Vasorelaxation, Induced by Dictyota pulchella (Dictyotaceae), a Brown Alga, Is Mediated via Inhibition of Calcium Influx in Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
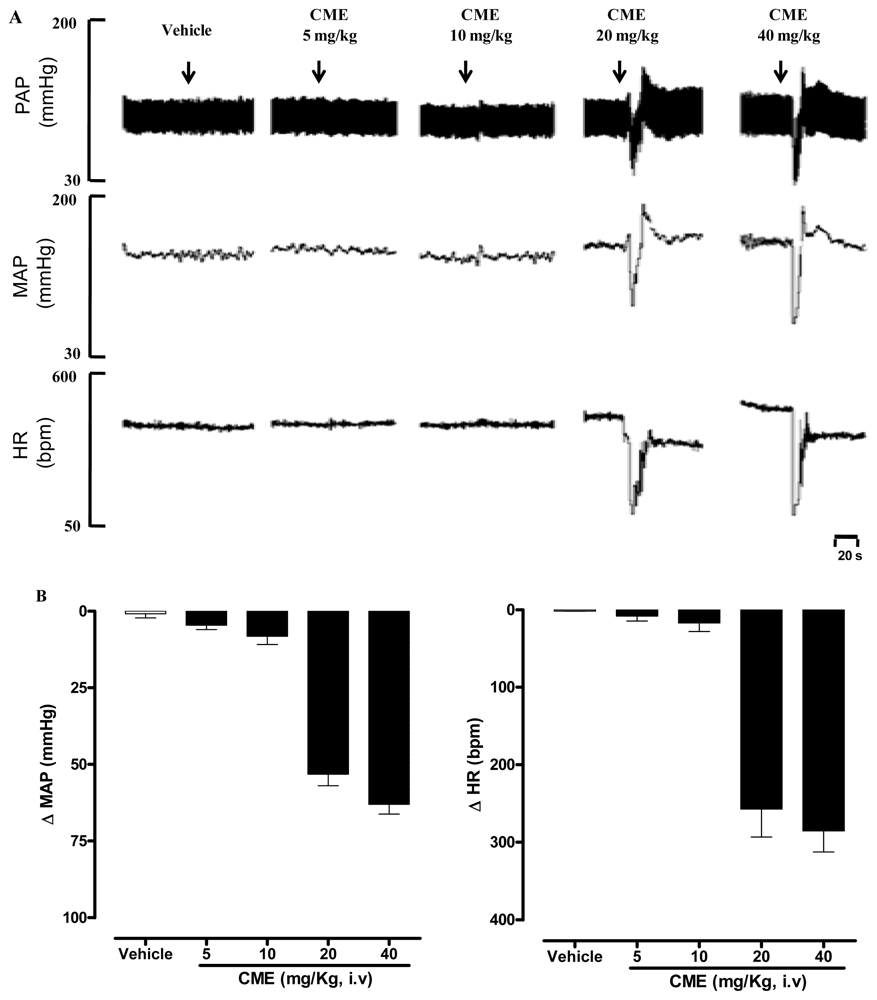
2.1. Dictyota pulchella Elicits Hypotension and Bradycardia in Rats
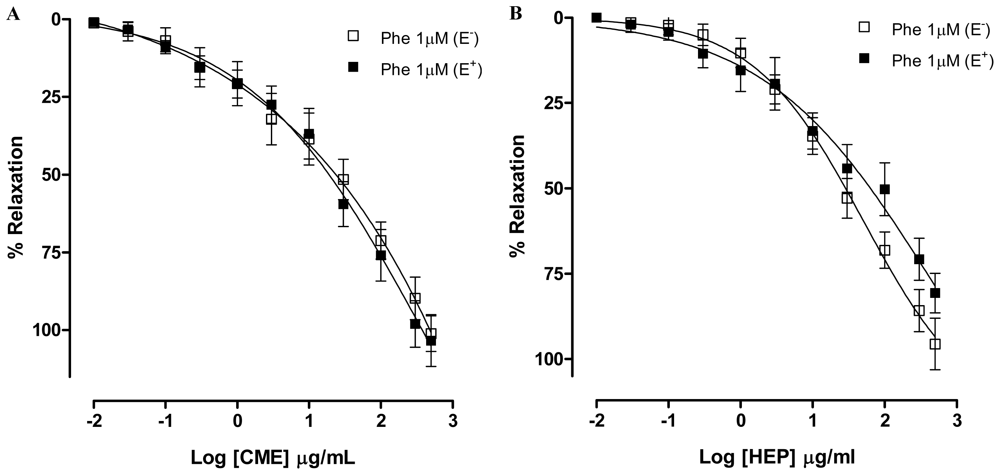
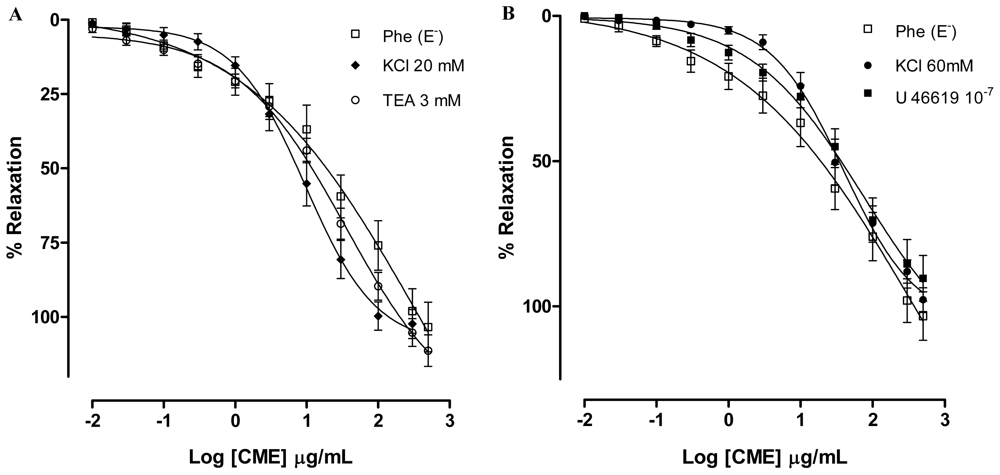
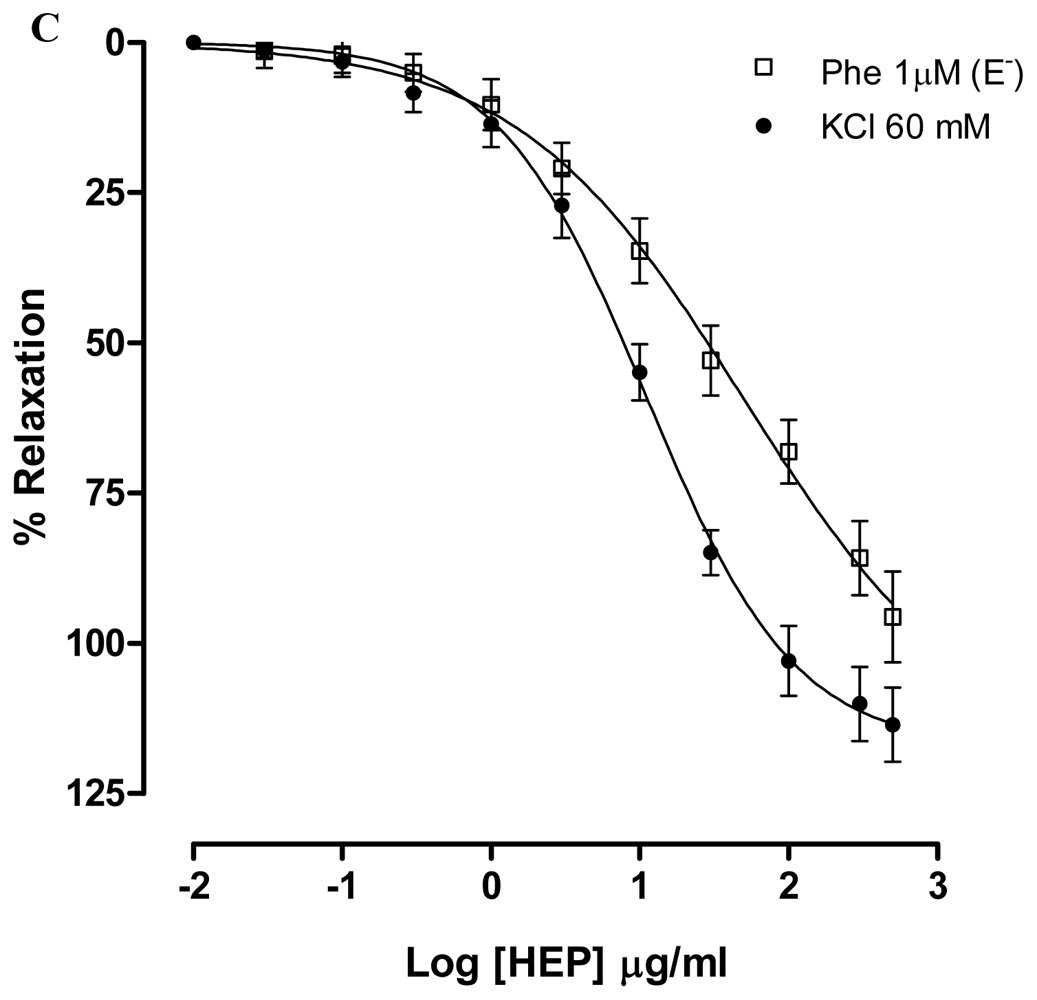
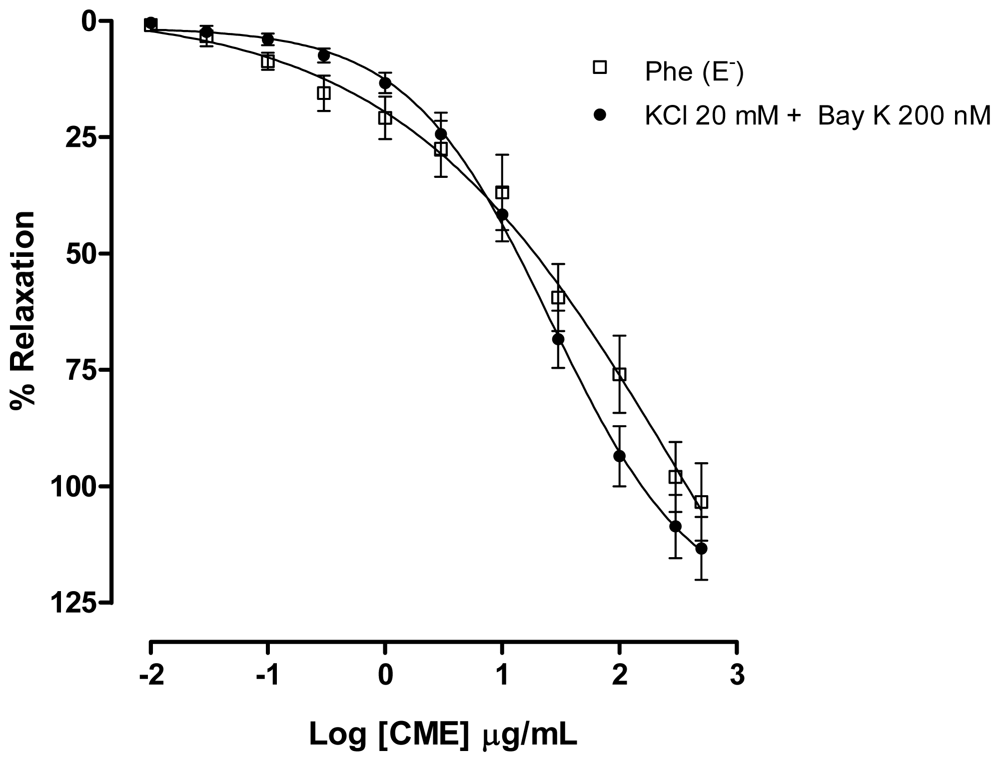
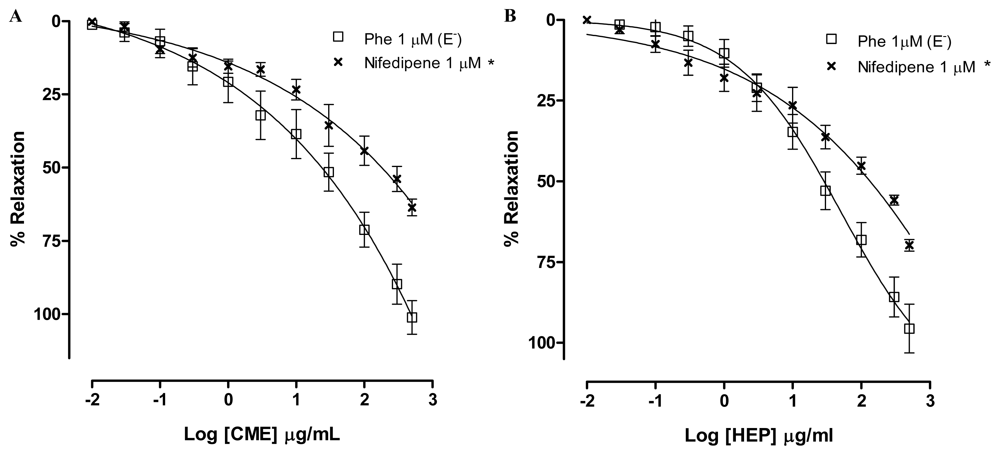
2.2. Dictyota pulchella Elicits Vasorelaxation Mediated by the Blockade of Calcium Influx in Isolated Mesenteric Artery Rings
3. Experimental Section
3.1. Algal Material and Preparation of CH2Cl2/MeOH Extract (CME) and Hexane/EtOAc Phase (HEP) from Dictyota pulchella
3.2. Drugs and Solutions
3.3. Animals
3.4. In Vivo Experiments
3.5. In Vitro Experiments
3.5.1. Tissue Preparation
3.5.2. Measurement of Vascular Relaxation Elicited by CME or HEP
3.5.3. Effect of CME or HEP on KCl (60 mM) or U-46619 (100 nM)-Induced Contractions in Rings without Endothelium or after KCl (20 mM) and Tetraethylamonium (3 mM) Incubation
