
Identification of New CTX Analogues in Fish from the Madeira and Selvagens Archipelagos by Neuro-2a CBA and LC-HRMS

 , , ,
, , ,  , , ,
, , ,
Abstract
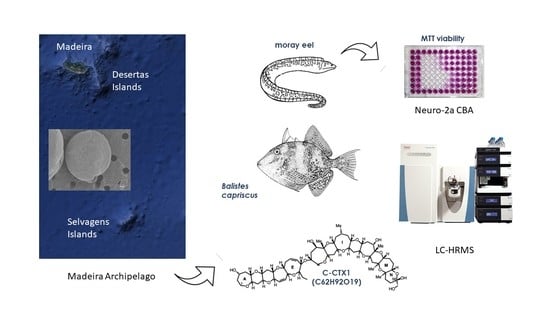
:
1. Introduction
2. Results
2.1. Identification of Fish by COI Sequencing
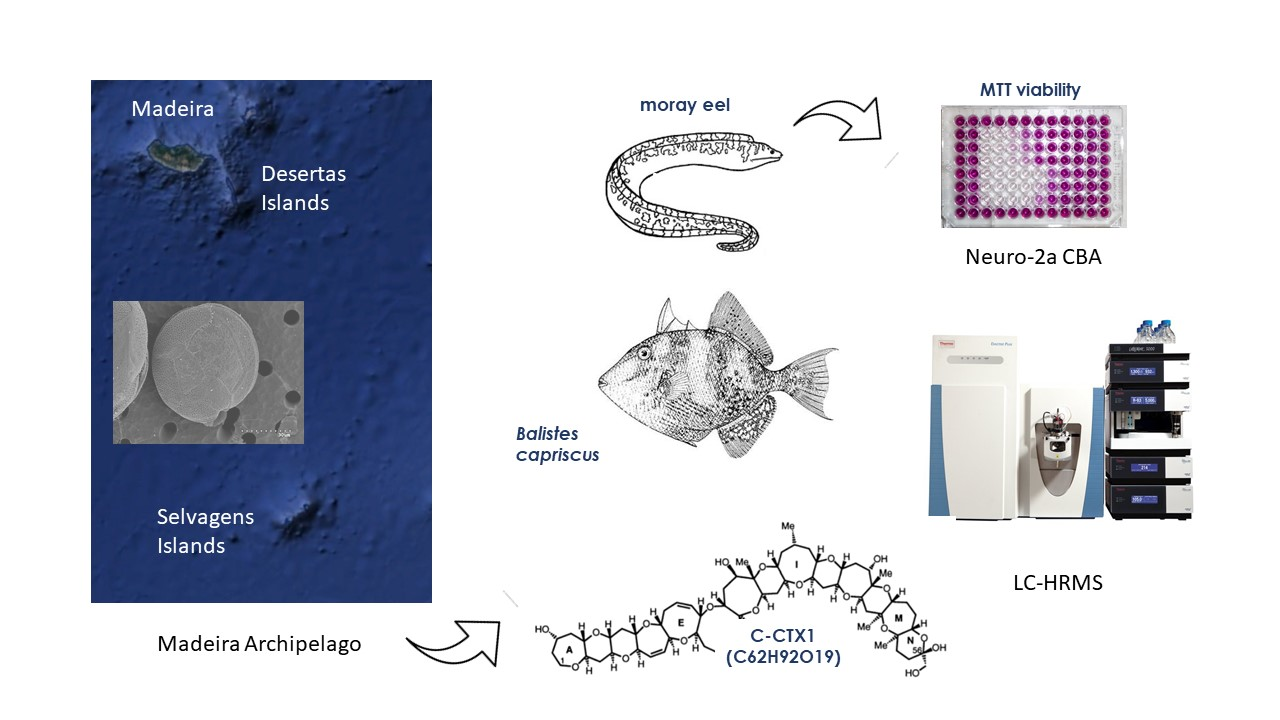
2.2. CTX-like Toxicity Evaluation by Neuro-2a CBA
2.3. Identification of CTXs by LC-HRMS Analysis
3. Discussion
3.1. Identification of Fish by COI Sequencing
3.2. Toxin Analysis by CBA and LC-HRMS Analysis
4. Materials and Methods
4.1. Reagents and Equipment
4.2. Fish Samples
4.3. Identification of Fish by COI Sequencing
4.4. Extraction Protocols
- Protocol-A
- Protocol-B
4.5. CTX-like Toxicity Evaluation by Neuro-2a CBA
4.6. LC-HRMS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinain, M.; Darius, H.T.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and toxin production in the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae) in culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.J.; Lewis, R.J.; Poli, M.A.; Gillespie, N.C. Strain dependent production of ciguatoxin precursors (gambiertoxins) by Gambierdiscus toxicus (Dinophyceae) in culture. Toxicon 1991, 29, 761–775. [Google Scholar] [CrossRef]
- Litaker, R.W.; Holland, W.C.; Hardison, D.R.; Pisapia, F.; Hess, P.; Kibler, S.R.; Tester, P.A. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, 12, e185776. [Google Scholar] [CrossRef]
- Roeder, K.; Erler, K.; Kibler, S.; Tester, P.; Ho Van, T.; Lam, N.-N.; Gerdts, G.; Luckas, B. Characteristic profiles of Ciguatera toxins in different strains of Gambierdiscus spp. Toxicon 2010, 56, 731–738. [Google Scholar] [CrossRef]
- Laza-Martínez, A.; David, H.; Riobó, P.; Miguel, I.; Orive, E. Characterization of a Strain of Fukuyoa paulensis (Dinophyceae) from the Western Mediterranean Sea. J. Eukaryot. Microbiol. 2016, 63, 481–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R.J.; Holmes, M.J. Origin and transfer of toxins involved in ciguatera. Comp. Biochem. Physiol. C Comp. Pharmacol. Toxicol. 1993, 106, 615–628. [Google Scholar] [CrossRef]
- Lewis, R.J.; Vernoux, J.-P.; Brereton, M.I. Structure of Caribbean ciguatoxin isolated from Caranx latus. J. Am. Chem. Soc. 1998, 120, 5914–5920. [Google Scholar] [CrossRef]
- Bagnis, R.; Kuberski, T.; Laugier, S. Clinical observations on 3009 cases of ciguatera (fish poisoning) in the South -Pacific. Am. J. Trop. Med. Hyg. 1979, 28, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Pearn, J. Neurology of ciguatera. J. Neurol. Neurosurg. Psychiatry 2001, 70, 4–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Haro, L.; Pommier, P.; Valli, M. Emergence of imported ciguatera in Europe: Report of 18 cases at the Poison Control Centre of Marseille. J. Toxicol. Clin. Toxicol. 2003, 41, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Arbor, K.; Goldstein, R.A.; Bayer, M.; McKay, C.A. Recurrent ciguatera symptoms nine years after initial poisoning. Clin. Toxicol. 