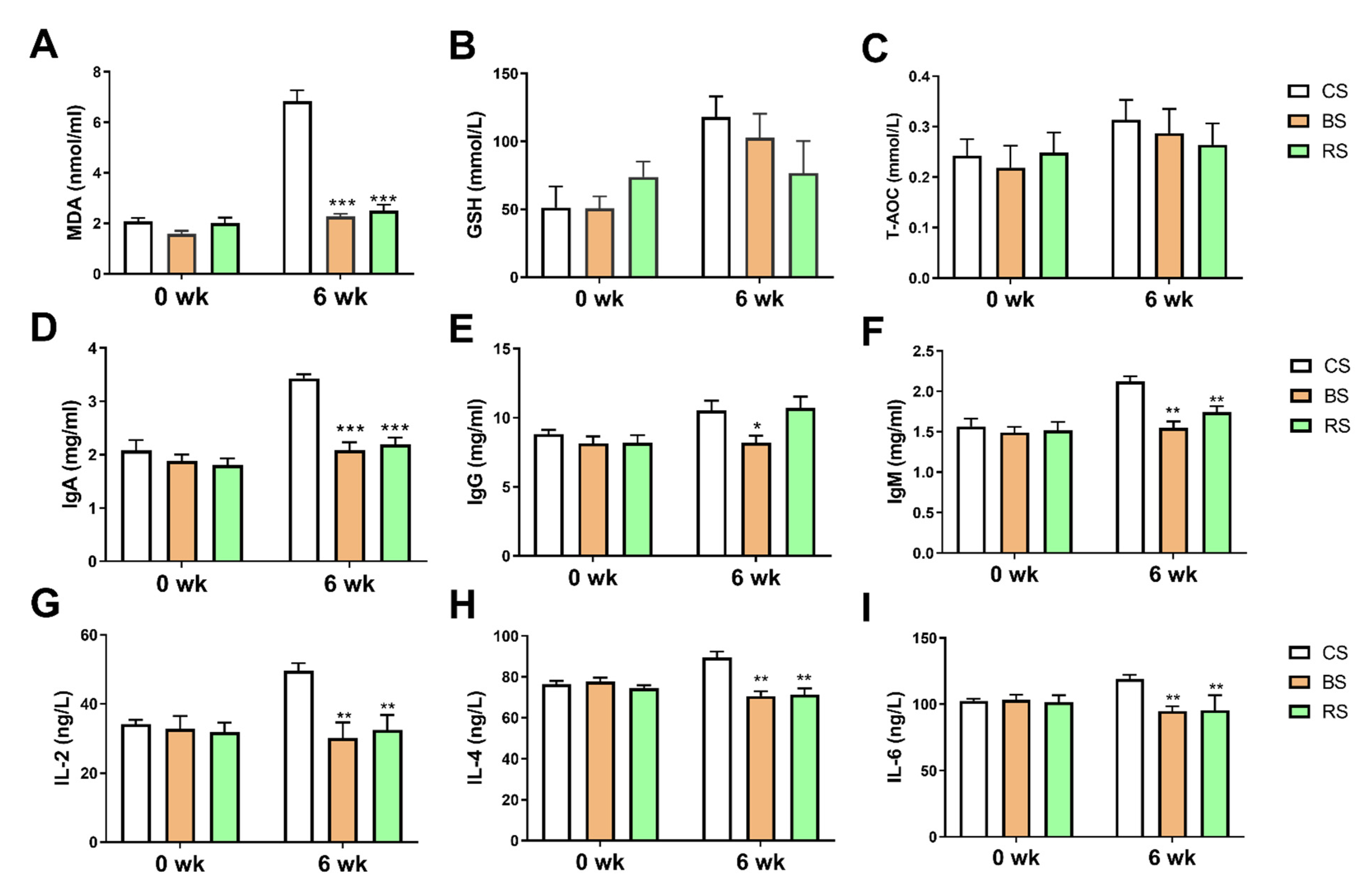
2.2. GB-9 and RT Altered Immune and Oxidative Stress Level in Serum
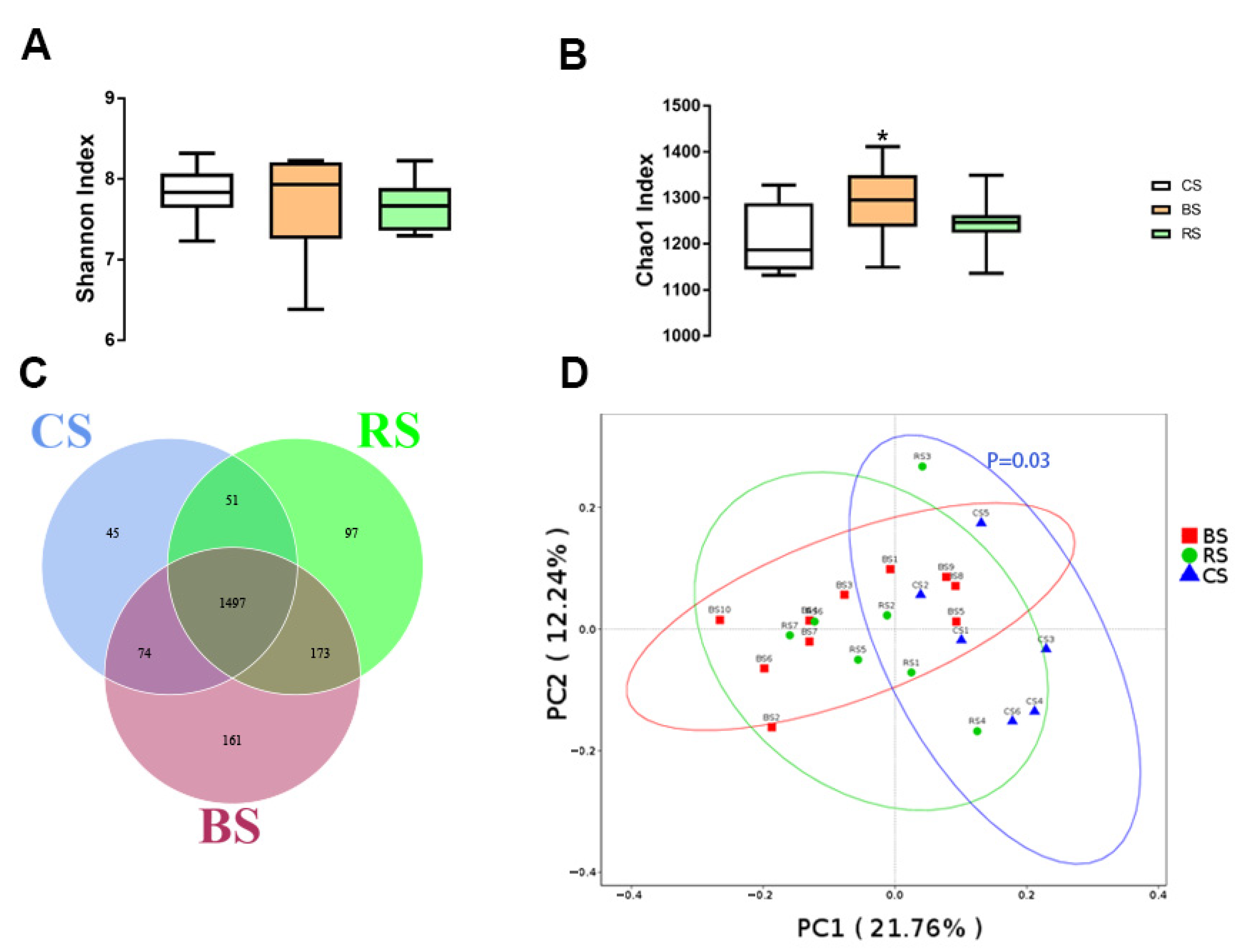
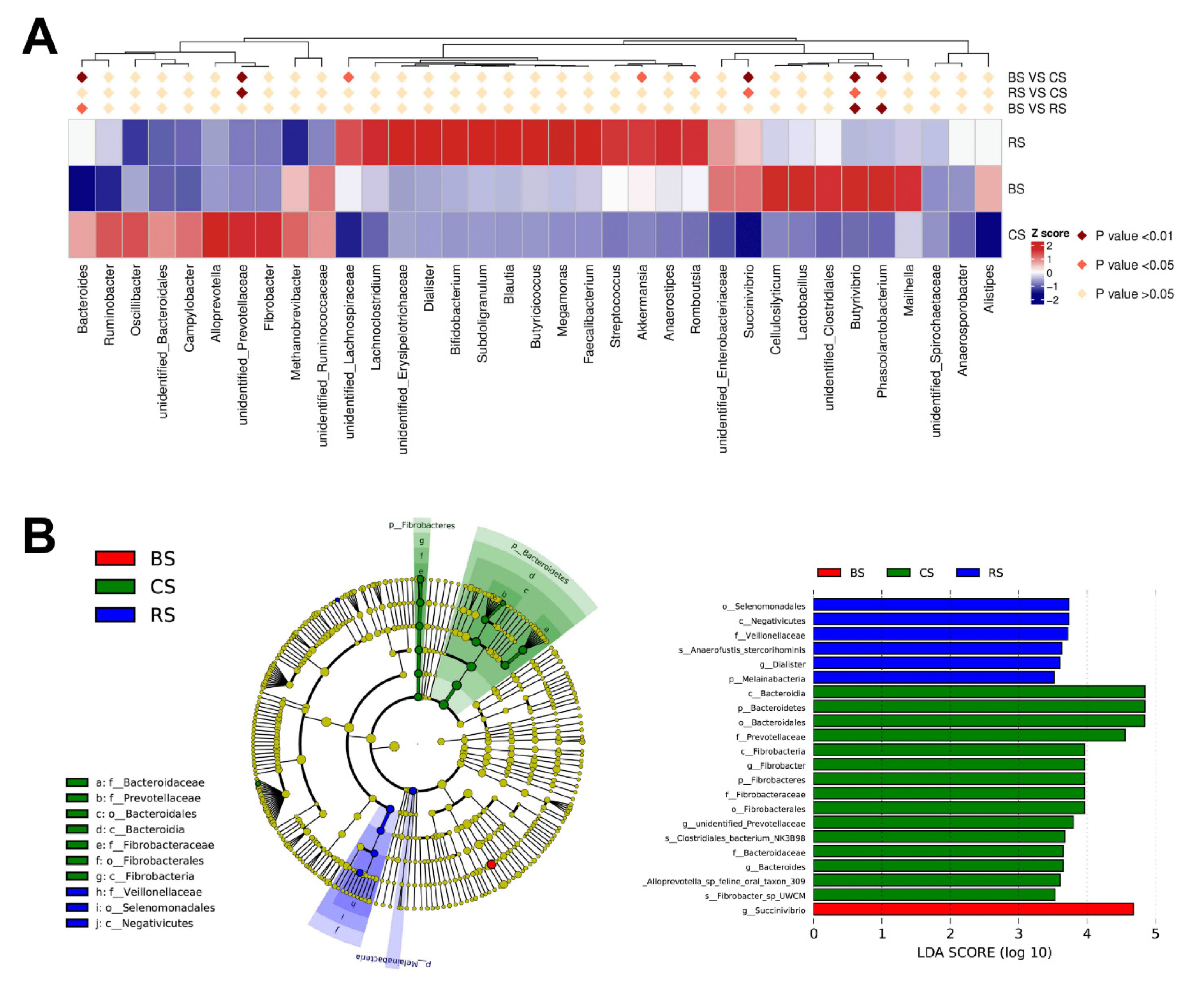
2.3. GB-9 or RT Altered Fecal Microbiota Profiling
2.4. Bacteroides Correlated with the Concertation of Immune Factors
3. Discussion
4. Materials and Methods
4.1. Experiment Design
4.2. Sample Collection and Analysis
4.3. 16S rRNA Gene Sequencing
4.4. Sequencing Data Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halasa, T.; Huijps, K.; Østerås, O.; Hogeveen, H. Economic effects of bovine mastitis and mastitis management: A review. Vet. Q. 2007, 29, 18–31. [Google Scholar] [CrossRef]
- Ma, C.; Sun, Z.; Zeng, B.; Huang, S.; Zhao, J.; Zhang, Y.; Su, X.; Xu, J.; Wei, H.; Zhang, H. Cow-to-mouse fecal transplantations suggest intestinal microbiome as one cause of mastitis. Microbiome 2018, 6, 200. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Xue, M.; Liu, J. Composition of rumen bacterial community in dairy cows with different levels of somatic cell counts. Front. Microbiol. 2018, 9, 3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guterbock, W.M.; Van Eenennaam, A.L.; Anderson, R.J.; Gardner, I.A.; Cullor, J.S.; Holmberg, C.A. Efficacy of intramammary antibiotic therapy for treatment of clinical mastitis caused by environmental pathogens. J. Dairy Sci. 1993, 76, 3437–3444. [Google Scholar] [CrossRef]
- Myllys, V.; Asplund, K.; Brofeldt, E.; Hirvelä-Koski, V.; Honkanen-Buzalski, T.; Junttila, J.; Kulkas, L.; Myllykangas, O.; Niskanen, M.; Saloniemi, H.; et al. Bovine mastitis in Finland in 1988 and 1995-changes in prevalence and antimicrobial resistance. Acta Vet. Scand. 1998, 39, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Pyörälä, S.; Kaartinen, L.; Käck, H.; Rainio, V. Efficacy of two therapy regimens for treatment of experimentally induced Escherichia coli mastitis in cows. J. Dairy Sci. 1994, 77, 453–461. [Google Scholar] [CrossRef]