3.5.4. Depolarization Induced by High Extracellular K+ Concentration
3.5.5. Effect of CME on the Contraction Elicited by S(−)-Bay K 8644 (200 nM) or Nifedipine (1 μM) in Mesenteric Artery Rings without Endothelium
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Matsubara, K; Matsuura, Y; Hori, K; Miyazawa, K. An anticoagulant proteoglycan from the marine green alga, Codium pugniformis. J. Appl. Phycol 2000, 12, 9–14. [Google Scholar]
- Athukorala, Y; Lee, K; Kim, S; Jeon, Y. Anticoagulant activity of marine green and brown algae collected from Jeju Island in Korea. Bioresour. Technol 2007, 98, 1711–1716. [Google Scholar]
- Artan, M; Li, Y; Karadeniz, F; Lee, S; Kim, M; Kim, S. Anti-HIV-1 activity of phloroglucinol derivative, 6,6′-bieckol, from Ecklonia cava. Bioorgan. Med. Chem 2008, 16, 7921–7926. [Google Scholar]
- Huheihel, M; Ishanu, V; Tal, J; Arad, S. Activity of Porphyridium sp. polysaccharide against herpes simplex viruses in vitro and in vivo. J. Biochem. Biophys. Methods 2002, 50, 189–200. [Google Scholar]