2005, 43, 6. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.J. Ion channel toxins and therapeutics: From cone snail venoms to ciguatera. Ther. Drug. Monit. 2000, 22, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An Updated Review of Ciguatera Fish Poisoning: Clinical, Epidemiological, Environmental, and Public Health Management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.E.; Baden, D.G.; Bean, J.A.; Weisman, R.; Blythe, D.G. Marine Seafood Toxin Diseases: Issues in Epidemiology & Community Outreach BT—Harmful Algae; Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1998; pp. 245–248. [Google Scholar]
- Skinner, M.P.; Brewer, T.D.; Johnstone, R.; Fleming, L.E.; Lewis, R.J. Ciguatera Fish Poisoning in the Pacific Islands (1998 to 2008). PLoS Negl. Trop. Dis. 2011, 5, e1416. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Lewis, R.J. Ciguatoxins: Cyclic polyether modulators of voltage-gated Iion channel function. Mar. Drugs 2006, 4, 82–118. [Google Scholar] [CrossRef] [Green Version]
- Abraham, A.; Jester, E.L.E.; Granade, H.R.; Plakas, S.M.; Dickey, R.W. Caribbean ciguatoxin profile in raw and cooked fish implicated in ciguatera. Food. Chem. 2012, 131, 192–198. [Google Scholar] [CrossRef]
- Tester, P.A.; Feldman, R.L.; Nau, A.W.; Kibler, S.R.; Litaker, R.W. Ciguatera fish poisoning and sea surface temperatures in the Caribbean Sea and the West Indies. Toxicon 2010, 56, 698–710. [Google Scholar] [CrossRef]
- Chinain, M.; Gatti, C.M.; Roue, M.; Darius, H.T. Ciguatera poisoning in French Polynesia: Insights into the novel trends of an ancient disease. New Microbes New Infect. 2019, 31, 100565. [Google Scholar] [CrossRef]
- Chan, T.Y.K. Regional Variations in the Risk and Severity of Ciguatera Caused by Eating Moray Eels. Toxins 2017, 9, 201. [Google Scholar] [CrossRef] [Green Version]
- Chan, T.Y.K. Ciguatera Fish Poisoning in East Asia and Southeast Asia. Mar. Drugs 2015, 13, 3466–3478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R.J. Ciguatera: Australian perspectives on a global problem. Toxicon 2006, 48, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Chinain, M.; Gatti, C.M.I.; Ung, A.; Cruchet, P.; Revel, T.; Viallon, J.; Sibat, M.; Varney, P.; Laurent, V.; Hess, P.; et al. Evidence for the Range Expansion of Ciguatera in French Polynesia: A Revisit of the 2009 Mass-Poisoning Outbreak in Rapa Island (Australes Archipelago). Toxins 2020, 12, 759. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H.; Edwards, A.; Zammit, A. Four recent ciguatera fish poisoning incidents in New South Wales, Australia linked to imported fish. Commun. Dis. Intell. 2019, 43, 1–9. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations & World Health Organization. Report of the Expert Meeting on Ciguatera Poisoning: Rome, 19–23 November 2018; World Health Organization: Geneva, Switzerland, 2020; Available online: https://apps.who.int/iris/handle/10665/332640 (accessed on 28 February 2022).