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Reviews. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Xie, Y.; Zajac, A.M.; Hu, Y.; Aroian, R.V.; Urban, J.F., Jr.; Li, R.W. Gut microbial signatures associated with moxidectin treatment efficacy of Haemonchus contortus in infected goats. Vet. Microbiol. 2020, 242, 108607. [Google Scholar] [CrossRef]
- Gutiérrez-Chávez, A.J.; Martínez-Ortega, E.A.; Valencia-Posadas, M.; León-Galván, M.F.; de la Fuente-Salcido, N.M.; Bideshi, D.K.; Barboza-Corona, J.E. Potential use of Bacillus thuringiensis bacteriocins to control antibiotic-resistant bacteria associated with mastitis in dairy goats. Folia Microbiol. 2016, 61, 11–19. [Google Scholar] [CrossRef]
- Fernández, L.; Arroyo, R.; Espinosa, I.; Marín, M.; Jiménez, E.; Rodríguez, J.M. Probiotics for human lactational mastitis. Benef. Microbes 2014, 5, 169–183. [Google Scholar] [CrossRef]
- Zhang, W.; Xin, H.; Jiang, N.; Lv, Z.; Shu, J.; Shi, H. Bacillus Amyloliquefaciens-9 as an Alternative Approach to Cure Diarrhea in Saanen Kids. Animals 2021, 11, 592. [Google Scholar] [CrossRef]
- Zhang, W.; Wei, L.; Xu, R.; Lin, G.; Xin, H.; Lv, Z.; Qian, H.; Shi, H. Evaluation of the antibacterial material production in the fermentation of Bacillus amyloliquefaciens-9 from Whitespotted Bamboo Shark (Chiloscyllium plagiosum). Mar. Drugs 2020, 18, 119. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Fukuda, M.; Amaki, Y.; Sakaguchi, T.; Inai, K.; Ishihara, A.; Nakajima, H. Importance of prumycin produced by Bacillus amyloliquefaciens SD-32 in biocontrol against cucumber powdery mildew disease. Pest Manag. Sci. 2017, 73, 2419–2428. [Google Scholar] [CrossRef]
- Regmi, S.; Choi, Y.H.; Choi, Y.S.; Kim, M.R.; Yoo, J.C. Antimicrobial peptide isolated from Bacillus amyloliquefaciens K14 revitalizes its use in combinatorial drug therapy. Folia Microbiol. 2017, 62, 127–138. [Google Scholar] [CrossRef]
- Kumar, S.; Haq, I.; Prakash, J.; Singh, S.K.; Mishra, S.; Raj, A. Purification, characterization and thermostability improvement of xylanase from Bacillus amyloliquefaciens and its application in pre-bleaching of kraft pulp. Biotech 2017, 7, 20. [Google Scholar] [CrossRef]