- Heo, SJ; Park, EJ; Lee, KW; Jeon, YJ. Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour. Technol 2005, 96, 1613–1623. [Google Scholar]
- Zou, Y; Qian, Z; Li, Y; Kim, M; Lee, S; Kim, S. Antioxidant effects of phlorotannins isolated from Ishige okamurae in free radical mediated oxidative systems. J. Agric. Food Chem 2008, 56, 7001–7009. [Google Scholar]
- Li, Y; Lee, S; Le, Q; Kim, M; Kim, S. Anti-allergic effects of phlorotannins on histamine release via binding inhibition between IgE and Fc RI. J. Agric. Food Chem 2008, 56, 12073–12080. [Google Scholar]
- Kong, CS; Kim, JA; Yoon, NY; Kim, SK. Induction of apoptosis by phloroglucinol derivative from Ecklonia cava in MCF-7 human breast cancer cells. Food Chem. Toxicol 2009, 47, 1653–1658. [Google Scholar]
- Pangestuti, R; Kim, S-K. Neuroprotective effects of marine algae. Mar. Drugs 2011, 9, 803–818. [Google Scholar]
- Kelecom, A. Secondary metabolites from marine microorganisms. An. Acad. Bras. Cienc 2002, 74, 151–170. [Google Scholar]
- Smit, AJ. Medicinal and pharmaceutical uses of seaweeds natural products: a review. J. Appl. Physcol 2004, 16, 245–262. [Google Scholar]