- Hoppenrath, M.; Kretzschmar, A.L.; Kaufmann, M.J.; Murray, S.A. Morphological and molecular phylogenetic identification and record verification of Gambierdiscus excentricus (Dinophyceae) from Madeira Island (NE Atlantic Ocean). Mar. Biodivers. Rec. 2019, 12, 16. [Google Scholar] [CrossRef] [Green Version]
- Reverté, L.; Toldrà, A.; Andree, K.B.; Fraga, S.; de Falco, G.; Campàs, M.; Diogène, J. Assessment of cytotoxicity in ten strains of Gambierdiscus australes from Macaronesian Islands by neuro-2a cell-based assays. J. Appl. Phycol. 2018, 30, 2447–2461. [Google Scholar] [CrossRef]
- Fraga, S.; Rodríguez, F.; Caillaud, A.; Diogène, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp. nov. (Dinophyceae), a benthic toxic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, F.; Fraga, S.; Ramilo, I.; Rial, P.; Figueroa, I.R.; Riobó, P.; Bravo, I. The Canary Islands (NE Atlantic) as a biodiversity hotspot of Gambierdiscus: Implications for future trends of ciguatera in the area. Harmful Algae 2017, 68, 131–143. [Google Scholar] [CrossRef]
- Boente-Juncal, A.; Vale, C.; Alfonso, A.; Botana, L.M. Synergistic Effect of Transient Receptor Potential Antagonist and Amiloride against Maitotoxin Induced Calcium Increase and Cytotoxicity in Human Neuronal Stem Cells. ACS Chem. Neurosci. 2018, 9, 2667–2678. [Google Scholar] [CrossRef]
- Tudó, À.; Gaiani, G.; Rey Varela, M.; Tsumuraya, T.; Andree, K.B.; Fernández-Tejedor, M.; Campàs, M.; Diogène, J. Further Advance of Gambierdiscus Species in the Canary Islands, with the First Report of Gambierdiscus belizeanus. Toxins 2020, 12, 692. [Google Scholar] [CrossRef]
- Rossignoli, A.E.; Tudó, A.; Bravo, I.; Díaz, P.A.; Diogène, J.; Riobó, P. Toxicity Characterisation of Gambierdiscus Species from the Canary Islands. Toxins 2020, 12, 134. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Arellano, J.L.; Luzardo, O.P.; Perez Brito, A.; Hernandez Cabrera, M.; Zumbado, M.; Carranza, C.; Àngel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera fish poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Henao, A.J.; García-Álvarez, N.; Fernández, A.; Saavedra, P.; Silva Sergent, F.; Padilla, D.; Acosta-Hernández, B.; Martel Suárez, M.; Diogène, J.; Real, F. Predictive score and probability of CTX-like toxicity in fish samples from the official control of ciguatera in the Canary Islands. Sci. Total Environ. 2019, 673, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Caillaud, A.; Cañete, E.; de la Iglesia, P.; Gímenez, G.; Diogène, J. Cell-based assay coupled with chromatographic fractioning: A strategy for marine toxins detection in natural samples. Toxicol. In Vitro 2009, 23, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Caillaud, A.; Eixarch, H.; de la Iglesia, P.; Rodríguez, M.; Domínguez, L.; Andree, K.B.; Diogène, J. Towards the standardisation of the neuroblastoma (neuro-2a) cell-based assay for ciguatoxin-like toxicity detection in fish: Application to fish caught in the Canary Islands. Food Addit. Contam. Part A Chem. Anal Control Expo. Risk Assess 2012, 29, 1000–1010. [Google Scholar] [CrossRef]
- Dechraoui, M.Y.B.; Wang, Z.; Turquet, J.; Chinain, M.; Darius, T.; Cruchet, P.; Radwan, F.F.Y.; Dickey, R.W.; Ramsdell, J.S. Biomonitoring of ciguatoxin exposure in mice using blood collection cards. Toxicon 2005, 46, 243–251. [Google Scholar] [CrossRef]
- LePage, K.T.; Dickey, R.W.; Gerwick, W.H.; Jester, E.L.; Murray, T.F. On the use of neuro-2a neuroblastoma cells versus intact neurons in primary culture for neurotoxicity studies. Crit. Rev. Neurobiol. 2005, 17, 27–50. [Google Scholar] [CrossRef]
- Castro, D.; Manger, R.; Vilarino, O.; Gago-Martínez, A. Evaluation of Matrix Issues in the Applicability of the Neuro-2a Cell Based Assay on the Detection of CTX in Fish Samples. Toxins 2020, 12, 308. [Google Scholar] [CrossRef]
- Pawlowiez, R.; Darius, H.T.; Cruchet, P.; Rossi, F.; Caillaud, A.; Laurent, D.; Chinain, M. Evaluation of seafood toxicity in the Australes archipelago (French Polynesia) using the neuroblastoma cell-based assay. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2013, 30, 567–586. [Google Scholar] [CrossRef]
- FDA. Fish and Fishery Products Hazards and Controls Guidance, 4th ed.; IFAS: Gainesville, FL, USA, 2021. Available online: https://www.fda.gov/media/80637/download (accessed on 28 February 2022).