- Fei, H.; Lin, G.D.; Zheng, C.C.; Huang, M.M.; Qian, S.C.; Wu, Z.J.; Sun, C.; Shi, Z.G.; Li, J.Y.; Han, B.N. Effects of Bacillus amyloliquefaciens and Yarrowia lipolytica lipase 2 on immunology and growth performance of Hybrid sturgeon. Fish Shellfish. Immunol. 2018, 82, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Lee, J.S.; Su, X.D.; Li, H.X.; Lee, S.K.; Wang, H.-S.; Kim, Y.H.; Yang, S.Y. Three new constituents from the aerial parts of Tetrastigma hemsleyanum. Phytochem. Lett. 2018, 27, 25–29. [Google Scholar] [CrossRef]
- Ye, C.-L.; Liu, X.-G. Extraction of flavonoids from tetrastigma hemsleyanum diels et Gilg and their antioxidant activity. J. Food Process Pres. 2015, 39, 2197–2205. [Google Scholar] [CrossRef]
- Lu, Y.; Hu, Y.L.; Kong, X.F.; Wang, D.Y. Selection of component drug in activating blood flow and removing blood stasis of Chinese herbal medicinal formula for dairy cow mastitis by hemorheological method. J. Ethnopharmacol. 2008, 116, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tao, L.; Ru, Y.; Weng, S.; Chen, Z.; Wang, J.; Guo, L.; Lin, Z.; Pan, W.; Qiu, B. Antibacterial mechanism of Tetrastigma hemsleyanum Diels et Gilg’s polysaccharides by metabolomics based on HPLC/MS. Int. J. Biol. Macromol. 2019, 140, 206–215. [Google Scholar] [CrossRef]
- Du, S.; Xiang, T.; Song, Y.; Huang, L.; Sun, Y.; Han, Y. Transgenic hairy roots of Tetrastigma hemsleyanum: Induction, propagation, genetic characteristics and medicinal components. Plant Cell Tissue Organ Culture (PCTOC) 2015, 122, 373–382. [Google Scholar] [CrossRef]
- Feng, Z.; Hao, W.; Lin, X.; Fan, D.; Zhou, J. Antitumor activity of total favonoids from tetrastigma hemsleyanum diels et gilg is associated with the inhibition of regulatory T cells in mice. OncoTargets Ther. 2014, 7, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Kritas, S.K.; Govaris, A.; Christodoulopoulos, G.; Burriel, A.R. Effect of Bacillus licheniformis and Bacillus subtilis supplementation of ewe’s feed on sheep milk production and young lamb mortality. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2006, 53, 170–173. [Google Scholar] [CrossRef]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef]