- Sousa, APA; Torres, MR; Pessoa, C; Moraes, MO; Rocha-Filho, FD; Alves, APNN; Costa-Lotufo, LV. In vivo growth-inhibition of Sarcoma 180 tumor by alginates from brown seaweed Sargassum vulgare. Carbohydr. Polym 2007, 69, 7–13. [Google Scholar]
- Siamopoulou, P; Bimplakis, A; Iliopoulou, D; Vagias, C; Cos, P; Vanden Berghe, D; Roussis, V. Diterpenes from the brown algae Dictyota dichotoma and Dictyota linearis. Phytochemistry 2004, 65, 2025–2030. [Google Scholar]
- Sacchettini, JC; Poulter, CD. Creating isoprenoid diversity. Science 1997, 277, 1788–1789. [Google Scholar]
- Dewick, PM. The biosynthesis of C5–C25 terpenoid compounds. Nat. Prod. Rep 2002, 19, 181–222. [Google Scholar]
- Dubey, VS; Bhalla, R; Luthra, R. An overview of the non-mevalonate pathway for terpenoid biosynthesis in plants. J. Biosci 2003, 28, 637–646. [Google Scholar]
- Tirapelli, CR; Ambrosio, SR; da Costa, FB; de Oliveira, AM. Evidence for the mechanisms underlying the effects of pimaradienoic acid isolated from the roots of Viguiera arenaria on rat aorta. Pharmacology 2004, 70, 31–38. [Google Scholar]