- EFSA Panel on Contaminants in the Food Chain. Scientific Opinion on marine biotoxins in shellfish—Emerging toxins: Ciguatoxin group. EFSA J. 2010, 8, 1627. [Google Scholar] [CrossRef]
- Varela Martínez, C.; León Gómez, I.; Martínez Sánchez, E.V.; Carmona Alférez, R.; Nuñez Gallo, D.; Friedemann, M.; Oleastro, M.; Boziaris, I. Incidence and epidemiological characteristics of ciguatera cases in Europe. EFSA Supporting Publ. 2021, 18, 6650E. [Google Scholar] [CrossRef]
- Otero, P.; Pérez, S.; Alfonso, A.; Vale, C.; Rodríguez, P.; Gouveia, N.N.; Gouveia, N.; Delgado, J.; Vale, P.; Hirama, M.; et al. First toxin profile of ciguateric fish in Madeira Arquipelago (Europe). Anal. Chem. 2010, 82, 6032–6039. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.R.; Estévez, P.; Castro, D.; Soliño, L.; Gouveia, N.; Santos, C.; Rodrigues, S.M.; Leao, J.M.; Gago-Martínez, A. New Insights into the Occurrence and Toxin Profile of Ciguatoxins in the Selvagens Islands (Madeira, Portugal). Toxins 2018, 10, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estévez, P.; Castro, D.; Manuel Leao, J.; Yasumoto, T.; Dickey, R.; Gago-Martínez, A. Implementation of liquid chromatography tandem mass spectrometry for the analysis of ciguatera fish poisoning in contaminated fish samples from Atlantic coasts. Food Chem. 2019, 280, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Estévez, P.; Castro, D.; Pequeno-Valtierra, A.; Leao, J.M.; Vilarino, O.; Diogène, J.; Gago-Martínez, A. An Attempt to Characterize the Ciguatoxin Profile in Seriola fasciata Causing Ciguatera Fish Poisoning in Macaronesia. Toxins 2019, 11, 221. [Google Scholar] [CrossRef] [Green Version]
- Canals, A.; Martínez, C.V.; Diogène, J.; Gago-Martínez, A.; Cebadera-Miranda, L.; de Vasconcelos, F.M.; Gómez, I.L.; Sánchez, E.V.M.; Alférez, R.C.; Núñez, D.; et al. Risk characterisation of ciguatera poisoning in Europe. EFSA Supporting Publ. 2021, 18, 6647E. [Google Scholar] [CrossRef]
- Estévez, P.; Sibat, M.; Leao-Martins, J.M.; Reis Costa, P.; Gago-Martínez, A.; Hess, P. Liquid Chromatography Coupled to High-Resolution Mass Spectrometry for the Confirmation of Caribbean Ciguatoxin-1 as the Main Toxin Responsible for Ciguatera Poisoning Caused by Fish from European Atlantic Coasts. Toxins 2020, 12, 267. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Rodriguez, I.; Barreiro, A.; Kaufmann, M.; Isabel Neto, A.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. First Report of Ciguatoxins in Two Starfish Species: Ophidiaster ophidianus and Marthasterias glacialis. Toxins 2015, 7, 3740–3757. [Google Scholar] [CrossRef] [Green Version]
- Caillaud, A.; de la Iglesia, P.; Darius, H.T.; Pauillac, S.; Aligizaki, K.; Fraga, S.; Chinain, M.; Diogène, J. Update on Methodologies Available for Ciguatoxin Determination: Perspectives to Confront the Onset of Ciguatera Fish Poisoning in Europe. Mar. Drugs 2010, 8, 1838–1907. [Google Scholar] [CrossRef]
- Kochzius, M.; Seidel, C.; Antoniou, A.; Botla, S.K.; Campo, D.; Cariani, A.; Vazquez, E.G.; Hauschild, J.; Hervet, C.; Hjörleifsdottir, S.; et al. Identifying Fishes through DNA Barcodes and Microarrays. PLoS ONE 2010, 5, e12620. [Google Scholar] [CrossRef] [Green Version]
- Soliño, L.; Widgy, S.; Pautonnier, A.; Turquet, J.; Loeffler, C.R.; Flores Quintana, H.A.; Diogène, J. Prevalence of ciguatoxins in lionfish (Pterois spp.) from Guadeloupe, Saint Martin, and Saint Barthélmy Islands (Caribbean). Toxicon 2015, 102, 62–68. [Google Scholar] [CrossRef]