- Barbosa, T.M.; Serra, C.R.; La Ragione, R.M.; Woodward, M.J.; Henriques, A.O. Screening for bacillus isolates in the broiler gastrointestinal tract. Appl. Environ. Microbiol. 2005, 71, 968–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, P.; Wang, J.Q.; Zhang, H.T. Effects of Bacillus subtilis natto on performance and immune function of preweaning calves. J. Dairy Sci. 2010, 93, 5851–5855. [Google Scholar] [CrossRef]
- Luan, S.; Duersteler, M.; Galbraith, E.A.; Cardoso, F.C. Effects of direct-fed Bacillus pumilus 8G-134 on feed intake, milk yield, milk composition, feed conversion, and health condition of pre- and postpartum Holstein cows. J. Dairy Sci. 2015, 98, 6423–6432. [Google Scholar] [CrossRef] [Green Version]
- Sadek, K.; Saleh, E.; Ayoub, M. Selective, reliable blood and milk bio-markers for diagnosing clinical and subclinical bovine mastitis. Trop. Anim. Health Prod. 2017, 49, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Nishi, K.; Hundal, S.S. Chlorpyrifos induced toxicity in reproductive organs of female Wistar rats. Food Chem. Toxicol. 2013, 62, 732–738. [Google Scholar] [CrossRef] [PubMed]
- McKee, M.D.; Fichera, A.; Nishimura, M.I. T cell immunotherapy. Front. Biosci. 2007, 12, 919–932. [Google Scholar] [CrossRef]
- Nickerson, S.C.; Owens, W.E.; Rejman, J.J.; Oliver, S.P. Effects of interleukin-1 and interleukin-2 on mammary gland leukocyte populations and histology during the early nonlactating period. Zentralbl. Vet. B 1993, 40, 621–633. [Google Scholar] [CrossRef]
- Bochniarz, M.; Zdzisińska, B.; Wawron, W.; Szczubiał, M.; Dąbrowski, R. Milk and serum IL-4, IL-6, IL-10, and amyloid A concentrations in cows with subclinical mastitis caused by coagulase-negative staphylococci. J. Dairy Sci. 2017, 100, 9674–9680. [Google Scholar] [CrossRef] [Green Version]
- Osman, K.M.; Hassan, H.M.; Ibrahim, I.M.; Mikhail, M.M. The impact of staphylococcal mastitis on the level of milk IL-6, lysozyme and nitric oxide. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 85–93. [Google Scholar] [CrossRef]