- Tirapelli, CR; dos Anjos, MNF; Bonaventura, D; Melo, MC; Ambrosio, SR; de Oliveira, AM; Bendhack, LM; da Costa, FB. Pimaradienoic acid inhibits vascular contraction and induces hypotension in normotensive rats. J. Pharm. Pharmacol 2008, 60, 453–459. [Google Scholar]
- Tirapelli, CR; Ambrosio, SR; de Oliveira, AM; Tostes, RC. Hypotensive action of naturally occurring diterpenes: A therapeutic promise for the treatment of hypertension. Fitoterapia 2010, 81, 690–702. [Google Scholar]
- Somova, LI; Shode, FO; Moodley, K; Govender, Y. Cardiovascular and diuretic activity of kaurene derivates of Xylopia aethiopica and Alepidea amatymbica. J. Ethnopharmacol 2001, 77, 165–174. [Google Scholar]
- Oliveira, AP; Furtado, FF; Silva, MS; Tavares, JF; Mafra, RA; Araújo, DA; Cruz, JS; Medeiros, IA. Calcium channel blockade as a target for the cardiovascular effects induced by the 8 (17), 12E, 14 labdatrien-18-oic acid (labdane-302). Vasc. Pharmacol 2006, 44, 338–344. [Google Scholar]
- American Heart Association. Heart disease and stroke statistics—2008 update. Circulation 2008, 117, 125–146.