- Chan, W.H.; Mak, Y.L.; Wu, J.J.; Jin, L.; Sit, W.H.; Lam, J.C.W.; de Mitcheson, Y.S.; Chan, L.L.; Lam, P.K.S.; Murphy, M.B. Spatial distribution of ciguateric fish in the Republic of Kiribati. Chemosphere 2011, 84, 117–123. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Vernoux, J.P.; Jones, A.; Lewis, R.J. Isolation and characterisation of Indian Ocean ciguatoxin. Toxicon 2002, 40, 685–693. [Google Scholar] [CrossRef]
- Oshiro, N.; Yogi, K.; Asato, S.; Sasaki, T.; Tamanaha, K.; Hirama, M.; Yasumoto, T.; Inafuku, Y. Ciguatera incidence and fish toxicity in Okinawa, Japan. Toxicon 2010, 56, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Pottier, I.; Hamilton, B.; Jones, A.; Lewis, R.J.; Vernoux, J.P. Identitication of slow and fast-acting toxins in a highly ciguatoxic barracuda (Sphyraena barracuda) by HPLC/MS and radiolabelled ligand binding. Toxicon 2003, 42, 663–672. [Google Scholar] [CrossRef]
- Sánchez-Henao, A.; García-Álvarez, N.; Silva Sergent, F.; Estévez, P.; Gago-Martínez, A.; Martín, F.; Ramos-Sosa, M.; Fernández, A.; Diogène, J.; Real, F. Presence of CTXs in moray eels and dusky groupers in the marine environment of the Canary Islands. Aquat. Toxicol. 2020, 221, 105427. [Google Scholar] [CrossRef]
- Costa, P.R.; Estévez, P.; Soliño, L.; Castro, D.; Rodrigues, S.M.; Timoteo, V.; Leao-Martins, J.M.; Santos, C.; Gouveia, N.; Diogène, J.; et al. An Update on Ciguatoxins and CTX-like Toxicity in Fish from Different Trophic Levels of the Selvagens Islands (NE Atlantic, Madeira, Portugal). Toxins 2021, 13, 580. [Google Scholar] [CrossRef]
- Bagnis, R.; Berglund, F.; Elias, P.S.; van Esch, G.J.; Halstead, B.W.; Kojima, K. Problems of toxicants in marine food products. 1. Marine biotoxins. Bull. World Health Organ. 1970, 42, 69–88. [Google Scholar] [PubMed]
- Gaboriau, M.; Ponton, D.; Darius, H.T.; Chinain, M. Ciguatera fish toxicity in French Polynesia: Size does not always matter. Toxicon 2014, 84, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Bennett, S.; Wernberg, T.; Harvey, E.S.; Santana-Garcon, J.; Saunders, B.J. Tropical herbivores provide resilience to a climate-mediated phase shift on temperate reefs. Ecol. Lett. 2015, 18, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Vergés, A.; Bennett, S.; Bellwood, D.R. Diversity among macroalgae-consuming fishes on coral reefs: A transcontinental comparison. PLoS ONE 2012, 7, e45543. [Google Scholar] [CrossRef]
- Diogène, J.; Rambla, M.; Campàs, M.; Fernández, M.; Andree, K.; Tudó, A.; Rey, M.; Sagristà, N.; Aguayo, P.; Leonardo, S.; et al. Evaluation of ciguatoxins in seafood and the environment in Europe. EFSA Supporting Publ. 2021, 18, 6648E. [Google Scholar] [CrossRef]
- Regulation (EC) No. 2019/624 of the European Parliament and of the Council of 8 February 2019 concerning specific rules for the performance of official controls on the production of meat and for production and relaying areas of live bivalve molluscs. Off. J. Eur. Union 2019, 131, 97.
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Yogi, K.; Oshiro, N.; Inafuku, Y.; Hirama, M.; Yasumoto, T. Regional and Species Characteristics in Fish and Causative Alga from the Pacific. Anal. Chem. 2011, 83, 8886–8891. [Google Scholar] [CrossRef]
- Lewis, R.J.; Inserra, M.; Vetter, I.; Holland, W.C.; Hardison, D.R.; Tester, P.A.; Litaker, R.W. Rapid Extraction and Identification of Maitotoxin and Ciguatoxin-Like Toxins from Caribbean and Pacific Gambierdiscus Using a New Functional Bioassay. PLoS ONE 2016, 11, e160006. [Google Scholar] [CrossRef] [PubMed]