- Sakemi, Y.; Tamura, Y.; Hagiwara, K. Interleukin-6 in quarter milk as a further prediction marker for bovine subclinical mastitis. J. Dairy Res. 2011, 78, 118–121. [Google Scholar] [CrossRef]
- Shi, L.; Zhao, D.; Hou, C.; Peng, Y.; Liu, J.; Zhang, S.; Liu, J.; Long, J. Early interleukin-6 enhances hepatic ketogenesis in APP(SWE)/PSEN1dE9 mice via 3-hydroxy-3-methylglutary-CoA synthase 2 signaling activation by p38/nuclear factor κB p65. Neurobiol. Aging. 2017, 56, 115–126. [Google Scholar] [CrossRef]
- Dan, D.; Bruckmaier, R.M.; Wellnitz, O. Ketoprofen affects the mammary immune response in dairy cows in vivo and in vitro. J. Dairy Sci. 2018, 101, 11321–11329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Xue, M.-Y.; Sun, H.-Z.; Valencak, T.G.; Guan, L.L.; Liu, J. Rumen and hindgut bacteria are potential indicators for mastitis of mid-lactating holstein dairy cows. Microorganisms 2020, 8, 2042. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lv, M.; Wang, J.; Tian, Z.; Yu, B.; Wang, B.; Liu, J.; Liu, H. Dandelion (Taraxacum mongolicum Hand.-Mazz.) Supplementation-Enhanced Rumen Fermentation through the Interaction between Ruminal Microbiome and Metabolome. Microorganisms 2021, 9, 83. [Google Scholar] [CrossRef] [PubMed]
- Mi, L.; Yang, B.; Hu, X.; Luo, Y.; Liu, J.; Yu, Z.; Wang, J. Comparative Analysis of the Microbiota Between Sheep Rumen and Rabbit Cecum Provides New Insight Into Their Differential Methane Production. Front. Microbiol. 2018, 9, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Le, J.; Wu, P.; Liu, J.; Guan, L.L.; Wang, J. Alfalfa Intervention Alters Rumen Microbial Community Development in Hu Lambs During Early Life. Front. Microbiol. 2018, 9, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Ji, Y.; Yin, C.; Deng, M.; Tang, T.; Deng, B.; Ren, W.; Deng, J.; Yin, Y.; Tan, C. Differential analysis of gut microbiota correlated with oxidative stress in sows with high or low litter performance during lactation. Front. Microbiol. 2018, 9, 1665. [Google Scholar] [CrossRef] [PubMed]