- Lefkowits, RJ; Willerson, J. Prospects for cardiovascular research. JAMA 2001, 285, 581–587. [Google Scholar]
- Santos, MRV; Moreira, FV; Fraga, BP; de Souza, DP; Bonjardim, LR; Quintans, LJ, Jr. Cardiovascular effects of monoterpenes: a review. Rev. Bras. Farmacogn 2011, 21, 764–771. [Google Scholar]
- Fluckiger, JP; Sonnay, M; Boillat, N; Atkinson, J. Attenuation of baroreceptor reflex by general anesthetic agent in the normotensive rat. Eur. J. Pharmacol 1985, 109, 105–109. [Google Scholar]
- Smith, TL; Hutchins, PM. Anesthetic effects on hemodynamics of spontaneously hypertensive on Wistar-Kyoto rats. Am. J. Physiol 1980, 238, 539–544. [Google Scholar]
- Mulvany, MJ; Aalkjaer, C. Structure and function of small arteries. Physiol. Res 1990, 70, 921–951. [Google Scholar]
- Grover-Páez, F; Zavalza-Gómez, AB. Endothelial dysfunction and cardiovascular risk factors. Diabetes Res. Clin. Pract 2009, 84, 1–10. [Google Scholar]
- Furchgott, FF; Zawadzki, JV. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980, 288, 373–376. [Google Scholar]
- Côrtes, SF; Rezende, BA; Corriu, C; Medeiros, IA; Teixeira, MM; Lopes, MJ; Lemos, VS. Pharmacological evidence for the activation of potassium channels as the mechanism involved in the hypotensive and vasorelaxant effect of dioclein in rat small resistance arteries. Br. J. Pharmacol 2001, 133, 849–858. [Google Scholar]
- McNeill, JR; Jurgens, TM. A systematic review of mechanisms by which natural products of plant origin evoke vasodilation. Can. J. Phys. Pharmacol 2006, 84, 803–821. [Google Scholar]
- Kim, HY; Oh, H; Li, X; Cho, KW; Kanga, DG; Lee, HS. Ethanol extract of seeds of Oenothera odorata induces vasorelaxation via endothelium dependent NO-cGMP signaling through activation of Akt-eNOS-sGC pathway. J. Ethnopharmacol 2011, 133, 315–323. [Google Scholar]
- Ribeiro, TP; Porto, DL; Menezes, CP; Antunes, AA; Silva, DF; Sousa, DP; Nakao, LS; Braga, VA; Medeiros, IA. Unravelling the cardiovascular effects induced by α-terpineol: A role for the nitric oxide–cGMP pathway. Clin. Exp. Pharmacol. Phys 2010, 37, 811–816. [Google Scholar]
- Somlyo, AP; Somlyo, AV. Signal transduction and regulation in smooth muscle. Nature 1994, 372, 231–236. [Google Scholar]
- Ratz, PH; Berg, KM. 2-Aminoethoxydiphenyl borate inhibits KCl-induced vascular smooth muscle contraction. Eur. J. Pharmacol 2006, 541, 177–183. [Google Scholar]
- Navarro-Gonzalez, MF; Grayson, TH; Meaney, KR; Cribbs, LL; Hill, CE. Non-L-type voltage-dependent calcium channels control vascular tone of the rat basilar artery. Clin. Exp. Pharmacol. Phys 2009, 36, 55–66. [Google Scholar]
- Braga, VA. Dietary salt enhances angiotensin-II-induced superoxide formation in the rostral ventrolateral medulla. Auton Neurosci Basic Clin 2010, 155, 14–18, Samples Availability: Available from the authors.. [Google Scholar]







© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Queiroz, T.M.; Machado, N.T.; Furtado, F.F.; Oliveira-Filho, A.A.; Alustau, M.C.; Figueiredo, C.S.; Miranda, G.E.C.; Barbosa-Filho, J.M.; Braga, V.A.; Medeiros, I.A. Vasorelaxation, Induced by Dictyota pulchella (Dictyotaceae), a Brown Alga, Is Mediated via Inhibition of Calcium Influx in Rats. Mar. Drugs 2011, 9, 2075-2088. https://0-doi-org.brum.beds.ac.uk/10.3390/md9102075
Queiroz TM, Machado NT, Furtado FF, Oliveira-Filho AA, Alustau MC, Figueiredo CS, Miranda GEC, Barbosa-Filho JM, Braga VA, Medeiros IA. Vasorelaxation, Induced by Dictyota pulchella (Dictyotaceae), a Brown Alga, Is Mediated via Inhibition of Calcium Influx in Rats. Marine Drugs. 2011; 9(10):2075-2088. https://0-doi-org.brum.beds.ac.uk/10.3390/md9102075
Chicago/Turabian StyleQueiroz, Thyago M., Natália T. Machado, Fabíola F. Furtado, Abrahão A. Oliveira-Filho, Maria C. Alustau, Camila S. Figueiredo, George E. C. Miranda, José M. Barbosa-Filho, Valdir A. Braga, and Isac A. Medeiros. 2011. "Vasorelaxation, Induced by Dictyota pulchella (Dictyotaceae), a Brown Alga, Is Mediated via Inhibition of Calcium Influx in Rats" Marine Drugs 9, no. 10: 2075-2088. https://0-doi-org.brum.beds.ac.uk/10.3390/md9102075