- Dickey, R.W.; Plakas, S.M. Ciguatera: A public health perspective. Toxicon 2010, 56, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Torigoe, K.; Satake, M.; Murata, M.; Yasumoto, T.; Hirota, H. Gambieric Acids: Unprecedented Potent Antifungal Substances Isolated from Cultures of a Marine Dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1992, 114, 1102–1103. [Google Scholar] [CrossRef]
- Inoue, M.; Hirama, M.; Satake, M.; Sugiyama, K.; Yasumoto, T. Inhibition of brevetoxin binding to the voltage-gated sodium channel by gambierol and gambieric acid-A. Toxicon 2003, 41, 469–474. [Google Scholar] [CrossRef]
- Diogène, J.; Reverté, L.; Rambla-Alegre, M.; del Río, V.; de la Iglesia, P.; Campàs, M.; Palacios, O.; Flores, C.; Caixach, J.; Ralijaona, C.; et al. Identification of ciguatoxins in a shark involved in a fatal food poisoning in the Indian Ocean. Sci. Rep. 2017, 7, 8240. [Google Scholar] [CrossRef]
- Boada, L.D.; Zumbado, M.; Luzardo, O.P.; Almeida-González, M.; Plakas, S.M.; Granade, H.R.; Abraham, A.; Jester, E.L.; Dickey, R. Ciguatera fish poisoning on the West Africa Coast: An emerging risk in the Canary Islands (Spain). Toxicon 2010, 56, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerforg, J.M.; Wekell, M.M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: Semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Caillaud, A.; Yasumoto, T.; Diogene, J. Detection and quantification of maitotoxin-like compounds using a neuroblastoma (Neuro-2a) cell based assay. Application to the screening of maitotoxin-like compounds in Gambierdiscus spp. Toxicon 2010, 56, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J. Detection of Toxins Associated with Ciguatera Fish Poisoning; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; UNESCO Publishing: Paris, France, 2003; pp. 270–271. [Google Scholar]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
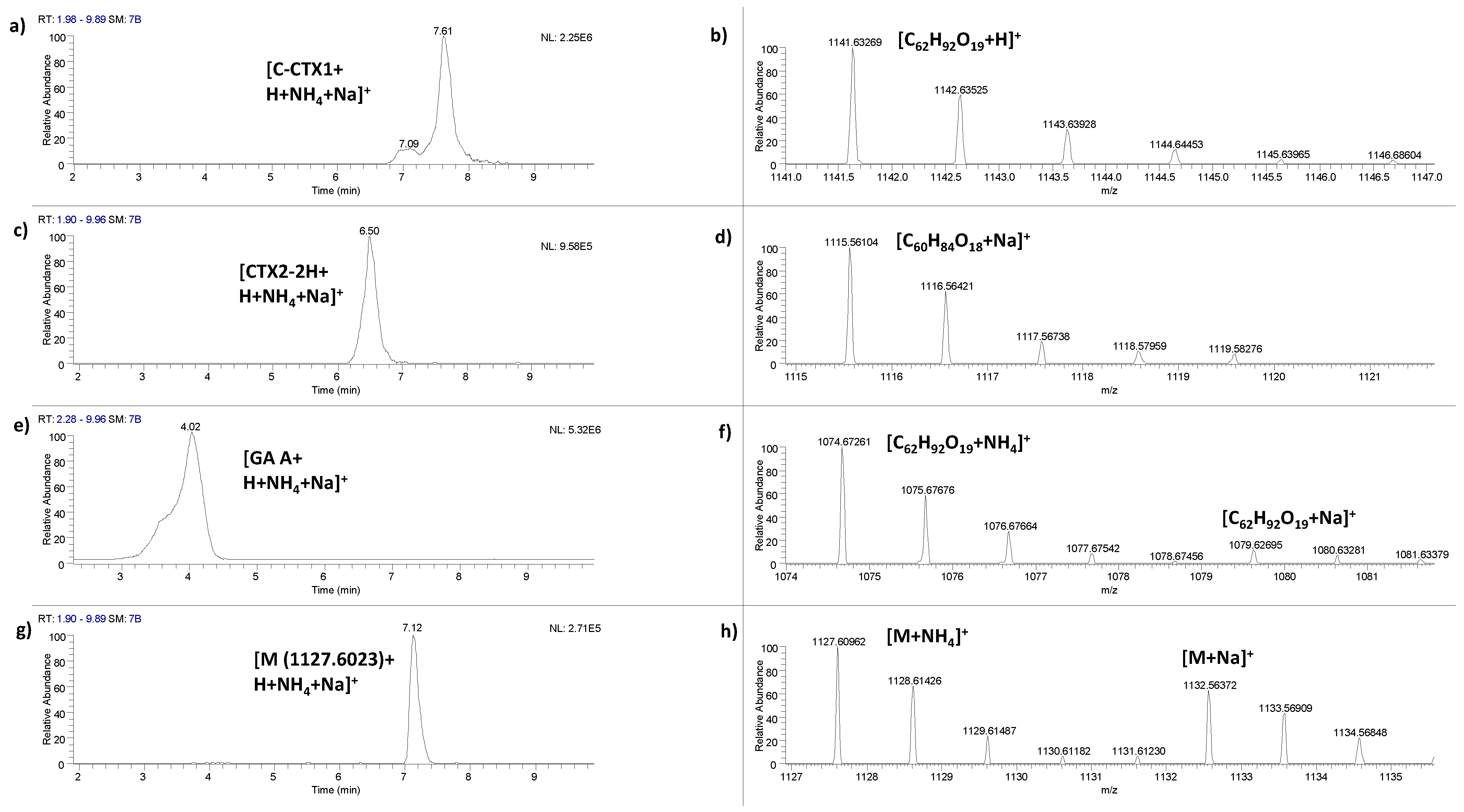{kind=link}
| MUSCLE | LIVER | |||||
|---|---|---|---|---|---|---|
| Code | Species | Location | CTX-like Toxicity (µg CTX1B equiv·kg−1) | LOQ (µg CTX1B equiv·kg−1) | CTX-like Toxicity (µg CTX1B equiv·kg−1) | LOQ (µg CTX1B equiv·kg−1) |
| 35 * | M. augusti | Desertas Islands | <LOQ | 0.013 | 0.270 ± 0.121 | 0.044 |
| 27 * | G. unicolor | Selvagens Islands | 0.034 ± 0.006 | 0.029 | 0.216 ± 0.051 | 0.011 |
| 25 * | G. unicolor | Selvagens Islands | <LOQ | 0.005 | 0.212 ± 0.125 | 0.041 |
| 26 * | G. unicolor | Selvagens Islands | 0.039 ± 0.001 | 0.002 | 0.187 ± 0.023 | 0.082 |
| 39 | M. helena | Desertas Islands | 0.083 ± 0.014 | 0.026 | 0.158 ± 0.022 | 0.044 |
| 34 | M. augusti | Desertas Islands | 0.065 ± 0.000 | 0.013 | 0.067 ± 0.022 | 0.044 |
| 14 | B. capriscus | Selvagens Islands | <LOQ | 0.010 | 0.060 ± 0.000 | 0.030 |
| 38 | M. helena | Desertas Islands | <LOQ | 0.013 | 0.050 ± 0.008 | 0.044 |
| 28 | G. unicolor | Selvagens Islands | <LOQ | 0.153 | 0.034 ± 0.000 | 0.004 |
| 6 | B. scrofa | Selvagens Islands | <LOQ | 0.016 | 0.015 ± 0.001 | 0.002 |
| 1 | M. fusca | Desertas Islands | <LOQ | 0.007 | 0.014 ± 0.000 | 0.008 |
| 13 | K. sectatrix | Selvagens Islands | <LOQ | 0.015 | 0.011 ± 0.008 | 0.003 |
| 37 | M. helena | Desertas Islands | <LOQ | 0.030 | 0.010 ± 0.002 | 0.004 |
| 2 | C. labrosus | Desertas Islands | <LOQ | 0.020 | <LOQ | 0.008 |
| 29 | C. labrosus | Desertas Islands | - | - | <LOQ | 0.047 |
| 31 | S. cretense | Desertas Islands | - | - | <LOQ | 0.178 |