- Pumbwe, L.; Skilbeck, C.A.; Wexler, H.M. Impact of anatomic site on growth, efflux-pump expression, cell structure, and stress responsiveness of Bacteroides fragilis. Curr Microbiol. 2007, 55, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Salyers, A.A. Bacteroides of the human lower intestinal tract. Annu. Rev. Microbiol. 1984, 38, 293–313. [Google Scholar] [CrossRef] [PubMed]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.T.T.; Cousin, F.J.; Lynch, D.B.; Menon, R.; Brulc, J.; Brown, J.R.; O’Herlihy, E.; Butto, L.F.; Power, K.; Jeffery, I.B.; et al. Prebiotic supplementation in frail older people affects specific gut microbiota taxa but not global diversity. Microbiome 2019, 7, 39. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Kasper, D.L. The love-hate relationship between bacterial polysaccharides and the host immune system. Nat. Rev. Immunol. 2006, 6, 849–858. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Xu, G.; Jin, Y.; Sun, C.; Zhou, L.; Lin, G.; Xu, R.; Wei, L.; Fei, H.; Wang, D.; et al. Isolation and characterization of Bacillus sp. GFP-2, a novel Bacillus strain with antimicrobial activities, from Whitespotted bamboo shark intestine. AMB Express 2018, 8, 84. [Google Scholar] [CrossRef]
- Cai, J.; Zhao, F.Q.; Liu, J.X.; Wang, D.M. Local mammary glucose supply regulates availability and intracellular metabolic pathways of glucose in the mammary gland of lactating dairy goats under malnutrition of energy. Front. Physiol. 2018, 9, 1467. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Jiang, N.; Zhang, W.; Lv, Z.; Liu, J.; Shi, H. Bacillus amyloliquefaciens-9 Reduces Somatic Cell Count and Modifies Fecal Microbiota in Lactating Goats. Mar. Drugs 2021, 19, 404. https://0-doi-org.brum.beds.ac.uk/10.3390/md19080404
Li Y, Jiang N, Zhang W, Lv Z, Liu J, Shi H. Bacillus amyloliquefaciens-9 Reduces Somatic Cell Count and Modifies Fecal Microbiota in Lactating Goats. Marine Drugs. 2021; 19(8):404. https://0-doi-org.brum.beds.ac.uk/10.3390/md19080404
Chicago/Turabian StyleLi, Yongtao, Nannan Jiang, Wenying Zhang, Zhengbing Lv, Jianxin Liu, and Hengbo Shi. 2021. "Bacillus amyloliquefaciens-9 Reduces Somatic Cell Count and Modifies Fecal Microbiota in Lactating Goats" Marine Drugs 19, no. 8: 404. https://0-doi-org.brum.beds.ac.uk/10.3390/md19080404