| 36 | M. helena | Desertas Islands | - | - | <LOQ | 0.088 |
| Species | Weight of Specimen (kg) | Location (year) | Tissue | Toxicity According to CBA (µg equiv·kg−1) | Toxin Concentration by LC-MS Methods (µg equiv·kg−1) | Reference |
|---|---|---|---|---|---|---|
| Balistes capriscus (Grey triggerfish) | 2.0 | SI (2017) | Muscle | 0.03 (C-CTX1) | [45] | |
| 0.5–2.6 | SI (2018) | Muscle | 0.09 (C-CTX1) | [59] | ||
| 0.99 | SI (2014) | Liver | 0.060 (CTX1B) | This study | ||
| Bodianus scrofa (Barred hogfish) | 1.6 | SI (2017) | Muscle | 0.11 (C-CTX1) | [45] | |
| 0.8 | SI (2017) | Muscle | 0.06 (C-CTX1) | [45] | ||
| n/a | SI (2018) | Muscle | id: C-CTX1 | [49] | ||
| 1.1–3.0 | SI (2018) | Muscle | 0.04–0.75 (CTX1B) | 0.08–0.48 (C-CTX1) | [59] | |
| 2.4 | SI (2014) | Liver | 0.015 (CTX1B) | This study | ||
| Diplodus cervinus (Zebra seabream) | 2.8 | SI (2018) | Muscle | 0.37 (CTX1B) | id: C-CTX analogue 1157 m/z | [59] |
| Epinephelus marginatus (Dusky grouper) | 19.5 | SI (2016) | Muscle | 0.05 (C-CTX1) | [45] | |
| Gymnothorax unicolor (Brown moray) | 1.69 | SI (2013) | Liver | 0.212 (CTX1B) | id: CTX analogue 1127.6 m/z, C-CTX1 analogue 1141.6 m/z | This study |
| 1.21 | SI (2013) | Muscle | 0.039 (CTX1B) | id: CTX analogue 1127.6 m/z, dihydro-CTX2 analogue 1115.6 m/z | This study | |
| Liver | 0.187 (CTX1B) | id: CTX analogue 1127.6 m/z, C-CTX1 analogue 1141.6 m/z, dihydro-CTX2 analogue 1115.6 m/z | This study | |||
| 1.3 | SI (2013) | Muscle | 0.034 (CTX1B) | id: CTX analogue 1127.6 m/z, dihydro-CTX2 analogue 1115.6 m/z | This study | |
| Liver | 0.216 (CTX1B) | id: CTX analogue 1127.6 m/z, C-CTX1 analogue 1141.6 m/z | This study | |||
| 0.72 | SI (2013) | Liver | 0.034 (CTX1B) | This study | ||
| Kyphosus sectatrix (Bermuda sea chub) | 3.4 | SI (2014) | Liver | 0.011 (CTX1B) | This study | |
| Mycteroperca fusca (Island grouper) | 4.6 | SI (2016) | Muscle | 0.25 (C-CTX1) | [45] | |
| 2.1 | DI (2014) | Liver | 0.014 (CTX1B) | This study | ||
| Muraena augusti (Black moray) | 1.7 | DI (2013) | Muscle | 0.065 (CTX1B) | This study | |
| Liver | 0.067 (CTX1B) | This study | ||||
| 1.3 | DI (2013) | Liver | 0.270 (CTX1B) | id:C-CTX1, dihydro-CTX2 analogue 1115.6 m/z, CTX analogue 1127.6 m/z | This study | |
| Muraena helena (Moray eel) | 1.68 | DI (2013) | Liver | 0.010 (CTX1B) | This study | |
| 3.23 | DI (2013) | Liver | 0.050 (CTX1B) | This study | ||
| 1.19 | DI (2013) | Muscle | 0.083 (CTX1B) | This study | ||
| Liver | 0.158 (CTX1B) | This study | ||||
| Pagrus pagrus (Red porgy) | 4.0 | SI (2016) | Muscle | 0.76 (C-CTX1) | [46] | |
| Seriola dumerili (Greater amberjack) | 70 | SI (2009) | Muscle (tail) | 37.3 (CTX3C) | 53.76 (CTX3C) | [44] |
| id: CTX3C, CTX analogue 1040.6 m/z, CTX analogue 1141.6 m/z | ||||||
| Muscle (head) | 40.6 (CTX3C) | 54.35 (CTX3C) | [44] | |||
| id: CTX3C, CTX analogue 1040.6 m/z, CTX analogue 1141.6 m/z | ||||||
| Muscle (ventral) | 5.1 (CTX3C) | 33.29 (CTX3C) | [44] | |||
| id: CTX1B, CTX3C, CTX analogue 1040.6 m/z, CTX analogue 1141.6 m/z | ||||||
| Muscle (mid) | 41.7 (CTX3C) | 53.37 (CTX3C) | [44] | |||
| id: CTX1B, CTX3C, CTX analogue 1040.6 m/z, CTX analogue 1141.6 m/z | ||||||
| Liver | 37.7 (CTX3C) | 48.60 (CTX3C) | [44] | |||
| id: CTX1B, CTX3C, CTX analogue 1040.6 m/z, CTX analogue 1141.6 m/z | ||||||
| Seriola fasciata (Lesser amberjack) | 30 | SI (2009) | Muscle (tail) | 40.6 (CTX3C) | 35.2 (CTX3C)id: CTX1B, CTX3C, CTX analogue 1040.6 m/z, CTX analogue 1141.6 m/z | [44] |
| 63 | SI (2012) | Muscle | 6.23 (CTX1B) | [36] | ||
| 37 | SI (2012) | Muscle | 4.59 (CTX1B) | [36] | ||
| 37 | SI (2008) | Muscle | 1.4 (C-CTX1) | 0.84 (C-CTX1) | [47] | |
| Seriola rivoliana (Longfin yellowtail) | 2.5–12.3 | SI (2018) | Muscle | 0.10 (CTX1B) | [59] | |
| n/a | SI (2008) | Muscle | 0.17 (C-CTX1) | id: C-CTX1 | [73] | |
| Serrianus atriacuda (Blacktail comber) | 0.2–0.3 | SI (2018) | Muscle | 0.006–0.02 (CTX1B) | [59] | |
| Sparisoma cretense (Parrotfish) | 0.4–0.8 | SI (2018) | Muscle | 0.006–0.04 (CTX1B) | [59] | |
| Sphyraena viridensis (Yellowmouth barracuda) | 2.2–5.9 | SI (2018) | Muscle | 0.22 (CTX1B) | 0.14 (C-CTX1) | [59] |
| Code | Common Name | Species | Location | Date of Capture | Weight (kg) | Length (mm) | FB | G |
|---|---|---|---|---|---|---|---|---|
| 1 | Comb grouper | M. fusca | Desertas Islands | 25 September 2014 | 2.106 | 505 | C | F |
| 2 | Grey mullet | C. labrosus | Desertas Islands | 25 September 2014 | 1.972 | 572 | O | M |
| 6 | Barred hogfish | B. scrofa | Selvagens Grande | 24 May 2014 | 2.432 | 500 | C | M |
| 13 | Bermuda sea chub | K. sectatrix | Selvagens Grande | 25 May 2014 | 3.431 | 570 | O | F |
| 14 | Grey triggerfish | B.capriscus | Selvagens Grande | 25 May 2014 | 0.997 | 388 | C | F |
| 25 | Brown moray | G. unicolor | Selvagens Grande | 31 December 2013 | 1.688 | 894 | C | no det |
| 26 | Brown moray | G. unicolor | Selvagens Grande | 31 December 2013 | 1.210 | 838 | C | no det |
| 27 | Brown moray | G. unicolor | Selvagens Grande | 31 December 2013 | 1.305 | 812 | C | no det |
| 28 | Brown moray | G. unicolor | Selvagens Grande | 31 December 2013 | 0.721 | 717 | C | no det |
| 29 | Grey mullet | C. labrosus | Desertas Islands | 27 December 2013 | 1.896 | 545 | O | M |
| 31 | Parrotfish | S. cretense | Desertas Islands | 27 December 2013 | 0.412 | 307 | O | F |
| 34 | Black moray | M. augusti | Desertas Grande | 8 November 2013 | 1.669 | 917 | C | M |
| 35 | Black moray | M. augusti | Desertas Grande | 8 November 2013 | 1.397 | 838 | C | M |
| 36 | Moray eel | M. helena | Desertas Grande | 8 November 2013 | 1.339 | 893 | C | no det |
| 37 | Moray eel | M. helena | Desertas Grande | 8 November 2013 | 1.680 | 931 | C | F |
| 38 | Moray eel | M. helena | Desertas Grande | 8 November 2013 | 3.234 | 1070 | C | M |
| 39 | Moray eel | M. helena | Desertas Grande | 8 November 2013 | 1.193 | 856 | C | M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tudó, À.; Rambla-Alegre, M.; Flores, C.; Sagristà, N.; Aguayo, P.; Reverté, L.; Campàs, M.; Gouveia, N.; Santos, C.; Andree, K.B.; et al. Identification of New CTX Analogues in Fish from the Madeira and Selvagens Archipelagos by Neuro-2a CBA and LC-HRMS. Mar. Drugs 2022, 20, 236. https://0-doi-org.brum.beds.ac.uk/10.3390/md20040236
Tudó À, Rambla-Alegre M, Flores C, Sagristà N, Aguayo P, Reverté L, Campàs M, Gouveia N, Santos C, Andree KB, et al. Identification of New CTX Analogues in Fish from the Madeira and Selvagens Archipelagos by Neuro-2a CBA and LC-HRMS. Marine Drugs. 2022; 20(4):236. https://0-doi-org.brum.beds.ac.uk/10.3390/md20040236
Chicago/Turabian StyleTudó, Àngels, Maria Rambla-Alegre, Cintia Flores, Núria Sagristà, Paloma Aguayo, Laia Reverté, Mònica Campàs, Neide Gouveia, Carolina Santos, Karl B. Andree, and et al. 2022. "Identification of New CTX Analogues in Fish from the Madeira and Selvagens Archipelagos by Neuro-2a CBA and LC-HRMS" Marine Drugs 20, no. 4: 236. https://0-doi-org.brum.beds.ac.uk/10.3390/md20040236